不同耐盐性玉米自交系苗期耐旱性的比较
2017-05-02杨晓杰张晶孙百良王瑶��
杨晓杰++张晶+孙百良+王瑶+��
摘要:以2份玉米自交系经3-1、05Ny-10为材料,设计4个不同强度的干旱胁迫,研究干旱胁迫对不同耐盐性[JP2]玉米自交系苗期形态及生理生化指标的影响。结果显示,在干旱胁迫下,2份玉米自交系幼苗的根粗、根长、须根数、株高、茎粗、叶宽、叶长、含水量、色素含量基本呈递减趋势;可溶性糖、可溶性蛋白、脯氨酸含量等抗逆性物质含量呈递增趋势;相对电导率、丙二醛含量均递增。综合形态及生理生化指标,不耐盐的自交系经3-1的耐旱性优于05Ny-10。[JP]
关键词:耐盐玉米;自交系;耐旱性;生理生化指标
中图分类号: S513.01文献标志码: A
文章编号:1002-1302(2017)03-0041-03
收稿日期:2015-09-28
基金项目:黑龙江省教育厅科学研究技术研究项目(编号:12541875)。
作者简介:杨晓杰(1962—),女,辽宁法库人,硕士,教授,研究方向为植物发育与资源植物研究。E-mail:yangxiaojie1980@163.com。
干旱是影响我国及世界玉米穩产的重要环境因子,每年因干旱造成玉米减产约20%~30%,严重旱灾年部分地区几乎绝收,干旱已经成为影响玉米产量的重要限制因素[1-2]。随着全球气候变暖和水资源的日益匮乏,有关玉米抗旱性研究及其应用更加受到重视。挖掘遗传潜力、选育抗旱玉米种质资源是提高玉米产量稳定性、抵御干旱的重要途径之一。国内外学者对玉米耐旱性研究已经作了大量工作[3-9],认为苗期耐旱性与成株期的耐旱性显著相关。植物在受到干旱胁迫后,会诱发植物体内其他多种不良反应,因而植物必须迅速启动对这些不良反应的应答以存活[10]。
但是目前以不同耐盐性玉米自交系为材料,研究其耐旱性机制的报道较少。[JP+1]本试验在前期玉米自交系耐盐性筛选的基础上,选用耐盐性较强的玉米自交系05Ny-10和耐盐性差的经3-1[11],比较2份玉米自交系的耐旱性,分析耐盐与耐旱的一致性,为筛选耐盐与耐旱玉米自交系提供试验依据。[JP]
1材料与方法
1.1供试材料
2种不同耐盐性玉米自交系:经3-1、05Ny-10的籽粒,每份籽粒均100粒。
1.2试验方法
挑选饱满且大小均匀一致的玉米籽粒,浸种后于33 ℃恒温箱培养催芽,将发芽的玉米种子种入排水良好的沙盘中,置于室温(29±1) ℃培养。用控制浇水频率的方法分为对照组(CK)、轻度干旱胁迫(LS)、中度干旱胁迫(MS)和重度干旱胁迫(SS)4个处理组。在预试验的基础上,以最适宜的每天喷水处理为对照组,其余为每3、5、7 d喷水,采用喷淋的方式各组喷等量的水,进行胁迫处理,处理第15天时开始测定各形态指标和各生理生化指标。
1.3指标测定
1.3.1形态指标测定
测定玉米的根粗、根长、株高、茎粗、叶长、叶宽和须根数等形态指标,每个处理重复3次(n=3),每个重复5株,数据以“平均值±标准差”表示。
1.3.2生理生化指标测定
色素含量、脯氨酸含量、丙二醛(MDA)含量测定参照张宪政的方法[12];相对电导率、可溶性糖含量、可溶性蛋白含量测定参照白宝璋等的方法[13]。
1.4数据统计
数据处理采用Excel 2003软件统计分析。
2结果与分析
2.1干旱对玉米自交系形态指标的影响
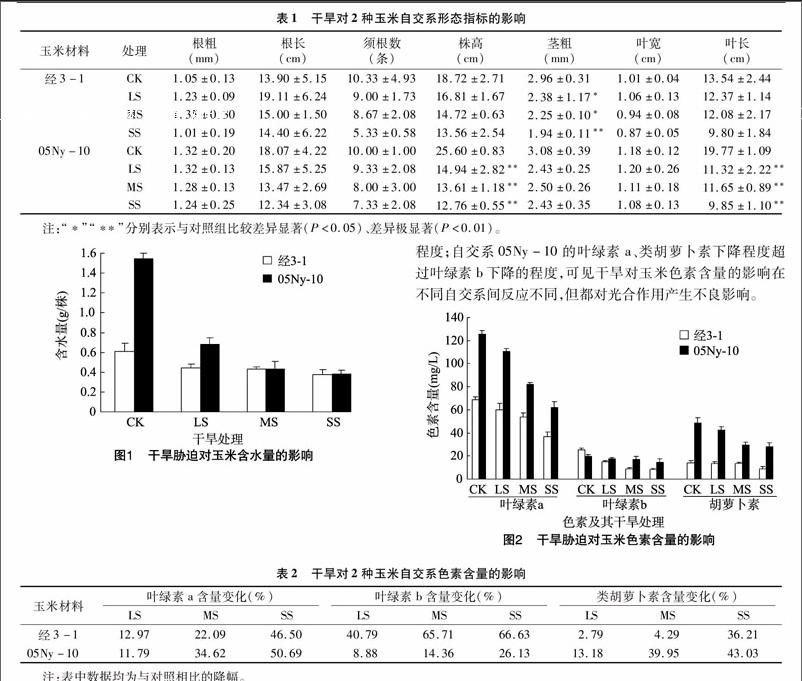
不同干旱条件对玉米自交系形态的影响,由表1可知,CK组的玉米自交系05Ny-10的根粗、根长、株高、茎粗、叶宽和叶长明显高于自交系经3-1,即自然生长条件下,二者存在明显差异。
随着干旱胁迫的加剧,2份玉米的根粗、须根数、株高、茎粗、叶宽和叶长基本呈递减趋势。由表1可见,自交系 05Ny-10的各个干旱胁迫组株高下降明显,与对照比差异极显著(P<0.01),说明干旱胁迫导致植株矮化,生长受阻,而茎粗变化不显著;自交系经3-1的株高下降不明显,而茎粗下降明显(LS、MS组P<0.05,SS组P<0.01);2份自交系各个处理组叶宽变化不大;自交系05Ny-10的各个干旱胁迫组叶长极显著变短(P<0.01)。2份自交系在干旱胁迫时,根长变化趋势不同,自交系经3-1干旱处理组比对照组的根长都有所增加,其抵御干旱的途径之一是通过增加根的长度向土壤深处吸收更多的水分,随干旱程度的增加根长增加量逐渐减小;自交系05Ny-10干旱处理组根长均小于对照组,随干旱胁迫程度的增加,根长变短的幅度增大。
2.2干旱对玉米自交系含水量的影响
由图1可见,随干旱加剧,2个自交系含水量都明显下降,自交系经3-1的LS、MS、SS处理组分别下降了27.62%、29.34%、38.34%,自交系05Ny-10分别下降了55.95%、72.10%、75.40%,可见自交系经3-1的持水能力较强。
2.3干旱对玉米自交系色素含量的影响
由图2、表2可知,叶绿素a、叶绿素b和类胡萝卜素3种色素均随着干旱胁迫的加剧呈明显递减趋势,自交系经3-1的叶绿素a、叶绿素b含量下降程度超过类胡萝卜素下降的程度;自交系05Ny-10的叶绿素a、类胡萝卜素下降程度超过叶绿素b下降的程度,可见干旱对玉米色素含量的影响在不同自交系间反应不同,但都对光合作用产生不良影响。
2.4干旱对玉米自交系叶片质膜透性的影响
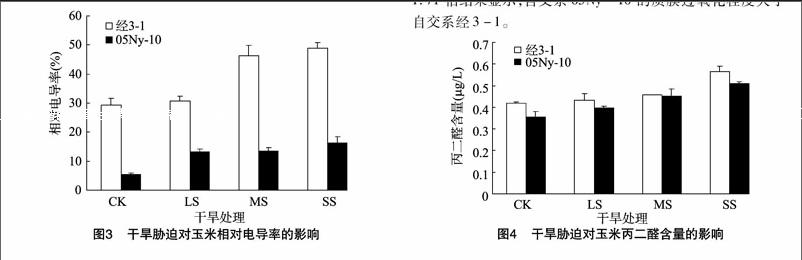
质膜的伤害程度用相对电导率表示。由图3可见,随着干旱程度的加剧,2份玉米自交系相对电导率均呈升高的趋势,自交系经3-1的3个干旱胁迫组(LS、MS、SS)分别是对照的 1.05、1.59、1.68倍,自交系05Ny-10的3个干旱胁迫组(LS、MS、SS)分别是对照的2.46、2.48、3.03倍,表明自交系05Ny-10的质膜受到的伤害更大。
2.5对丙二醛含量的影响
在逆境胁迫下,由于自由基对细胞膜的过氧化伤害产生了丙二醛。丙二醛含量的变化能够反映质膜的过氧化的程度。由图4可见,2个玉米自交系在干旱胁迫下,丙二醛含量均呈明显递增趋势。自交系经3-1的3个干旱胁迫组(LS、MS、SS)分别是对照的1.03、1.09、1.35倍,自交系05Ny-10的3个干旱胁迫组(LS、MS、SS)分别是对照的1.12、1.27、 1.71 倍结果显示,自交系05Ny-10的质膜过氧化程度大于自交系经3-1。
2.6干旱对玉米自交系渗透调节物质含量的影响
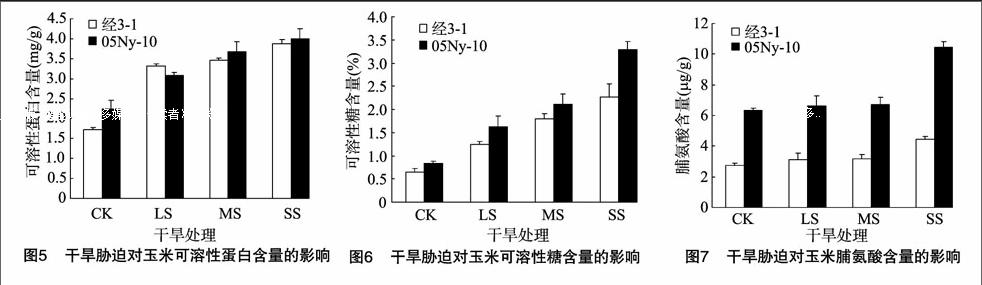
可溶性糖、可溶性蛋白、脯氨酸等是植物在遭受逆境胁迫时起渗透调节作用的调节剂。植物遭受干旱时,可以通过产生上述物质调节体内的渗透压以抵御干旱。2个玉米自交系在干旱胁迫下,3种物质含量都呈递增的趋势(图5至图7)。自交系经3-1 LS、MS、SS处理的可溶性糖含量分别是对照的1.95、2.81、3.53倍,可溶性蛋白含量分别是对照的1.92、2.02、2.26倍(图5),脯氨酸含量分别是对照的1.15、1.42、1.63倍(图7);自交系05Ny-10的可溶性糖含量分别是对照的194、2.53、3.95倍,可溶性蛋白含量分别是对照的138、1.64、1.78倍,脯氨酸含量分别是对照的1.05、1.06、165倍(圖5至图7)。2个自交系的可溶性糖、脯氨酸含量变化差异不大,自交系经3-1可溶性蛋白含量增幅明显高于自交系05Ny-10,即在渗透调节物积累能力上,自交系经 3-1 强于自交系05Ny-10。[FL)]
[TPYXJ5.tif][FK)]
3结论与讨论
抗旱性是植物在环境因素作用下,表现出的形态特征和生理生化特性变化的复杂性状。研究表明,抗旱性强的植物具有良好的抵御和适应不良环境的生长策略[14],干旱胁迫时,在其他形态指标都降低的情况下,自交系经3-1通过增加根长的策略,增加吸收土壤深处水分的能力,以保证体内生长过程中对水分的需求。
丙二醛含量和相对电导率的增加能反映膜受伤害的程度,在干旱胁迫下丙二醛含量增加说明膜受到的过氧化伤害大。自交系经3-1丙二醛的含量和相对电导率的增加幅度均小于自交系05Ny-10,说明在干旱胁迫时,经3-1受到的过氧化伤害低于自交系05Ny-10。
渗透调节物质可以使玉米在受到逆境时维持一定的压力,保证各生理功能的顺利进行。渗透调节物质积累程度是衡量作物抗旱能力的重要指标。在干旱胁迫下,抗旱的自交系积累的渗透调节物质相对较多,不抗旱的自交系积累渗透调节物质则较少。不同的玉米自交系对逆境胁迫的形态和生理响应是不同的,不应单纯以单个指标判别其抗旱性的强弱,要综合形态特征和生理特征综合分析。在正常生长条件下,自交系05Ny-10的各项形态指标都好于自交系经3-1,且05Ny-10的耐盐性强于经3-1,但在受到干旱胁迫时,经 3-1的脯氨酸含量、可溶性糖含量、可溶性蛋白含量的相对增幅要高于05Ny-10。结合根长等形态指标、含水量及其他生理指标综合分析,自交系经3-1在干旱胁迫下的调节能力要强于05Ny-10,表现出较好的抗旱性,为筛选玉米抗旱资源提供了参考依据,说明不同自交系的抗盐、抗旱机制是有差异的,还有待对其差异的产生机制作进一步的研究。
[HS2]参考文献:
[1]Dai A G,Trenberth K E,Qian T T. A global dataset of Palmer Drought Severity Index for 1870-2002:relationship with soil moisture and effects of surface warming[J]. J Hydrometeor,2009,5(6):1117-1130.[HJ1.75mm]
[2]Nicholls N. The changing nature of Australian droughts[J]. Climatic Change,2004,63(3):323-336.
[3]张宝石,徐世昌,宋凤斌,等. 玉米抗旱基因型鉴定方法和指标的探讨[J]. 玉米科学,1996,4(3):19-22.
[4]Belo A,Zheng P Z,Luck S,et al. Whole genome scan detects an allelic variant of fad2 associated with increased oleic acid levels in maize[J]. Molecular Genetics and Genomics,2008,279(1):1-10.
[5]Bernardo R,Yu J. Prospects for genomewide selection for quantitative traits in maize[J]. Crop Science,2007,47:1082-1090.
[6]Bolanos J,Edmeades G. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize[J]. Field Crops Res,1996,48:65-80.
[7]Harjes C E,Rocheford T R,Bai L,et al. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification[J]. Science,2008,319(5861):330-333.
[8]苏治军,郝转芳,谢传晓,等. 我国常用玉米自交系的耐旱性评价[J]. 玉米科学,2009,17(5):19-24.
[9]李晚忱,付凤玲,袁佐清. 玉米苗期耐旱性鉴定方法研究[J]. 西南农业学报,2001,14(3):29-32.
[10]徐立明,张振葆,梁晓玲,等. 植物抗旱基因工程研究进展[J]. 草业学报,2014,23(6):293-303.
[11]杨晓杰,李旭业,王海艳,等. 玉米自交系耐盐种质的筛选及芽苗期耐盐性评价[J]. 玉米科学,2014,22(4):19-25.
[12]张宪政. 作物生理研究法[M]. 北京:农业出版社,1992:119-218.
[13]白宝璋,王景安,孙玉霞,等. 植物生理学测试技术[M]. 北京:中国科技出版社,1993:148-156.
[14]黄彩变,曾凡江,雷加强. 塔克拉玛干沙漠南缘3个沙拐枣种的抗旱特性比较[J]. 草业学报,2014,23(3):136-143.
