不同生境对赤皮青冈光合特性的影响
2017-05-11朱宁华曹基武
张 斌, 朱宁华, 曹基武
(1中南林业科技大学 林学院,湖南 长沙 410004;2中南林业科技大学 植物园,湖南 长沙 410004)
不同生境对赤皮青冈光合特性的影响
张 斌1, 朱宁华1, 曹基武2
(1中南林业科技大学 林学院,湖南 长沙 410004;2中南林业科技大学 植物园,湖南 长沙 410004)
【目的】分析赤皮青冈Cyclobalanopsisgilva的光合作用对不同生境条件的适应特征,为赤皮青冈的良种选育和科学栽培提供理论和数据支撑。【方法】以3种不同生境条件下的赤皮青冈天然居群为研究对象,用LI-6400便携式光合仪测定天然状态下赤皮青冈的光合生理指标日变化,并得出其光响应曲线的变化规律。【结果】不同生境条件下,赤皮青冈的净光合速率与蒸腾速率变化趋势基本一致,都在10:00和14:00出现峰值,13:00出现谷值;叶片温度、光合有效辐射、气孔导度与光合速率日变化均有很强的正相关关系。光响应曲线表明:赤皮青冈利用光能的能力受不同生境条件的影响(山间平地>山地阳坡>山地阴坡),3种生境条件下的光补偿点(Lcp)都比较低,山地阴坡条件下Lcp最低(2.257 mol·m-2·s-1),山间平地和山地阳坡条件下Lcp较接近(51 mol·m-2·s-1左右);光饱和点(Lsp)表现为山间平地>山地阳坡>山地阴坡。【结论】3种生境下的赤皮青冈均具有较强的环境适应能力, 赤皮青冈适应了山间平地和山地阳坡生境下光照时间长、强度大的特点,具有较强的光合能力和强光利用能力;而山地阴坡生境下的赤皮青冈提高了弱光利用能力。对不同生境条件的长期适应是赤皮青冈的光合能力产生差异的主要原因。
珍稀树种; 光合特性; 生境; 赤皮青冈
森林的光环境复杂多变,在空间上从林冠到林下、林窗到林缘,在时间上从森林破坏到恢复发展,植物的光环境无不在变化当中,而长期生活在不同生境条件下的同种植物,经过漫长的自然选择,产生了形态和生理特征上的差异,这反映了该植物对生境条件的适应能力[1]。因此,光合作用适应性及其与植物的生态分布和演替状态之间的联系多年来一直倍受关注[2-4]。光合作用是绝大多数植物生存的物质和能量基础,也是植物干物质积累和能量合成的重要途径,光合能力的大小受植物的遗传性和环境条件的共同影响,光合进程的启动和停止受光照强度的影响,因此植物对不同强度光照辐射的利用能力成为衡量植物对外界环境适应能力的重要指标,因此研究树木在特定生境条件下形成的需光特性和对光照辐射的生理响应,可以对树木引种驯化策略做出科学的估计[5-7]。此外,光补偿点、光饱和点等是反映植物需光特性的重要生理参数,已成为树木速生丰产早期鉴定及栽培措施制定的科学依据[8-10]。
赤皮青冈Cyclobalanopsisgilva是壳斗科Fagaceae青冈属Cyclobalanopsis常绿高大乔木,木材红褐色,是极具开发价值的优良用材树种[11]。目前赤皮青冈的研究多集中在幼苗繁育、需肥规律、抗性生理、光合规律等方面[12-15],不同学者研究发现,赤皮青冈10年生幼树光合速率日变化呈双峰型,蒸腾速率和气孔导度为单峰型[16],施肥对2年生赤皮青冈幼苗的光合促进作用明显[13],轻度水分胁迫能促使1年生赤皮青冈幼苗叶绿素含量显著增加[17],而不同种源间幼苗光合特性的差异主要来自气孔导度的影响[18],但少见对赤皮青冈在天然生境下的光合生理差异的研究。因此开展野生赤皮青冈大树的光合生理研究对于指导赤皮青冈优良品种选育和制定造林技术措施具有重要的意义。
1 材料与方法
1.1 试验区概况
试验在湖南八大公山保护区内进行,选择处于3种不同生境的赤皮青冈天然居群(树龄42~50年),对胸径30~40 cm、树高15 m左右的赤皮青冈进行光合生理指标的测定,3种不同的生境条件见表1。保护区属北亚热带山地湿润季风气候区,最低平均气温-2 ℃,最高平均气温22.8 ℃,常年相对湿度90%以上,年均降雨量2 000 mm以上,全年无霜期为258 d。

表1 试验地生境特征
1) 檵木Loropetalumchinense, 冬青Ilexchinensis, 构树Broussonetiapapyrifera, 杜鹃Rhododendronsimsii, 山茶Camelliajaponica, 三尖杉Cephalotaxusfortunei, 山矾Symplocossumuntia, 木姜子Litseapungens。
1.2 试验方法
1.2.1 光合日变化测定 采用LI-6400便携式光合仪测定光合生理指标,试验在5月下旬进行,选择天气晴朗少云的连续3 d,每天测量一个生境条件,从08:00—17:00每小时测量1次,每个生境条件下选1~2株生长良好的成年大树,从树冠4个方向上选取4片无机械损伤、无虫眼、无病虫害的成熟叶标记并测定,测试结果取平均值。主要测定指标有净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、叶片温度(θL)、光合有效辐射(PAR),并计算:光能利用效率(SUE), SUE =Pn/PAR; 水分利用效率(WUE),WUE =Pn/Tr; 瞬时羧化效率(Ce),Ce=Pn/Ci; 气孔限制值(Ls),Ls=1-Ci/Ca, Ca为大气CO2浓度。
1.2.2 光响应曲线的测定 由红蓝光源控制光照辐射范围为0~2 000 μmol·m-2·s-1,梯度设置为0、20、50、100、150、200、400、600、800、1 000、1 200、1 500、1 800、2 000 μmol·m-2·s-1,分别测量相应光合生理生态因子的变化,间隔2 min。测定时叶片温度为(25±1) ℃,光合有效辐射为(500±5) μmol·m-2·s-1,空气相对湿度为(75±1)%。根据光响应曲线模型计算赤皮青冈在不同生境下的光补偿点(Lcp)、光饱和点(Lsp)及表观量子效率(AQY)等光合生理参数,光响应曲线拟合采用经典的Farquhar模型[19-20],模型公式如下:
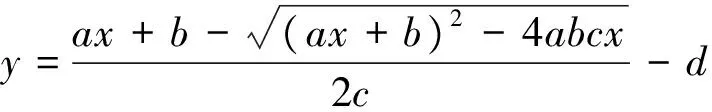
式中,y为净光合速率(Pn),x为光合有效辐射(PAR),a、b、c、d分别为表观量子效率(AQY)、最大净光合速率(Pnmax)、方程系数、呼吸速率(RD)。
1.3 数据处理
采用SPSS 17.0软件进行均值和标准差统计、单因素方差分析、多重比较分析(LSD法)、数据的相关性分析及相关系数统计(Pearson法);采用Excel 2013软件制作图表。
2 结果与分析
2.1 光合特性日变化
2.1.1 净光合速率日变化 由图1A可知,3种生境条件下赤皮青冈的净光合速率日变化趋势基本一致,呈双峰型变化,在10:00之前,随着光照辐射的增加,3种生境条件下赤皮青冈的净光合速率随之上升,10:00时3种生境条件下的净光合速率出现第1个峰值,且为山间平地(8.248 μmol·m-2·s-1)>山地阳坡(7.866 μmol·m-2·s-1)>山地阴坡(6.806 μmol·m-2·s-1),15:00又达到另一个峰值,山间平地(7.738 μmol·m-2·s-1)>山地阳坡(7.470 μmol·m-2·s-1)>山地阴坡(6.818 μmol·m-2·s-1)。10:00—15:00光照辐射略有增加,净光合速率下降不明显,但不同生境条件间差异不显著(显著性水平0.604~0.880),此时可能是赤皮青冈逐渐进入光合午休状态的缘故。净光合速率日均值之间的差异不明显,但表现出山间平地>山地阳坡>山地阴坡的趋势(图1B)。
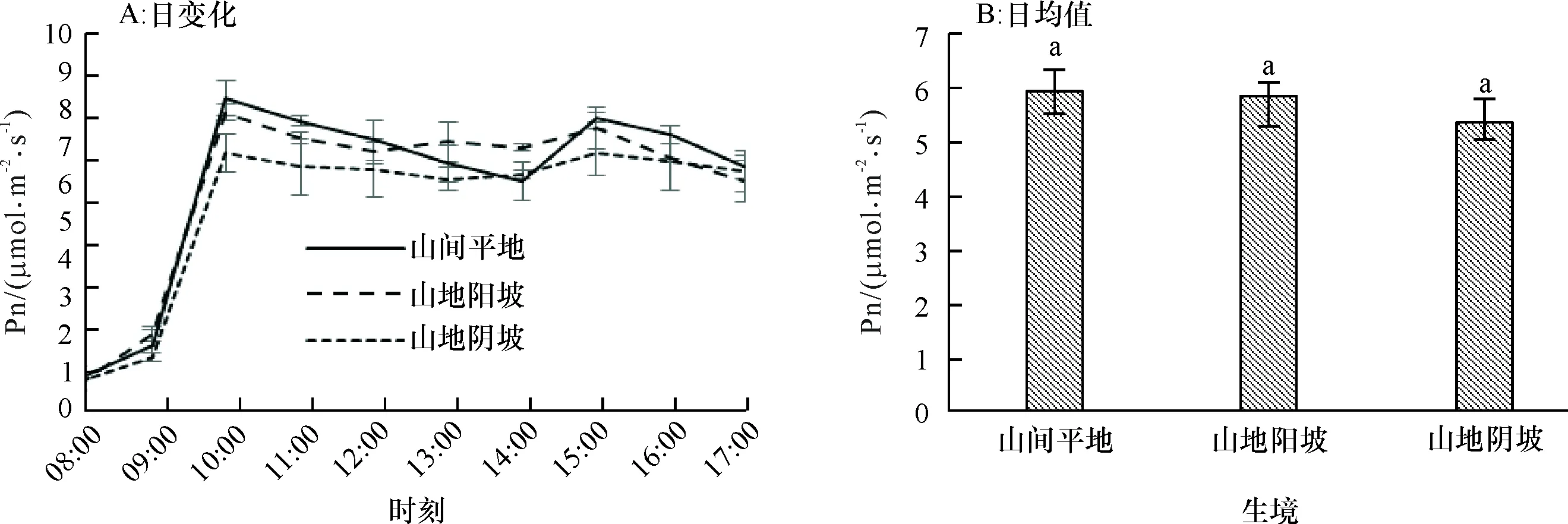
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.2 蒸腾速率日变化 山间平地生境下赤皮青冈蒸腾速率日变化呈双峰曲线变化趋势,叶片蒸腾速率在10:00出现第1个峰值(0.760 mmol·m-2·s-1),13:00出现谷值(0.206 mmol·m-2·s-1),14:00出现第2个峰值(1.926 mmol·m-2·s-1),之后蒸腾速率下降。山地阴坡生境下赤皮青冈蒸腾速率日变化呈单峰曲线,08:00—14:00呈逐渐增加趋势,并在14:00出现峰值(1.083 mmol·m-2·s-1),之后呈下降趋势。山地阳坡生境下赤皮青冈蒸腾速率日变化呈不太典型的双峰曲线,10:00出现第1个峰值(0.756 mmol·m-2·s-1),13:00出现谷值(0.161 mmol·m-2·s-1),14:00出现第2个峰值(0.355 mmol·m-2·s-1)(图2A)。可见,山地阳坡生境下赤皮青冈蒸腾速率变化大致趋势与山间平地一致,而山地阴坡生境下赤皮青冈则呈现单峰现象,利用全天不同时刻的蒸腾速率进行比较发现,3种生境条件下赤皮青冈蒸腾速率日变化无显著性差异(P为0.168~0.488),但在14:00外界光照辐射达到最大值,不同生境条件下的赤皮青冈出现的第2个峰值存在显著性差异(P为0.042)。
从蒸腾速率日均值变化规律来看,不同生境条件对赤皮青冈的蒸腾速率产生了显著的影响(图2B),山间平地的日均蒸腾速率最大,山地阳坡的蒸腾速率最小,比山间平地生境下的赤皮青冈日均蒸腾速率下降了35.13%,其原因可能是在阳坡生境下接受的光照辐射强、升温快、且土壤含水量较低,导致长期在此生境下的赤皮青冈对高温蒸腾和土壤相对较低含水量形成了这样的适应机制。
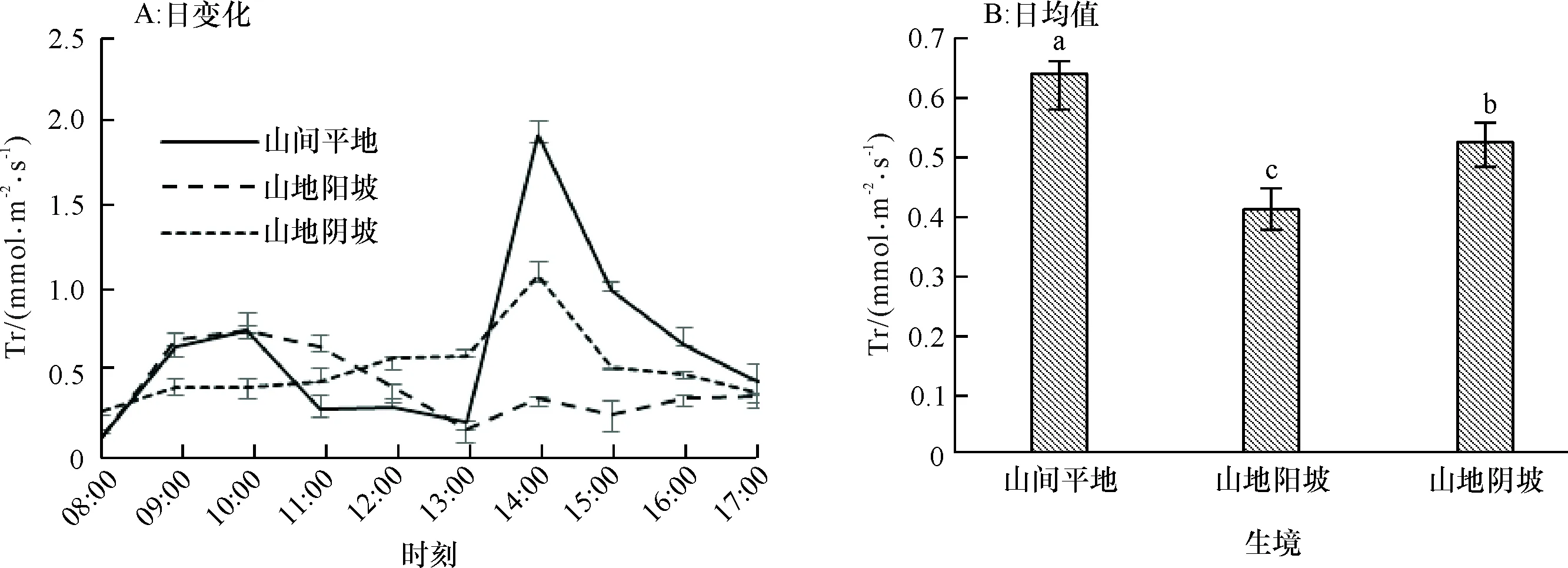
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.3 气孔导度日变化 从08:00—10:00,由于林内光照强度增加,山间平地、山地阳坡和山地阴坡生境下赤皮青冈的气孔导度均明显增加,之后增幅略有下降(图3A),在光照强度较高的14:00,气孔的开放程度达到最大值,其中山间平地下赤皮青冈的气孔导度达到最大值(4.703 mmol·m-2·s-1),山地阳坡和山地阴坡分别比山间平地生境低了12.31%和22.32%,但山地阳坡生境的气孔导度最大值较其他生境晚1 h出现,且最大值达到了5.096 mmol·m-2·s-1,说明阳坡生境下光照辐射在15:00达到最大,赤皮青冈通过调整气孔导度来达到降低温度、减少高温伤害的效果;3种生境条件下气孔导度日变化均值差别并不显著(P为0.107~0.593)(图3B),说明生境对赤皮青冈的气孔导度存在影响,但影响程度不大,特别是阳坡生境下接受的光照辐射相对较多,赤皮青冈为适应强光照和高温的影响,通过降低气孔导度来减少蒸腾,而阴坡生境下,赤皮青冈通过增加气孔导度来增加体内CO2浓度,从而达到提高光合速率的能力。
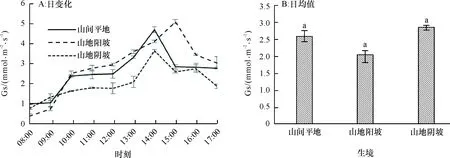
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.4 叶片温度日变化 一天中随着大气温度和光照强度的增加,不同生境条件下赤皮青冈的叶片温度(θL)均呈逐渐上升的趋势(图4A),上升趋势与大气温度同步,气温的上升导致空气相对湿度降低,叶片水压亏缺也呈缓慢加大趋势,并与光照强度和外界温度变化规律一致,高温低湿的外界环境可引起植物蒸腾失水而产生生理干旱。10:00之后,随着环境温度的上升,气孔导度增加、蒸腾速率增大,叶片温度降低,13:00之后,随着太阳辐射的减弱,大气温度降低,但由于植物温度相对空气温度的滞后现象,叶片温度出现短暂的轻微回升(P为0.713~0.921),不同生境之间的差异也不显著(P为0.464~0.804)(图4B)。
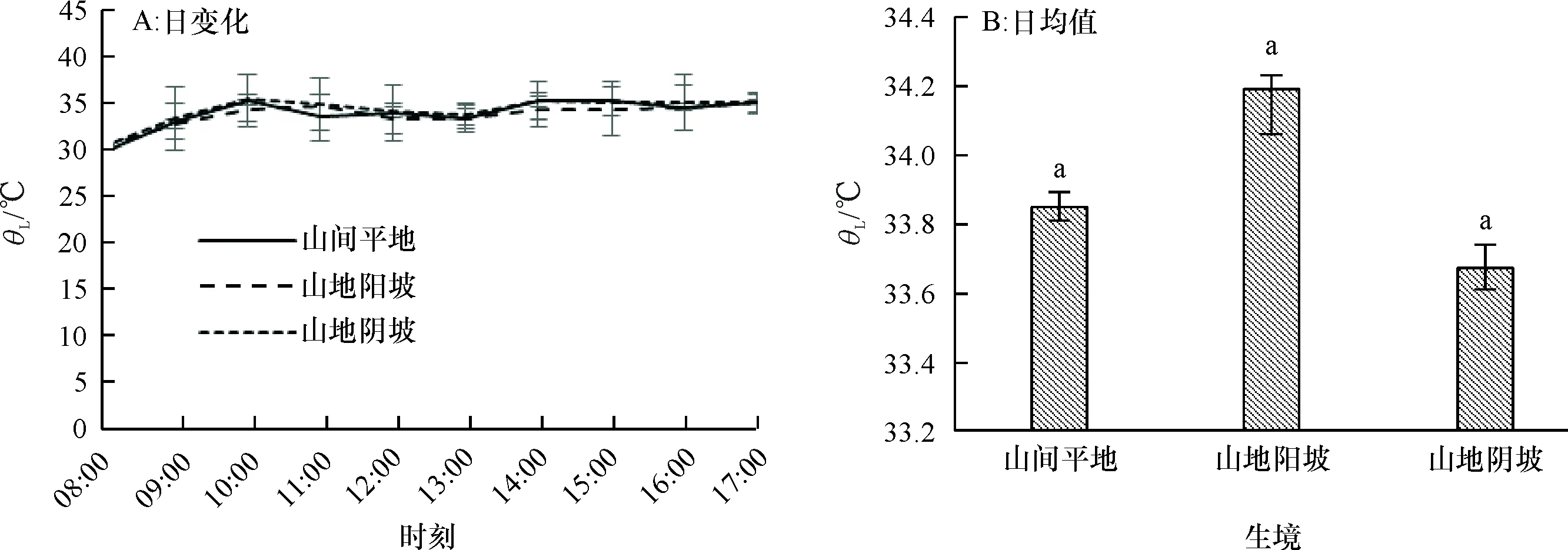
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.5 胞间二氧化碳浓度日变化 从图5A可以看出,不同生境条件下,赤皮青冈的胞间二氧化碳浓度(Ci)的日变化呈M型,即在光照强度最大的中午和傍晚Ci值下降,中午通过光合午休现象调节气孔的关闭来降低细胞内的CO2浓度,从而保护赤皮青冈免受高温的不良影响,从Ci的日均值(图5B)来看,山地阴坡>山地阳坡>山间平地,平地比山地环境的Ci值低13.27%,但差异不显著(P为0.052),Ci越小,说明该生境条件下赤皮青冈对于高温和强光照辐射环境越敏感。
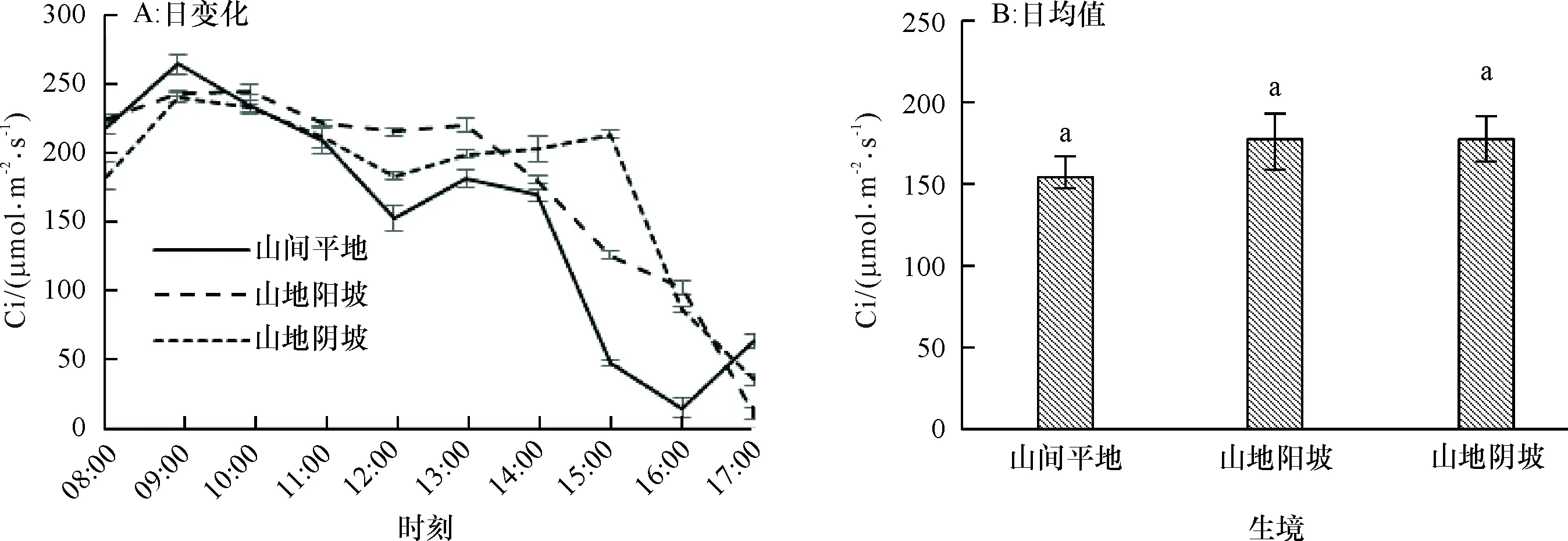
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
Fig.5 Daily changes and average values of intercellular CO2concentrations (Ci) ofCyclobalanopsisgilvain different habitats
2.1.6 气孔限制值日变化 气孔限制值(Ls)反映了植物叶片气孔导度的强弱以及由此带来的光合速率的变化[21],从图6A可以看出,不同生境条件下赤皮青冈的Ls表现出下午时段(14:00—17:00)高于其他时段,其中山间平地生境下的Ls值在16:00达到最大,而后略有下降;山地阳坡生境的Ls值呈缓慢增加的趋势,在14:00后迅速增加;山地阴坡生境下的Ls在15:00之前变化较小,之后开始迅速增加。Ls在不同生境下的变化趋势与胞间CO2浓度趋势相反,随着Ls的增加,Pn下降不明显,说明Pn的变化与Ls和Ci的关系不大。从Ls的日均值(图6B)来看,山地阳坡和山地阴坡环境下赤皮青冈的Ls比山间平地环境低了9.49%和9.63%,达到了显著性的差异(P为0.048)。
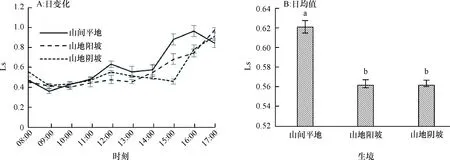
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.7 瞬时羧化效率日变化 瞬时羧化效率(Ce)反映了植物叶片利用CO2能力的高低。山间平地生境条件下的赤皮青冈在16:00时表现出最大的羧化效率,而山地阳坡和阴坡生境下最大羧化效率均出现在17:00时(图7A),特别是山间平地和山地阳坡生境条件,比山地阴坡生境下的赤皮青冈日均瞬时羧化效率高出120.31%和96.64%,均达到了显著水平(P为0.000),山地阴坡生境降低了赤皮青冈对CO2的利用效率。从瞬时羧化效率日均值(图7B)来看,山间平地生境下的Ce为0.101,山地阳坡的Ce比山间平地下降了10.74%,但差异不显著(P为0.062),山地阴坡的Ce比山间平地下降了54.61%,且差异显著(P为0.000)。
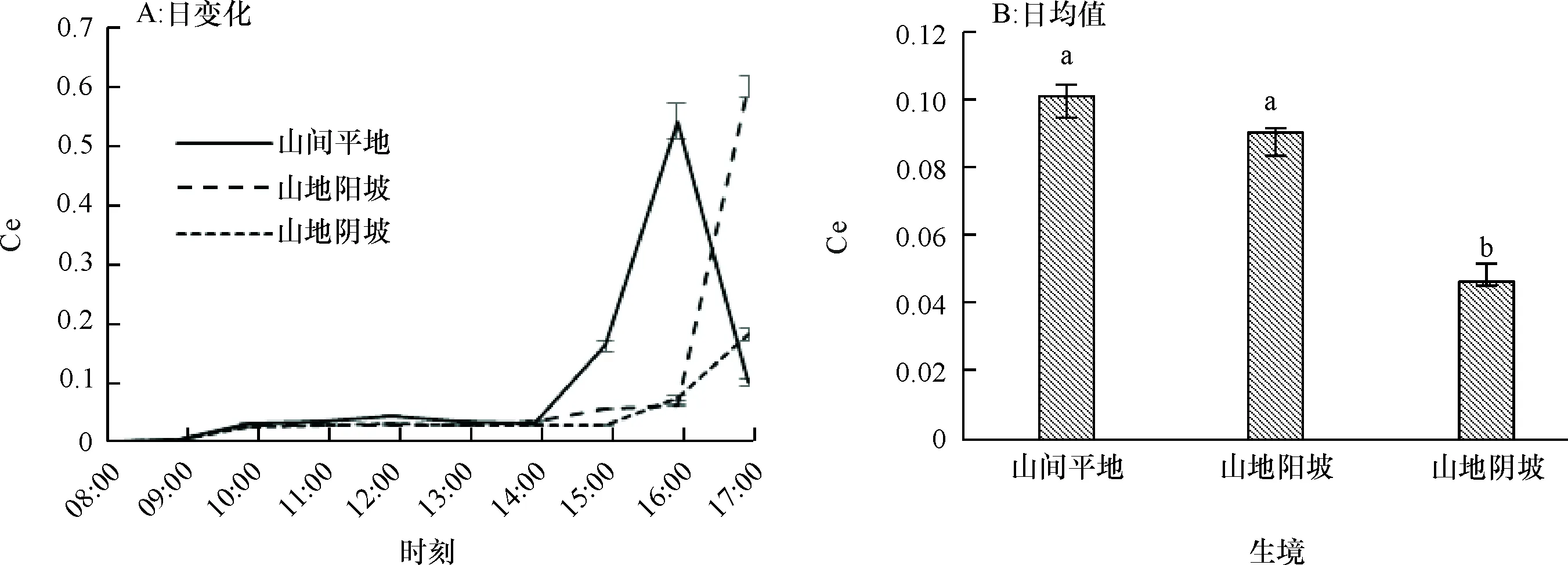
图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.8 光能利用效率日变化 光能利用效率(SUE)反映不同生境条件下赤皮青冈对于光照辐射利用潜能的大小,从图8A看出,在不同的生境条件下,赤皮青冈的光能利用效率日变化均呈现典型的W型趋势,即在日出和中午光合午休时的光能利用效率最低,其他时间的光能利用效率较高。在光能利用效率较高的10:00—12:00及15:00—17:00时,光能利用效率表现为山间平地>山地阳坡>山地阴坡生境。光能利用效率的日均值(图8B)也表现出山地生境低于平地生境,特别是山地阴坡生境条件下光能利用效率显著降低(P为0.049),较平地生境低了10.14%。
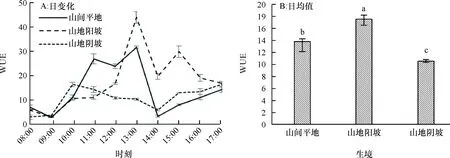
2.1.9 水分利用效率日变化 山间平地和山地阳坡生境下的赤皮青冈水分利用效率(WUE)日变化均表现为9:00略有降低,然后迅速增加,在13:00时最高,14:00时迅速下降的规律,同时山间平地生境下赤皮青冈上午的水分利用效率总体高于下午时段,而山地阳坡生境下的水分利用效率下午总体高于上午时段;山地阴坡生境下赤皮青冈的水分利用效率表现出同其他生境下不同的趋势,即10:00达到最大值后开始下降,14:00又开始缓慢上升的变化趋势。从图9B可以看出,生境条件对水分利用效率日均值的影响为:山地阳坡>山间平地>山地阴坡,山间平地和山地阴坡生境下的水分利用效率分别比山地阳坡生境低20.97%和39.82%,且差异显著(P为0.022和0.010)。

图B中, 柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
图B中,柱子上方凡有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
2.1.10 光合指标日变化的相关性分析 根据不同生境条件下光合参数与净光合速率的Pearson相关性分析(表2),除胞间CO2浓度在不同生境下与净光合速率呈非显著的负相关关系外,其他光合参数均与净光合速率呈正相关关系。特别是光合有效辐射、气孔导度和叶片温度,在不同生境条件下均与净光合速率呈显著的正相关关系(P为0.050~0.001),分析光合参数与净光合速率的相关性发现:在山间平地生境下的相关系数表现为θL(0.793)> PAR (0.680)> Gs(0.631),在山地阳坡生境下的相关系数表现为Gs(0.845)>θL(0.822)>PAR(0.798),在山地阴坡生境下的相关系数表现为θL(0.861)>PAR(0.729)>Gs(0.634),蒸腾速率、气孔限制值和瞬时羧化效率对不同生境下赤皮青冈的光合速率影响最小。
2.2 光响应曲线
从图10可以看出,山地阴坡生境下赤皮青冈的净光合速率和光饱和点明显低于其他生境条件;山间平地生境下赤皮青冈的净光合速率和光饱和点与山地阳坡生境时相近。结合表3数据可以看出,山地阴坡生境下赤皮青冈的光补偿点仅为2.257 μmol·m-2·s-1,而山地阳坡和山间平地生境下赤皮青冈的光补偿点均在51 μmol·m-2·s-1左右,说明赤皮青冈对长期阴坡的生境产生了适应性,能充分利用弱光进行光合作用;光饱和点(Lsp)表现为山间平地>山地阳坡>山地阴坡,山地生境分别比平地生境的光饱和点低了18.96%和53.25%,差异显著(P为0.047和0.001),反映出山地生境提高了赤皮青冈对光合有效辐射的利用能力。光补偿点低的植物能在较低的光照强度下进行光合作用,而光饱和点高的植物在较高光照强度下不会出现光合午休现象,表明植物具有较宽的利用光照辐射的能力。对于任何一种环境,如果植物具有较高的Pnmax、Lsp、AQY和较低的Lcp,可预测该植物具有更宽的光照适应性。赤皮青冈表观量子效率(AQY)的大小表现为山间平地>山地阴坡>山地阳坡,这表明长期生长在山间平地生境条件下的赤皮青冈对光照辐射的适应性广,既有较强的弱光利用能力,又有较强的强光利用潜力。而对于山地阴坡生境下的赤皮青冈来说,其具有很强的弱光利用能力,但是只有较弱的强光利用潜力;山地阳坡与山间平地生境下的赤皮青冈光合能力表现相似,但对光合辐射的利用能力弱于山间平地。
表2 不同生境下赤皮青冈光合指标与净光合速率相关系数1)
Tab.2 Correlation coefficients of photosynthetic indexes and net photosynthetic rates (Pn) ofCyclobalanopsisgilvain different habitats
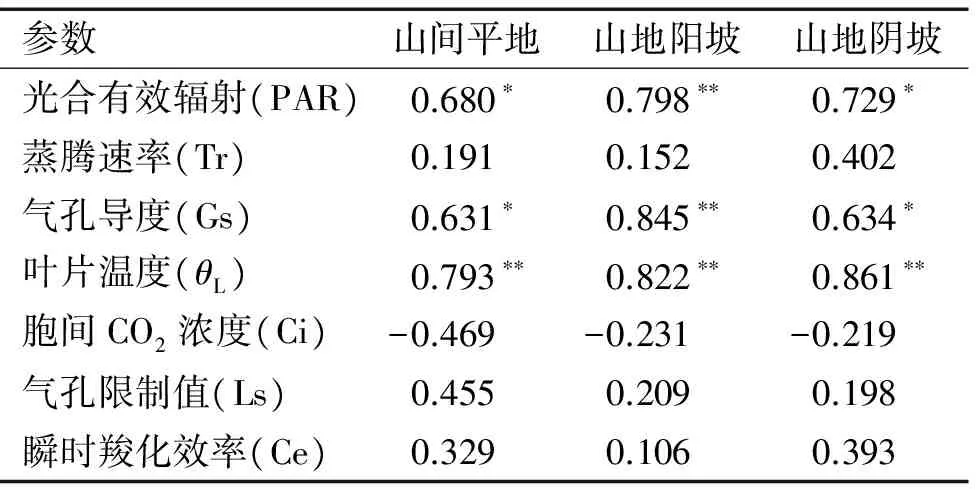
参数山间平地山地阳坡山地阴坡光合有效辐射(PAR)0.680∗0.798∗∗0.729∗蒸腾速率(Tr)0.1910.1520.402气孔导度(Gs)0.631∗0.845∗∗0.634∗叶片温度(θL)0.793∗∗0.822∗∗0.861∗∗胞间CO2浓度(Ci)-0.469-0.231-0.219气孔限制值(Ls)0.4550.2090.198瞬时羧化效率(Ce)0.3290.1060.393
1)*和**分别表示0.05和0.01水平显著相关(Pearson法)。
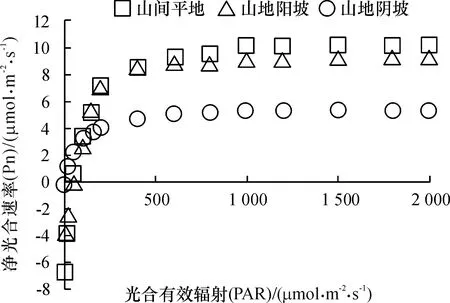
图10 不同生境条件下赤皮青冈的光响应曲线
Fig.10 Light response curves ofCyclobalanopsisgilvain different habitats
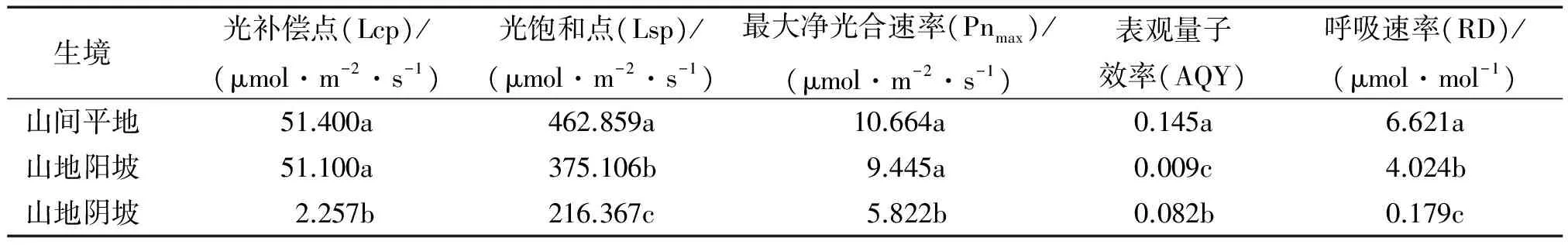
表3 不同生境下赤皮青冈的光响应曲线特征参数1)
1)同列数据后凡是有一个相同小写字母者,表示不同生境间在0.05水平差异不显著(LSD法)。
3 讨论与结论
赤皮青冈光合作用的强弱是其生存和生长的基础,也决定了其健康状况和物质积累,而光合速率的大小不仅与赤皮青冈本身的遗传特性有关,也受光照、温度等环境因素的密切影响[22];长期生长在不同生境条件下的赤皮青冈,逐渐演替出对环境的适应性。赤皮青冈在山间平地和山地阳坡生境条件下经过对强光照和长日照的长期适应,具有较高的净光合速率,同时为减少强光照辐射带来的高温和干旱等不利因素的影响,叶片通过调整气孔的开闭控制蒸腾速率的大小和胞间CO2浓度的高低,使叶片温度的变化变缓,从而减少强光辐射带来的高温伤害和生理干旱,实现净光合速率的最大化,表现出同樟树Cinnamomumbodinieri等阔叶树种类似的生理反应[23];而山地阴坡生境下的赤皮青冈适应了光照少和温度低的环境,由于光照辐射对叶片温度有直接的影响,因此促使赤皮青冈在阴坡环境下形成了较快的温度响应能力,但从净光合速率的双峰型趋势来分析,山地阴坡生境较其他生境条件下的赤皮青冈光合午休现象不明显,说明其光合午休的形成和波动是由非气孔因素造成的[24-27]。不同生境下赤皮青冈的净光合速率变化不大,但山地生境下的胞间CO2浓度增加且气孔限制值明显下降,表明不同生境下赤皮青冈光合能力的差异是由非气孔因素造成的。净光合速率在山地环境下比平地环境略有下降,说明山地环境导致赤皮青冈的叶绿体、Rubisco活性下降,RuBP羧化酶再生能力也下降,从而导致赤皮青冈光合能力的变化,但生境对赤皮青冈光合能力影响的大小还有待进一步的研究,特别是不同的土壤水分和肥力状况可能引起植物RuBP羧化酶活性变化[28-29],对特定土壤条件的长期适应可对植物的光合生理产生直接的影响。
对比分析发现,山间平地、山地阳坡生境条件下赤皮青冈的净光合速率较高,但对光照强度反应的敏感程度不如山地阴坡生境下的赤皮青冈,长期生长在山地阴坡生境条件下的赤皮青冈较其他2种生境条件下的赤皮青冈更耐受低光照条件。植物光补偿点的高低是植物耐荫性评价的重要指标,不同生境条件下的赤皮青冈需光性介于阴生植物和阳生植物之间,山间平地和山地阳坡生境下的赤皮青冈对强光的适应能力较好,特别是山间平地生境下的赤皮青冈对不同强度的光照辐射表现出较宽的适应性,既能充分利用弱光进行光合作用,也能对强光有较高的利用能力,光照生态幅较宽,其潜在的光合作用较强,这些光合能力的差异是树木对种源的长期适应及遗传选择的结果[23-24]。
[1] 郑元, 唐军荣, 王连春, 等. 干热河谷牛角瓜苗期光合生理特性的种源间差异[J]. 浙江农林大学学报, 2016, 33(3): 440-448.
[2] 杜旭华, 丁兴萃, 陈岩, 等. 不同纬度引种地马来甜龙竹(Dendrocalamusasper)光合特性的比较[J]. 南京林业大学学报(自然科学版), 2012, 36(6): 53-57.
[3] 刘嘉君, 王志刚, 阎爱华, 等. 12种彩叶树种光合特性及固碳释氧功能[J]. 东北林业大学学报, 2011, 39 (9): 23-25.
[4] 管铭, 金则新, 李月灵, 等. 千岛湖次生林优势种植物光合生理生态特性[J]. 生态学报, 2015, 35(7):2057-2066.
[5] 谭晓红, 王爽, 马履一, 等. 豫西刺槐能源林培育的光合生理生态理论基础[J]. 生态学报, 2010, 30(11): 2940-2948.
[6] 许怡玲, 遇文婧, 宋小双, 等. 6种玉簪耐阴性分析[J]. 东北林业大学学报, 2012, 40(11): 31-34.
[7] 王旭军, 吴际友, 廖德志, 等. 长沙市8种常见绿化树种的耐阴性[J]. 东北林业大学学报, 2010, 38(3): 14-16.
[8] 蒋冬月, 钱永强, 费英杰, 等. 柳属植物光合-光响应曲线模型拟合[J]. 核农学报, 2015, 29(1): 169-177.
[9] 韩刚, 赵忠. 不同土壤水分下4 种沙生灌木的光合光响应特性[J]. 生态学报, 2010, 30(15): 4019-4026.
[10]洪伟, 杨细明, 林勇明, 等. 不同光合光响应模型在雷公藤研究中的应用[J]. 福建农林大学学报(自然科学版), 2012, 41(1): 29-33.
[11]祁承经, 林亲众. 湖南树木志[M]. 长沙: 湖南科学技术出版社, 2000:376.
[12]吴小林, 张东北, 楚秀丽, 等. 赤皮青冈容器苗不同基质配比和缓释肥施用量的生长效应[J]. 林业科学研究, 2014, 27(6): 794-800.
[13]赵嫦妮, 徐德禄, 李志辉. 配方施肥对赤皮青冈容器苗生长的影响[J]. 中南林业科技大学学报, 2013, 33(5): 22-25.
[14]吴丽君, 李志辉, 杨模华, 等. 赤皮青冈幼苗叶片解剖结构对干旱胁迫的响应[J]. 应用生态学报, 2015, 26(12): 3619-3626.
[15]朱品红, 李志辉, 杨模华, 等. 赤皮青冈ISSR-PCR反应体系的建立与优化[J]. 中南林业科技大学学报, 2014, 34(6): 61-65.
[16]吴丽君, 李志辉, 赵嫦妮. 赤皮青冈光合作用的日变化研究[J]. 江西林业科技, 2011, 39(5): 1-3.
[17]吴丽君, 李志辉, 邹峰. 水分胁迫对珍贵树种赤皮青冈幼苗生理特性的影响[J]. 中南林业科技大学学报, 2012, 32(7): 10-13.
[18]李金华, 赵嫦妮, 徐德禄, 等. 浙江宁波赤皮青冈不同家系幼苗光合特性差异分析[J]. 中国农学通报, 2013, 29(22):6-9.
[19]王明怀, 许秀玉, 魏龙, 等. 木麻黄无性系光响应特性研究[J]. 华南农业大学学报, 2011, 32(2):76-79.
[20]FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis[J]. Plant Biol, 1982, 33(33):317-345.
[21]HERRERA A, TEZARA W, MARíN O, et al. Stomatal and non-stomatal limitations of photosynthesis in trees of a tropical seasonally flooded fores[J]. Physiol Plant, 2008, 134(1):41-48.
[22]薛雪, 李娟娟, 郑云峰, 等. 5个常绿园林树种的夏季光合蒸腾特性[J]. 林业科学, 2015, 51(9):150-155.
[23]胡启鹏, 孙玲玲, 郭志华, 等. 常绿阔叶林中几个不同功能组树种叶形态、光合及光诱导对不同光环境的响应[J]. 华南农业大学学报, 2013, 34(2):207-212.
[24]全先奎, 王传宽. 兴安落叶松对环境变化的物候驯化和光合能力适应[J]. 生态学报, 2015, 35(13):4538-4546.
[25]周祥斌, 周玮, 邓丽婷, 等. 14个香椿种源光合特性的比较研究[J]. 华南农业大学学报, 2016, 37(4):51-56.
[26]段爱国, 张建国, 何彩云, 等. 干热河谷主要造林树种气体交换特性的坡位效应[J]. 生态学报, 2013, 33(12): 3630-3638.
[27]JI D H, MAO Q Z, WATANABE Y, et al. Effect of nitrogen loading on the growth and photosynthetic responses of Japanese larch seedlings grown under different light regimes[J]. J Agric Meteorol, 2015, 71(3):232-238.
[28]赵秀莲, 夏新莉, 尹伟伦, 等. 不同苗龄沙地柏抗旱生理特性比较研究[J]. 西北植物学报, 2013, 33(12): 2513-2520.
[29]李泽, 谭晓风, 卢锟, 等. 供镁水平对油桐幼苗生长及光合特性的影响[J]. 生态学杂志, 2015, 34(9): 2440-2447.
【责任编辑 李晓卉】
Effects of habitats on photosynthesis characteristics ofCyclobalanopsisgilva
ZHANG Bin1, ZHU Ninghua1, CAO Jiwu2
(1 College of Forestry, Central South University of Forestry and Technology, Changsha 410004, China;2 Botanical Garden, Central South University of Forestry and Technology, Changsha 410004, China)
【Objective】 To analyze the photosynthetic adaptations ofCyclobalanopsisgilvato different habitats, and provide theoretical and data supports for scientific breeding and cultivation.【Method】Three natural populations ofC.gilvain different habitats were studied. Diurnal changes in photosynthetic parameters ofC.gilvaunder natural condition were measured using LI-6400 portable photosynthesis system. The light response curves were obtained.【Result】Under different habitats, net photosynthetic rate(Pn) and transpiration rate(Tr) ofC.gilvahad similar changing patterns, both reached peak at 10:00 and 14:00, and were lowest at 13:00. Leaf temperature (θL), photosynthetically active radiation(PAR), and stomatal conductance(Gs) were all strongly positively correlated with Pn. The ability of using light forC.gilvawas influenced by habitat as shown by the light response curves;C.gilvagrown on flat between mountains had the best ability of using light, followed byC.gilvagrown on southern slope of mountain, and then northern slope of mountain. Light compensation points (Lcp) were low in all three habitats. Lcp was the lowest (2.257 mol·m-2·s-1) forC.gilvagrown on northern slope of mountain, and around 51 mol·m-2·s-1forC.gilvagrown on flat between mountains and southern slope of mountain. Light saturation point (Lsp) was the highest forC.gilvagrown on flat between mountains, followed byC.gilvagrown on southern slope of mountain, and then northern slope of mountain.【Conclusion】C.gilvahad strong adaptability to all three habitats.C.gilvagrown on flat between mountains and southern slope of mountain have adapted to longer light duration and higher light intensity, and have strong photosynthetic capacity and ability of using hight light intensity.C.gilvagrown on southern slope of mountain have the ability of using weak light intensity. Long adaptation to different habitats was the main cause of photosynthetic capacity differences ofC.gilva.
precious tree; photosynthetic characteristic; habitat;Cyclobalanopsisgilva
2016- 06- 21 优先出版时间:2017-04-12
张 斌(1980—),男,讲师,博士研究生,E-mail: zhangbin@csuft.edu.cn
国家科技支撑项目( 2012BAD21B03)
Q945.11
A
1001- 411X(2017)03- 0070- 09
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.s.20170412.1413.002.html
张 斌, 朱宁华, 曹基武.不同生境对赤皮青冈光合特性的影响[J].华南农业大学学报,2017,38(3):70- 78.
