香蕉茎叶粉固态发酵条件优化及鹅对其养分利用率的研究
2017-04-24王增煌王文策翟双双
王增煌 王文策 翟双双 谢 强 左 鑫 杨 琳
(华南农业大学动物科学学院,广州510642)
香蕉茎叶粉固态发酵条件优化及鹅对其养分利用率的研究
王增煌 王文策 翟双双 谢 强 左 鑫 杨 琳*
(华南农业大学动物科学学院,广州510642)
本试验旨在研究应用固态发酵技术改善香蕉茎叶粉养分组成的工艺参数及鹅对其养分的利用率。选择米曲霉和产朊假丝酵母作为发酵菌种,逐步探究5个因素(单菌种发酵、硫酸铵添加量、米曲霉和产朊假丝酵母接种比例、混合菌液接种量)对发酵后香蕉茎叶粉蛋白质含量的影响。利用正交设计筛选最优发酵温度、底物水分和发酵时间的组合。结果显示:米曲霉和产朊假丝酵母单独发酵均能显著或极显著提高发酵后香蕉茎叶粉的粗蛋白质含量(P<0.05或P<0.01)。添加硫酸铵能显著或极显著提高发酵后香蕉茎叶粉的真蛋白质含量(P<0.05或P<0.01),其中添加2%硫酸铵组的真蛋白质含量最高。米曲霉∶产朊假丝酵母接种比例为2∶1的处理发酵后香蕉茎叶粉的蛋白质净增加量显著或极显著高于比例为1∶1、1∶3和3∶2的处理(P<0.05或P<0.01)。正交试验结果显示,以4%的接种量、接种比例为2∶1(米曲霉∶产朊假丝酵母)、2%的硫酸铵添加量,在基质水分为50%,30 ℃的环境下发酵4 d效果最佳。经过该工艺发酵后的香蕉茎叶粉粗蛋白质含量提高了33.82%,氨基酸分析结果显示,除赖氨酸和精氨酸外其余的15种氨基酸含量均有不同程度的提高。马冈鹅的代谢试验结果显示,发酵后香蕉茎叶粉中的粗蛋白质利用率提高了52.66%,极显著高于发酵前(P<0.01);此外,代谢能和能量利用率也都略有提高(P>0.05)。由此可见,经过该发酵工艺发酵后的香蕉茎叶粉营养价值不仅得到了改善,也促进了鹅对其养分的消化吸收。
香蕉茎叶粉;固态发酵;蛋白质;养分利用率;马冈鹅;米曲霉;产朊假丝酵母
我国饲料资源短缺,开发和利用非常规饲料资源以替代传统的饲料原料是行业关注的热点之一。据统计,中国香蕉年产量逾1×107t,香蕉茎叶是香蕉生产过程当中的副产物,数量庞大,其可溶性糖含量高,矿物质含量丰富,含有B族维生素和胡萝卜素等养分,有一定的饲用价值,具备成为饲料资源的潜力。当前缺乏香蕉茎叶饲料化加工技术的研究,没有配套的设备,加上香蕉茎叶中蛋白质含量低的缺陷,导致了不能物尽其用的局面,如果能将其充分应用于饲料中,不仅可降低饲料成本,还可以缓解饲料资源紧张的问题[1]。
大量研究证实,固态发酵(solid state fermentation,SSF)技术具有能耗低、设备较简易、不产生废水等特点,应用该发酵技术将玉米秸秆、苹果渣、豆渣[2-3]等废弃物转化为微生物蛋白质资源的研究已取得了较好的成果。本试验选择米曲霉(Aspergillusoryzae)和产朊假丝酵母(Candidautilis)作为菌种,探索利用这2种微生物固态发酵香蕉茎叶粉的适宜条件,并通过马冈鹅的代谢试验比较发酵前后香蕉茎叶粉的养分利用情况,为微生物处理香蕉茎叶粉提供数据参考。
1 材料与方法
1.1 试验菌种
米曲霉GIM 3.545和产朊假丝酵母购于广东省微生物菌种保藏中心。
1.2 试验材料
香蕉茎叶(叶片和叶柄后端的一小段假茎)采自广州天河长湴村香蕉园,经过晒干粉碎过40目筛。硫酸铵(分析纯)、马铃薯葡萄糖琼脂培养基(PDA)、麦芽提取物粉、酵母浸膏葡萄糖琼脂培养基,购于广州维宁生物有限公司,使用前按使用量称取相应的培养基粉末溶解于蒸馏水后置于121 ℃高压蒸汽灭菌锅灭菌20 min后待用。
1.3 试验方法
1.3.1 米曲霉孢子悬液的制备
用马铃薯葡萄糖琼脂培养基培养米曲霉7 d后,用无菌水将米曲霉孢子冲洗入锥形瓶中,经过4层纱布过滤后放入摇床28 ℃、200 r/min摇4 h打散孢子,用平板菌落计数法计算孢子悬液浓度以确保制备的孢子悬液浓度≥1×108个/mL。
1.3.2 产朊假丝酵母菌液的制备
用麦芽提取物粉配制产朊假丝酵母液体培养基,在28 ℃、200 r/min摇床中培养24 h后,用平板菌落计数法计算产朊假丝酵母菌液浓度以确保菌液浓度≥1×108个/mL。
1.3.3 单菌种发酵
称取15 g香蕉茎叶粉于容积为480 mL的玻璃罐中,封口膜封口后用121 ℃高压灭菌锅灭菌20 min,待用。将事先制作好的菌液以3%的接种量接种入罐中,调节底物水分为50%,自然pH,在28 ℃恒温培养箱中发酵60 h,每隔12 h摇瓶以确保菌株正常生长,每株菌种6个发酵罐。发酵产物用65 ℃烘箱烘干,测定粗蛋白质含量。
1.3.4 双菌种发酵
1.3.4.1 硫酸铵添加量的确定
称取15 g香蕉茎叶粉于发酵罐中并加入不同质量(0、0.5%、1.0%、1.5%、2.0%、2.5%,占香蕉茎叶粉的比重)的硫酸铵,封口膜封口后在121 ℃高温灭菌锅中灭菌20 min,待用。向发酵罐内接入米曲霉∶产朊假丝酵母比例为1∶1的菌液,接种量3%,调节底物水分为50%,28 ℃恒温培养箱发酵60 h,每隔12 h摇瓶以确保菌株正常生长,每种硫酸铵添加量设置6个发酵罐。发酵产物用65 ℃烘箱烘干,测定真蛋白质含量。
1.3.4.2 菌种比例的筛选
称取15 g香蕉茎叶粉于发酵罐中,硫酸铵根据硫酸铵添加量的确定结果添加,封口膜封口后用121 ℃高温灭菌锅灭菌20 min,待用。向发酵罐内接入米曲霉:产朊假丝酵母菌数不同比例(1∶1、1∶3、3∶2、2∶3、2∶1)的混合菌液,接种量3%,调节底物水分为50%,28 ℃恒温培养箱发酵60 h,每隔12 h摇瓶以确保菌株正常生长,每种比例设置6个发酵罐。发酵产物用65 ℃烘箱烘干,测定真蛋白质含量。
1.3.4.3 混合菌液接种量的选择
称取15 g香蕉茎叶粉于发酵罐中,硫酸铵根据硫酸铵添加量的确定结果添加,封口膜封口后用121 ℃高温灭菌锅灭菌20 min,待用。向发酵罐内接入不同质量(2%、3%、4%、5%、6%,占香蕉茎叶粉的比重)的混合菌液,2种菌种比例依照菌种接种比例的筛选结果确定,调节底物水分为50%,28 ℃恒温培养箱发酵60 h,每隔12 h摇瓶以确保菌株正常生长,每种硫酸铵添加量设置6个发酵罐。发酵产物用65 ℃烘箱烘干,测定真蛋白质含量。
1.3.5 正交试验设计
根据确定的硫酸铵添加量、菌种比例和混合菌液接种量为基础,在该发酵条件下优化发酵参数,以发酵温度、发酵时间、底物水分作为因素进行3因素4水平的正交试验,正交试验L16(43)因素水平表见表1。
1.4 马冈鹅对香蕉茎叶粉和发酵香蕉茎叶粉养分利用率的测定
选取120日龄体重为(4.00±0.50) kg的健康成年雄性马冈鹅32只,随机分成4个组(基础饲粮组、发酵香蕉茎叶粉组、未发酵香蕉茎叶粉组和空腹组),每组8个重复,每个重复1只鹅。基础饲粮为某品牌鹅全价颗粒料,购于广州市汇升粮油店,原料组成为玉米、豆粕、次粉、面粉、小麦、石粉、磷酸氢钙、维生素、矿物质、蛋氨酸、大豆油等。基础饲粮营养水平见表2。香蕉茎叶粉占试验饲粮的百分数为15%,按每只鹅饲喂量为60 g的强饲排空法测定代谢能和养分利用率。

表1 正交试验L16(43)因素水平表
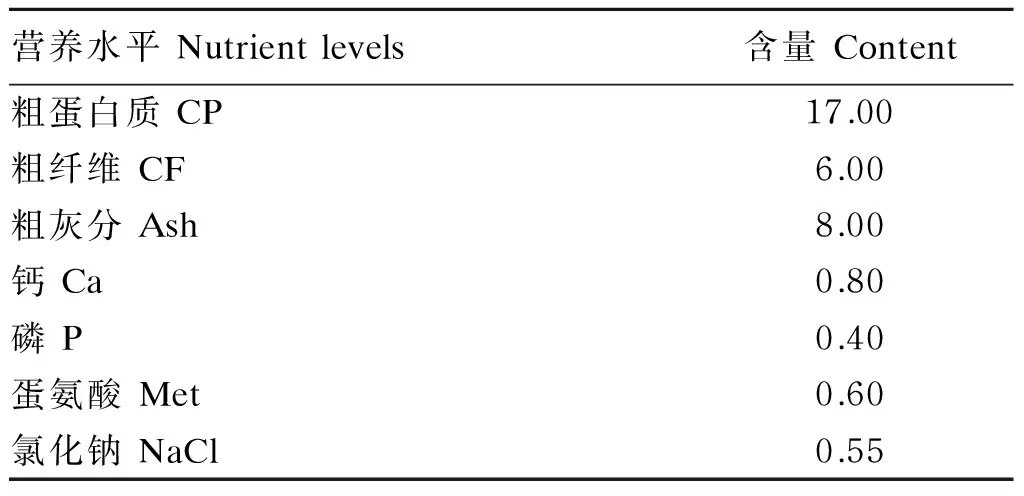
表2 基础饲粮营养水平(风干基础)
1.5 指标测定方法与计算公式
1.5.1 测定方法
粗蛋白质和真蛋白质含量用凯氏定氮法(GB/T 6432—1994)测定;粗纤维、中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)含量用范氏(Van Soest)的洗涤纤维分析法测定;总能用全自动测热仪测定;氨基酸含量使用氨基酸分析仪测定;钙含量用乙二胺四乙酸二钠络合滴定法(GB/T 6436—2002)测定;磷含量用钼酸铵分光光度法(GB/T 6437—2002)测定;粗脂肪含量用索氏抽提法(GB/T 6433—1994)测定;粗灰分含量用灼烧称重法(GB/T 6438—1992)测定。
1.5.2 代谢能和养分利用率的计算
饲粮表观代谢能(MJ/kg)=[摄入饲粮总能(MJ)-
排泄物总能(MJ)]/摄入饲粮的量(kg);
饲粮真代谢能(MJ/kg)=[摄入饲粮
总能(MJ)-排泄物总能(MJ)+内源排泄物
总能(MJ)]/摄入饲粮的量(kg);
香蕉茎叶粉表观代谢能(MJ/kg)=(试验
饲粮表观代谢能-基础饲粮表观代谢能×
0.85)/0.15;
香蕉茎叶粉真代谢能(MJ/kg)=(试验饲粮
真代谢能-基础饲粮真代谢能×0.85)/0.15;
基础饲粮养分真利用率(%)=100×[养分
总摄入量(g)-粪中养分排出量(g)+
内源养分排出量(g)]/养分总摄入量(g);
香蕉茎叶粉养分真利用率(%)=100-100×
[养分总排出量(g)-基础饲粮的养分
排出量(g)-内源养分排出量(g)]/香蕉茎叶粉的
养分摄入量(g)。
1.6 数据统计与分析
数据采用SPSS 19.0进行单因素方差分析,结合Duncan氏法进行多重比较,P<0.05表示差异显著,P<0.01表示差异极显著,结果以平均值±标准误表示。
2 结 果
2.1 单菌种发酵对发酵后香蕉茎叶粉粗蛋白质含量的影响
通过分别使用产朊假丝酵母和米曲霉对香蕉茎叶粉进行发酵,由表3可知,产朊假丝酵母发酵后的香蕉茎叶粉粗蛋白质含量显著高于空白组(P<0.05),米曲霉发酵后的香蕉茎叶粉粗蛋白质含量极显著高于空白组(P<0.01),二者均可用于做发酵菌种。
2.2 双菌种发酵
2.2.1 硫酸铵添加量对发酵后香蕉茎叶粉真蛋白质含量的影响
由表4可知,与未添加硫酸铵的发酵组相比,添加硫酸铵发酵组发酵后香蕉茎叶粉真蛋白质含量显著或极显著提高(P<0.05或P<0.01),随着硫酸铵添加量的增加,发酵后香蕉茎叶粉的真蛋白质含量呈现先增后减的趋势,在硫酸铵添加量为2%时,发酵后香蕉茎叶粉的真蛋白质含量最高,故确定硫酸铵添加量为2%。

表3 单菌种发酵对发酵后香蕉茎叶粉粗蛋白质含量的影响(绝干基础)
同列数据肩标不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01),相同或无字母表示差异不显著(P>0.05)。表2至表6同。
In the same column, values with different small letter superscripts mean significant difference (P<0.05), and with different capital letter superscripts mean significant difference (P<0.01), while with the same or no letter superscripts mean no significant difference (P>0.05). The same as Table 2 to Table 6.
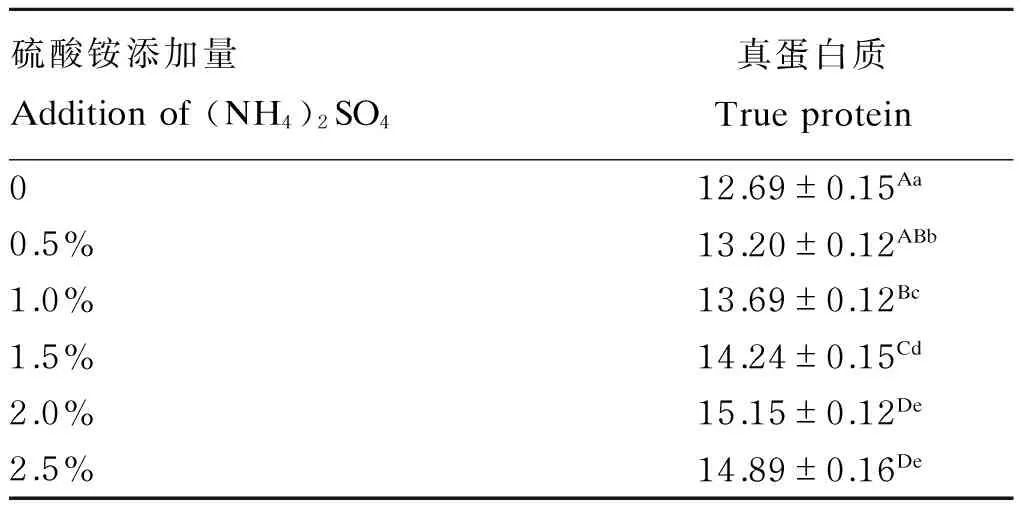
表4 硫酸铵添加量对发酵后香蕉茎叶粉真蛋白质含量的影响(绝干基础)
2种微生物的不同接种比例对发酵后香蕉茎叶粉真蛋白质含量和蛋白质净增加量都有一定的影响,由表5可知,当米曲霉和产朊假丝酵母的接种比例为2∶1时,发酵后香蕉茎叶粉的真蛋白质含量和蛋白质净增加量均为最高,故以米曲霉∶产朊假丝酵母的比例为2∶1作为2种微生物的最适接种比例。
2.2.3 混合菌液接种量对发酵后香蕉茎叶粉真蛋白质含量的影响
由表6可知,随着混合菌种接种量的增加,发酵后香蕉茎叶粉的真蛋白质含量和蛋白质净增加量呈逐渐上升的趋势,当混合菌液接种量为4%时,发酵后香蕉茎叶粉的真蛋白质含量和蛋白质净增加量最高,因此,以4%作为最适混合菌液接种量。
2.3 正交试验结果
试验按照表7的组合进行,每组6个重复,结果取平均值,结果见表7至表9。由表7正交试验直观分析中蛋白质净增加量的极差(R)值看出,发酵时间对蛋白质净增加量的影响最大,底物水分次之,发酵温度影响最小,即B>C>A。根据蛋白质净增加量的R值判断,A3B4C2为最佳的组合条件,即发酵温度30 ℃,发酵时间4 d,底物水分50%。
由表8可知,发酵温度、发酵时间、底物水分对发酵后香蕉茎叶粉蛋白质净增加量的影响不显著(P>0.05)。因素B(发酵时间)对蛋白质净增加量的影响高于因素C(底物水分)和因素A(发酵温度)。
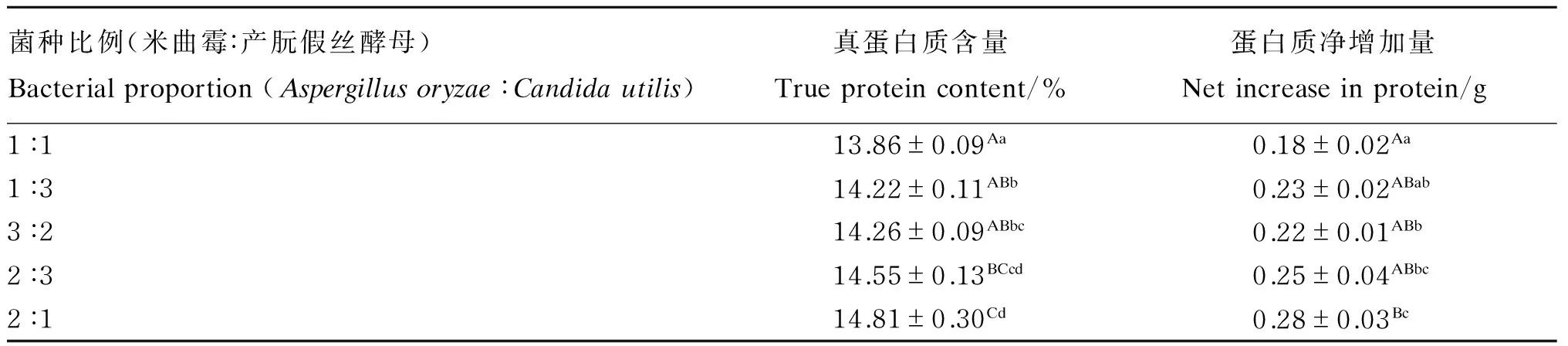
表5 菌种比例对发酵后香蕉茎叶粉真蛋白质含量的影响(绝干基础)
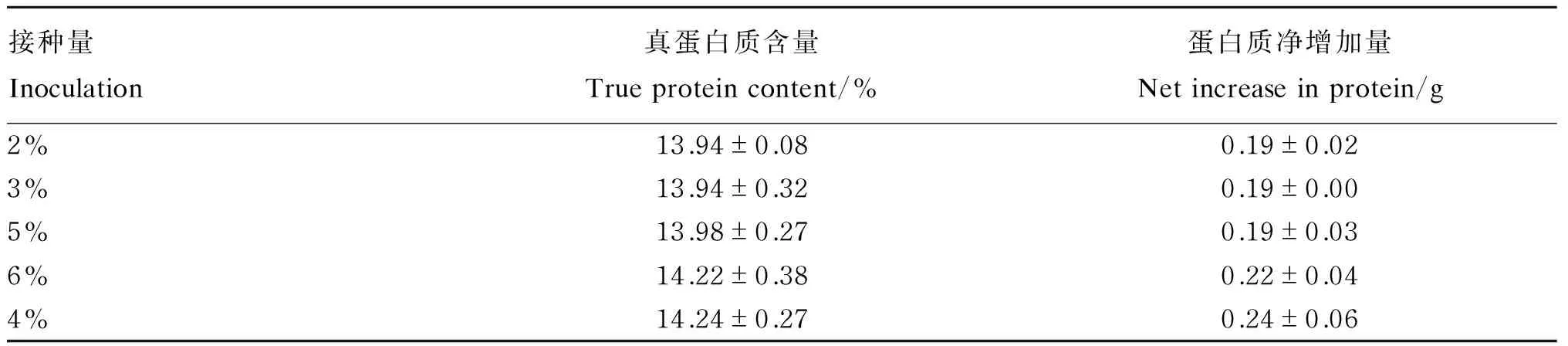
表6 混合菌液接种量对发酵后香蕉茎叶粉真蛋白质含量的影响
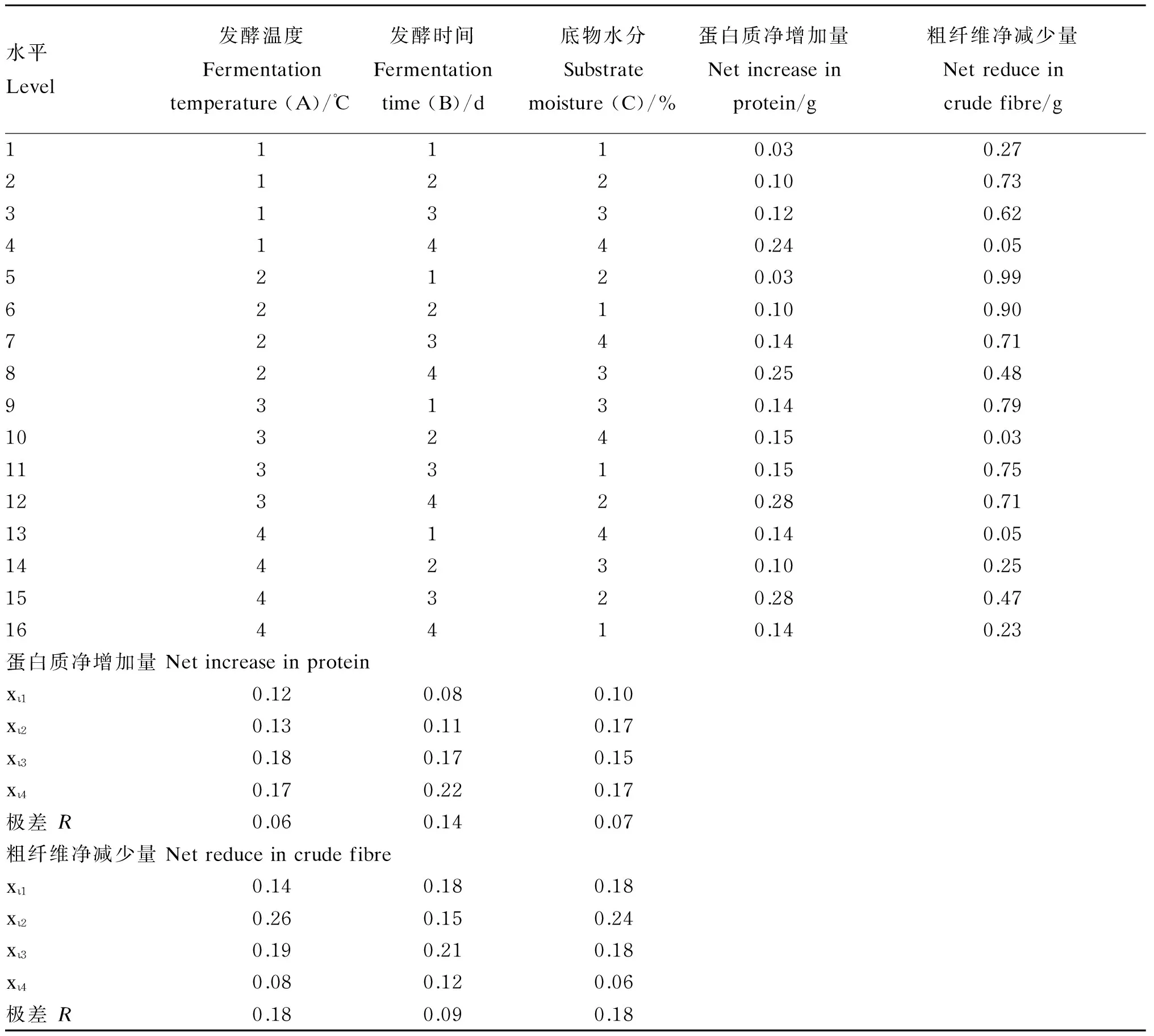
表7 正交试验结果
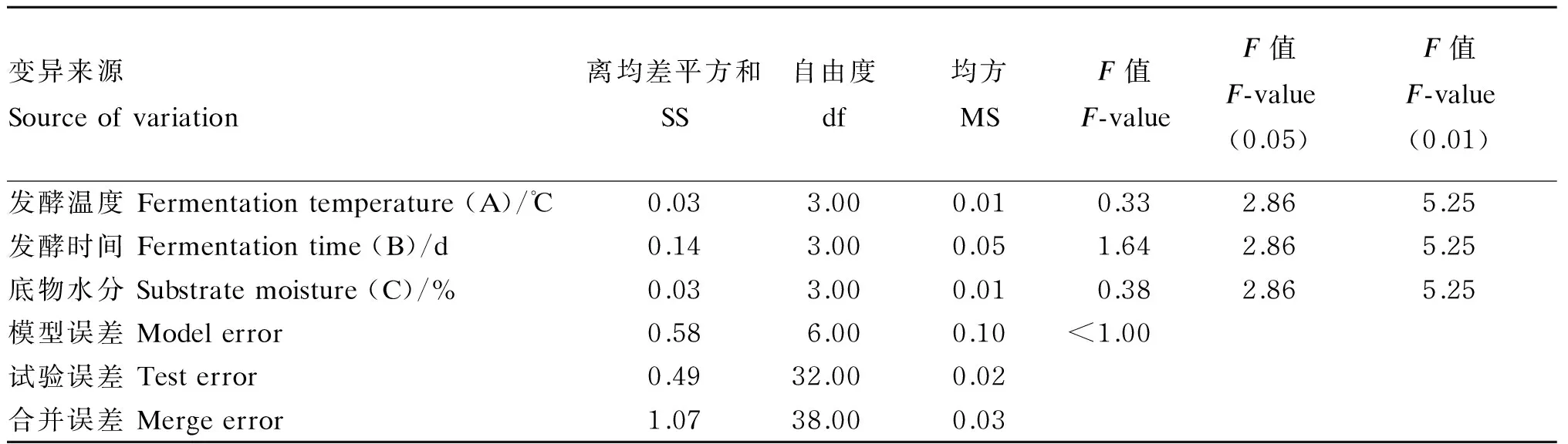
表8 正交设计试验结果方差分析(蛋白质净增加量)
由表9可知,发酵温度、发酵时间、底物水分对发酵后香蕉茎叶粉粗纤维净减少量的影响不显著(P>0.05)。因素A(发酵温度)和因素C(底物
中央空调主机智能节能系统利用云端网络建立管理平台,控制器与管理服务器双向传输。系统的控制器和数字电表等可支持RS485接口,接到主机加/卸载控制柜。系统管理人员可以通过各种网络终端设备(包括移动终端设备)即时掌握空调主机运行状况的所有信息,示意图如4所示(注:此图仅供参考与示意,实际架构以设备数量为主)。
水分)对粗纤维净减少量的影响均大于因素B(发酵时间)。
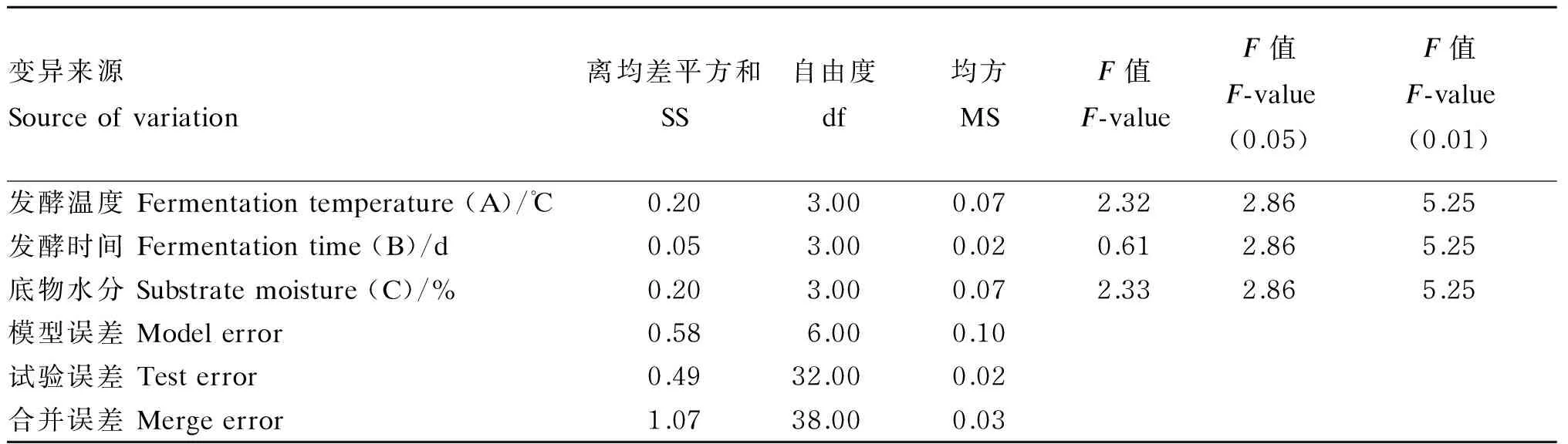
表9 正交设计试验结果方差分析(粗纤维净减少量)
2.4 香蕉茎叶粉氨基酸含量分析
依据本试验所获得的发酵工艺,即硫酸铵添加量2%,菌种接种比例2∶1(米曲霉∶产朊假丝酵母),菌液接种量4%,发酵温度30 ℃,底物水分50%,发酵时间4 d,经过该工艺发酵后的香蕉茎叶粉氨基酸含量如表10所示,发酵后香蕉茎叶粉的必需氨基酸中除了赖氨酸含量降低了13.56%,其余的氨基酸含量均有不同程度地提高,其中蛋氨酸、异亮氨酸和苏氨酸含量提高最明显,分别为40.00%、32.00%和23.21%,非必需氨基酸中半胱氨酸和组氨酸含量提高明显,分别为100.00%和78.79%。
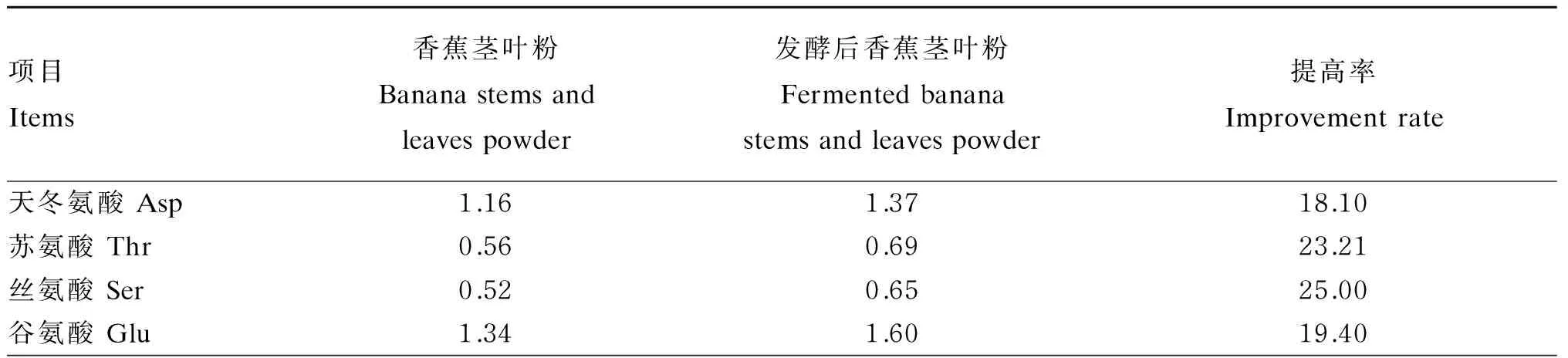
表10 发酵前后香蕉茎叶粉氨基酸含量(风干基础)
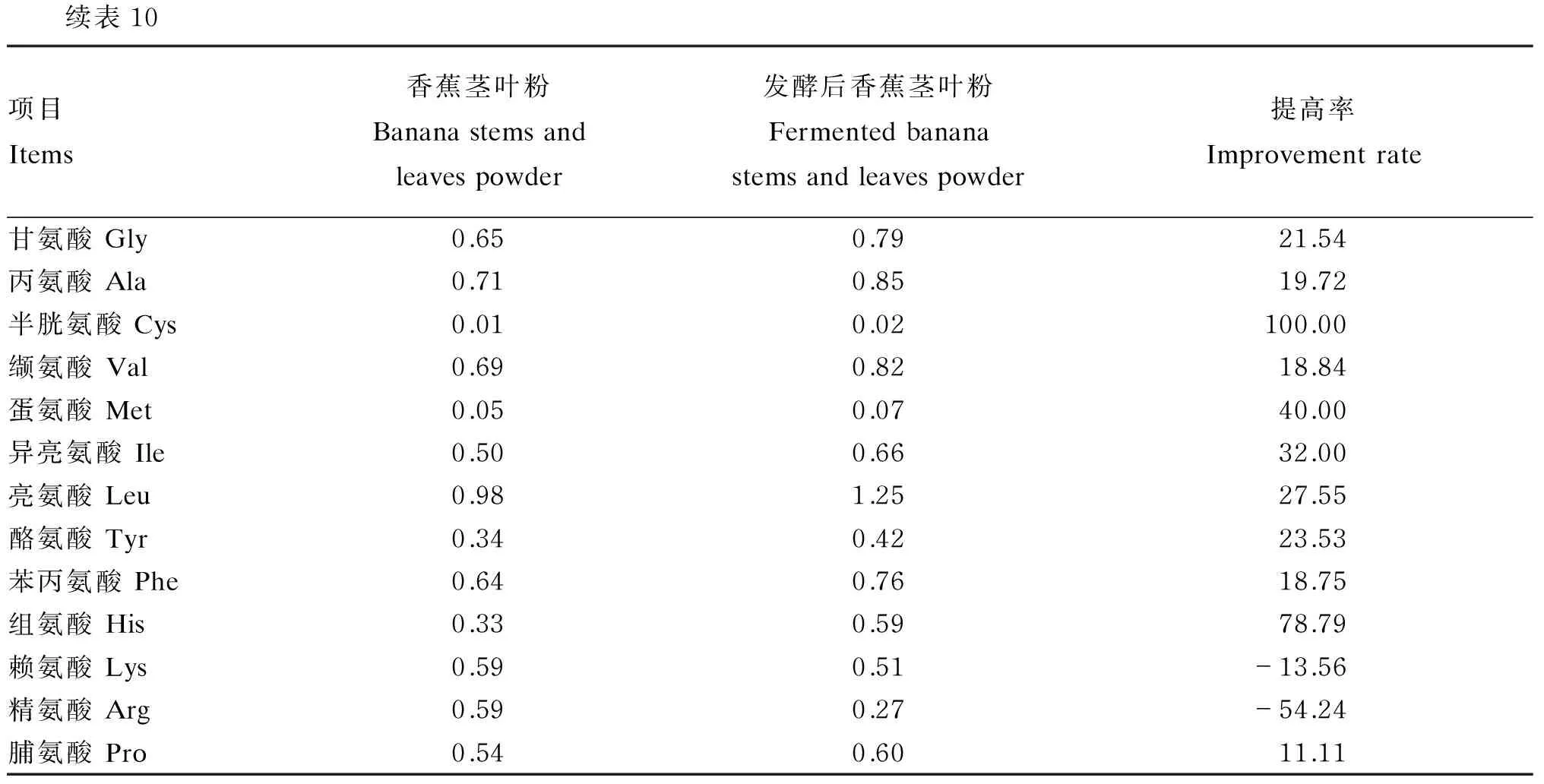
续表10项目Items香蕉茎叶粉Bananastemsandleavespowder发酵后香蕉茎叶粉Fermentedbananastemsandleavespowder提高率Improvementrate甘氨酸Gly0.650.7921.54丙氨酸Ala0.710.8519.72半胱氨酸Cys0.010.02100.00缬氨酸Val0.690.8218.84蛋氨酸Met0.050.0740.00异亮氨酸Ile0.500.6632.00亮氨酸Leu0.981.2527.55酪氨酸Tyr0.340.4223.53苯丙氨酸Phe0.640.7618.75组氨酸His0.330.5978.79赖氨酸Lys0.590.51-13.56精氨酸Arg0.590.27-54.24脯氨酸Pro0.540.6011.11
2.5 马冈鹅对香蕉茎叶粉养分利用率
发酵前后香蕉茎叶粉养分含量见表11,与发酵前相比,发酵后香蕉茎叶粉粗蛋白质含量提高了33.82%,单宁含量降低了9.38%。
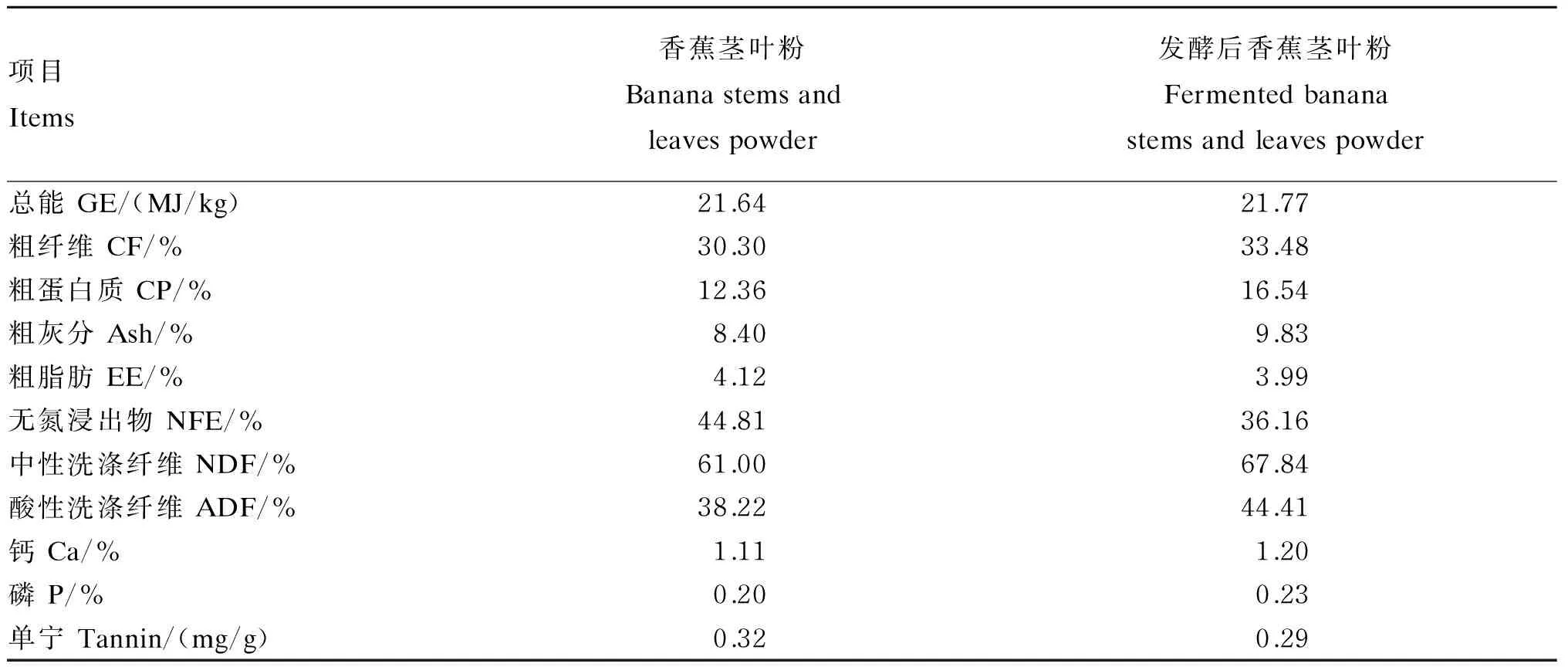
表11 香蕉茎叶粉发酵前后营养成分和单宁含量(绝干基础)
由表12可知,与发酵前相比,发酵后香蕉茎叶粉真代谢能提高了7.64%(P>0.05),粗蛋白质真利用率提高了52.66%(P<0.01)。
3 讨 论
3.1 单菌种发酵对发酵后香蕉茎叶粉粗蛋白质含量的影响
微生物的生长繁殖需要摄取基质的养分来维持,本试验的单菌种发酵结果显示,米曲霉和产朊假丝酵母都能在以香蕉茎叶粉为唯一养分来源的条件下生长繁殖,较之产朊假丝酵母,经过米曲霉发酵后的香蕉茎叶粉粗蛋白质含量更高,这可能是米曲霉能更快地消耗香蕉茎叶中的可溶性糖以及纤维素,造成粗蛋白质的含量升高。
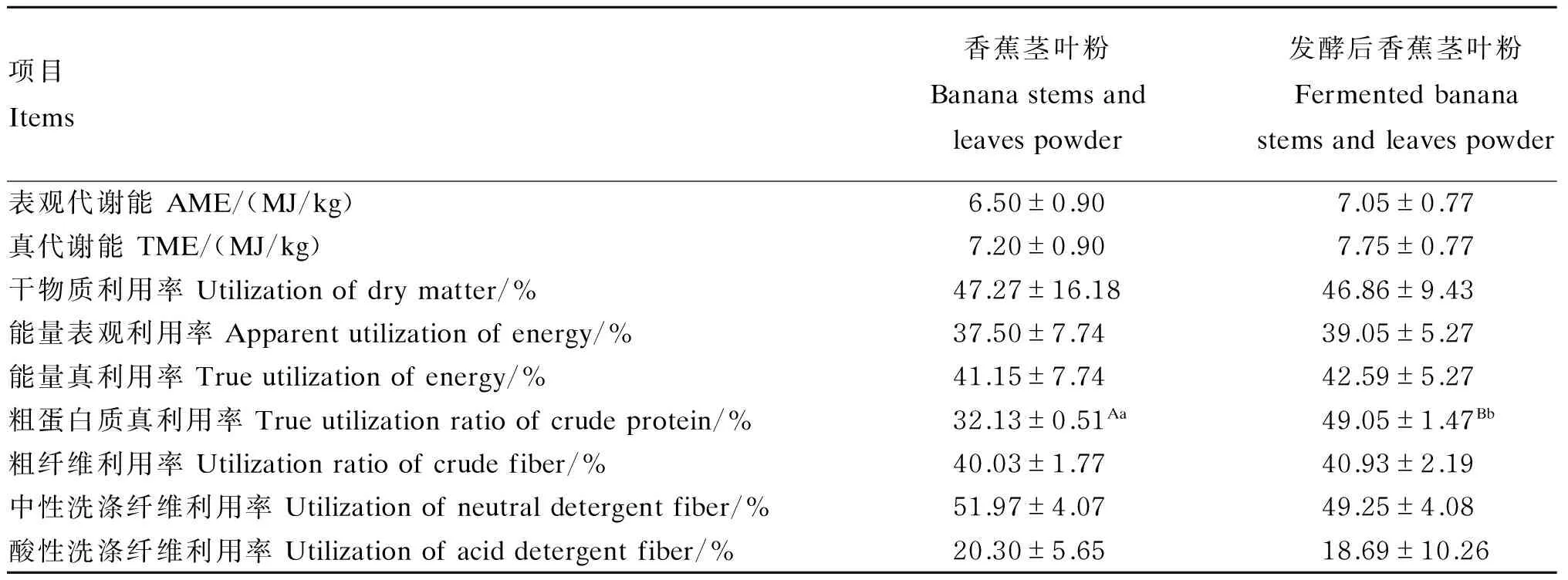
表12 香蕉茎叶粉发酵前后养分利用率
同行数据肩标不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01),相同或无字母表示差异不显著(P>0.05)。
In the same row, values with different small letter superscripts mean significant difference (P<0.05), and with different capital letter superscripts mean significant difference (P<0.01), while with the same or no letter superscripts mean no significant difference (P>0.05).
3.2 硫酸铵添加量对发酵后香蕉茎叶粉真蛋白质含量的影响

3.3 菌种比例和接种量对发酵后香蕉茎叶粉真蛋白质含量的影响
发酵过程中米曲霉和产朊假丝酵母是以一个整体的形式存在,米曲霉和产朊假丝酵母在发酵过程中存在着协作与竞争的关系,相互制约又彼此依存。酵母菌主要利用糖源,生长过程中可以产生大量的菌体蛋白,霉菌可分泌纤维素酶,降解基质中的粗纤维[9]。本试验结果显示,当2种微生物的接种比例一定时,菌种的接种量对发酵产物的真蛋白质增加量影响不显著,这与前人的研究结果[10]基本一致,说明菌种的接种量对发酵的过程影响相对较小,因每批发酵试验所制备的菌液浓度略有差异,故不同批次的发酵不做比较。
3.4 正交试验结果分析
本试验结果显示,发酵温度28 ℃时,粗纤维净减少量最多,El-Ghonemy等[11]的试验显示,米曲霉产纤维素酶的最适温度为28 ℃,这与本试验的结果相对应。一般认为固态发酵底物合适的水分在40%~70%之间[12]。发酵底物的含水量过高会导致基质间的空隙变少,降低了氧气的渗透,底物水分过低不利于底物养分的溶解,阻碍微生物生长[13]。Bhanja等[14]的试验结果发现,米曲霉固态培养基干湿比为1∶1即水分含量为50%时,米曲霉生长最旺盛,这与本试验发酵水分为50%时粗纤维净减少量最多的结果相同。发酵产物中的蛋白质含量提高依赖于2种微生物在发酵过程中利用有机氮源和无机氮源的大量繁殖,前人的研究认为,固态发酵系统适宜的发酵温度是25~35 ℃[15-16],微生物中的核酸和蛋白质对温度非常的敏感,温度通过影响微生物的酶系统从而影响微生物对养分的吸收和积累,温度过高或过低都不利于微生物的生长。前人的研究结果显示,产朊假丝酵母适宜的生长温度与水分为30 ℃和50%~125%[17-18]。本试验结果显示,发酵前3 d随着发酵天数的延长,粗纤维净减少量增加。而蛋白质净增加量随着发酵时间的延长不断增多,这说明产朊假丝酵母在米曲霉的作用达到稳定后仍然利用硫酸铵持续增殖。
3.5 发酵前后香蕉茎叶粉氨基酸含量和概略养分的变化分析
米曲霉和产朊假丝酵母合成自身蛋白质的过程将硫酸铵转化为自身需要的氨基酸,使发酵产物的氨基酸组成发生了变化。陈中平[19]的试验结果证明,米曲霉在生长过程会消耗物料中的赖氨酸;余冰等[20]的试验表明,产朊假丝酵母生长需要消耗精氨酸,这也是本试验发酵后的香蕉茎叶粉中赖氨酸和精氨酸含量较发酵前低的原因。米曲霉和产朊假丝酵母的生命活动将底物中的可溶性糖转化为水和二氧化碳,使发酵后无氮浸出物含量减少了8.65%,在发酵过程中尽管米曲霉分泌的纤维素酶分解了底物中的部分纤维素,但由于可溶性糖的消耗使粗纤维、中性洗涤纤维和酸性洗涤纤维含量反而提高。发酵后的粗蛋白质含量较发酵前提高了4.18个百分点,因为微生物利用硫酸铵合成自身蛋白质,加之可溶性糖的消耗使蛋白质得到累积,这与Hong等[21]的研究结果一致。微生物在发酵过程当中消耗了基质中部分的能量用于自身的生命活动,但发酵后的香蕉茎叶粉总能较发酵前略有提高,余冰等[20]研究表明发酵豆粕总能也比发酵前高出5.43%,说明微生物在发酵过程中将原料的养分进行了浓缩,使总能提高。
3.6 发酵前后鹅对香蕉茎叶粉养分利用率分析
发酵后香蕉茎叶粉粗蛋白质的利用率极显著高于发酵前。香蕉茎叶中的蛋白质部分与单宁结合在一起阻碍动物对蛋白质的利用,进入消化道的单宁会与肠道黏膜蛋白质结合形成不溶物并排出体外,造成体氮流失[22-23]。米曲霉具有分泌单宁酶的功能,能分解发酵物料中的单宁[24-26]。本试验结果显示,发酵后的香蕉茎叶粉单宁含量较发酵前减少了9.38%,这对于提高发酵后香蕉茎叶蛋白粉的蛋白质利用率有一定的作用。发酵过程使香蕉茎叶粉的蛋白质一部分转化为菌体自身的蛋白质,一部分被蛋白酶分解为多肽,寡肽和氨基酸成为易被动物肠道吸收的氮源,加上米曲霉和产朊假丝酵母利用硫酸铵合成菌体蛋白也成为能够被动物消化利用的蛋白质,最终使发酵后的香蕉茎叶粉的蛋白质利用率极显著的高于发酵前。发酵前的香蕉茎叶粉的粗纤维含量低于发酵后的香蕉茎叶粉,但马冈鹅对发酵后的香蕉茎叶粉中的粗纤维利用率略高于发酵前,这说明发酵后香蕉茎叶粉中的粗纤维更容易被鹅消化和利用。牛建涛等[27]利用米曲霉分解纤维的研究表明,随着米曲霉的生长纤维表面遭到破坏,部分纤维素开始断裂,纤维的内部结构受到破坏,晶体结构发生改变。发酵过程中米曲霉使香蕉茎叶粉中的粗纤维在不同程度上受到了破坏,进入鹅肠道后,在肠道微生物的作用下更容易被分解利用,使发酵后香蕉茎叶粉的粗纤维利用率提高。因发酵后香蕉茎叶粉的蛋白质和粗纤维利用率提高,加上发酵后香蕉茎叶粉的总能略高于发酵前,所以发酵后的代谢能和能量利用率也高于发酵前。
4 结 论
① 向香蕉茎叶粉中加入2%的硫酸铵,按接种量为4%,接入米曲霉孢子与产朊假丝酵母比例为2∶1的混合菌液,将水分调至50%,在30 ℃下发酵4 d后,其总能略有提高,粗蛋白质含量提高了33.82%,15种氨基酸含量有不同程度的提高。
② 马冈鹅对香蕉茎叶粉中的能量、中性洗涤纤维和粗纤维利用率较高,香蕉茎叶粉蛋白质含量不高,直接饲喂马冈鹅对其蛋白质利用率较低。
③ 马冈鹅对发酵香蕉茎叶粉的代谢能、能量利用率、粗纤维利用率较发酵前略有提高,蛋白质的利用率极显著高于发酵前,说明经过发酵后的香蕉茎叶粉养分得到改善,有益于鹅对香蕉茎叶粉养分的利用。
[1] 王增煌,王文策,杨琳.香蕉茎叶作为饲料原料的研究进展[J].中国畜牧杂志,2016,52(17):82-86.
[2] 李日强,张峰,张伟峰.氨化和固态发酵玉米秸秆生产饲料蛋白的研究[J].农业环境科学学报,2006,25(6):1636-1639.
[3] 莫重文.混合菌发酵豆渣生产蛋白质饲料的研究[J].中国饲料,2007(14):36-38.
[4] ROSMA A,CHEONG M W.Effects of nitrogen supplementation on yeast (Candidautilis) biomass production by using pineapple (Ananascomosus) waste extracted medium[J].Malaysian Journal of Microbiology,2007,3(1):19-26.
[5] 刘延琳,刘伟,夏加贵.白葡萄酒活性干酵母对不同氮源利用的研究[J].微生物学杂志,2007,27(1):88-90.
[6] 刘明启,刘光富,戴贤君,等.优化米曲霉固体发酵产果胶酶及产物酶学性质[J].中国计量学院学报,2010,21(2):64-69,178.
[7] 李平兰.食品微生物学教程[M].北京:中国林业出版社,2011.
[8] 籍保平,尤希风,张博润.苹果渣发酵生产饲料蛋白的培养基[J].中国农业大学学报,1999,4(6):53-56.
[9] 孙展英.马铃薯渣固态发酵工艺研究及生物学价值评定[D].硕士学位论文.保定:河北农业大学,2014.
[10] 欧荣娣,范觉鑫,王升平,等.红薯渣固态发酵条件优化[J].动物营养学报,2015,27(10):3302-3310.
[11] EL-GHONEMY D H,ALI T H,EL-BONDKLY A M,et al.Improvement ofAspergillusoryzaeNRRL 3484 by mutagenesis and optimization of culture conditions in solid-state fermentation for the hyper-production of extracellular cellulase[J].Antonie van Leeuwenhoek,2014,106(5):853-864.
[12] DOELLE H W,MITCHELL D A,ROLZ C E.Solid substrate cultivation[M].New York:Springer,1992.
[13] VU V H,PHAM T A,KIM K.Improvement of fungal strain by repeated and sequential mutagenesis and optimization of solid state fermentation for the hyper-production of raw-starch-digesting enzyme[J].Journal of Microbiology and Biotechnology,2010,20(4):718-726.
[14] BHANJA T,ROUT S,BANERJEE R,et al.Comparative profiles of α-amylase production in conventional tray reactor and GROWTEK bioreactor[J].Bioprocess and Biosystems Engineering,2007,30(5):369-376.
[15] MAURYA D P,SINGH D,PRATAP D,et al.Optimization of solid state fermentation conditions for the production of cellulase byTrichodermareesei[J].Journal of Environmental Biology,2012,33(1):5-8.
[16] KIM D M,CHO E J,KIM J W,et al.Production of cellulases byPenicilliumsp.in a solid-state fermentation of oil palm empty fruit bunch[J].African Journal of Biotechnology,2014,13(1):145-155.
[17] 张冬艳,张瑞兰,张通.绿色木酶与产朊假丝酵母混合发酵生产纤维素酶和单细胞蛋白[J].内蒙古工业大学学报,2003,22(3):180-185.
[18] 吴远根,彭湘屏,张晓娟,等.产朊假丝酵母固态发酵麻疯树饼粕产菌体蛋白的研究[J].食品工业科技,2009(2):161-164.
[19] 陈中平.米曲霉发酵对豆粕营养特性的影响[D].硕士学位论文.雅安:四川农业大学,2010.
[20] 余冰,傅娅梅,叶楠,等.固态发酵对复合蛋白质饲料营养价值改善效果的研究[J].动物营养学报,2009,21(4):546-553.
[21] HONG K J,LEE C H,KIM S W.AspergillusoryzaeGB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals[J].Journal of Medicinal Food,2004,7(4):430-435.
[22] AHMED A E,SMITHARD R,ELLIS M.Activities of enzymes of the pancreas,and the lumen and mucosa of the small intestine in growing broiler cockerels fed on tannin-containing diets[J].British Journal of Nutrition,1991,65(2):189-197.
[23] 张子仪.中国饲料学[M].北京:中国农业出版社,2000.
[24] ABDEL-NABY M A,EL-TANASH A B,SHERIEF A D A.Structural characterization,catalytic,kinetic and thermodynamic properties ofAspergillusoryzaetannase[J].International Journal of Biological Macromolecules,2016,92:803-811.
[25] PARANTHAMAN R,VIDYALAKSHMI R,SINGARAVADIVEL K.Comparative study on the suitability of different substrates for tannin acyl hydrolase production usingAspergillusoryzae[J].Journal of Pharmaceutical Sciences & Research,2009,1(4):36-42.
[26] PILLAI A R,DEVEGOWDA G,ANANDAKUMAR K.Influence of tannase and driedAspergillusoryzaeculture on performance of broilers fed tannin rich diets[J].Indian Journal of Animal Nutrition,1996,13(4):209-213.
[27] 牛建涛,张小英,官伟波.纤维素纤维织物在米曲霉作用下的生物降解[J].上海纺织科技,2013,41(5):1-3,19.
*Corresponding author, professor, E-mail: ylin@scau.edu.cn
(责任编辑 武海龙)
Study on Optimal Solid State Fermentation Conditions of Banana Stems and Leaves Powder and Its Nutrient Utilization for Geese
WANG Zenghuang WANG Wence ZHAI Shuangshuang XIE Qiang ZUO Xin YANG Lin*
(CollegeofAnimalScience,SouthChinaAgriculturalUniversity,Guangzhou510642,China)
This study was conducted to use the solid state fermentation technology to improve nutrient composition of banana stems and leaves and its nutrient utilization for geese. ChoosingAspergillusoryzaeand Candida utilis as fermentation strains to investigate the influences of 5 factors [single strain fermentation, addition of (NH4)2SO4, proportion ofAspergillusoryzaeandCandidautilis, mixed bacteria liquid inoculation] on the protein content of banana stems and leaves powder after fermentation. Using orthogonal design to find out the optimal combination of fermentation temperature, fermentation time and substrate moisture. The results showed that usingAspergillusoryzaeorCandidautilisas single fermentation strain could significantly increase the crude protein content of fermented banana stems and leaves powder (P<0.05 orP<0.01). Fermentation with (NH4)2SO4addition could significantly increase true protein content of fermented banana stems and leaves powder (P<0.05 orP<0.01), especially 2% addition. The net increase in protein treated with the proportion ofAspergillusoryzaeandCandidautilisby 2∶1 significantly higher than that of the proportion of 1∶1, 1∶3 and 3∶2 (P<0.05 orP<0.01). The results of orthogonal test showed that the best fermentation condition were 4% inoculation (the proportion ofAspergillusoryzaeandCandidautilisby 2∶1) and 2% (NH4)2SO4addition, and fermenting in 50% substrate moisture, 30 ℃ for 4 days. The crude protein content of banana stems and leaves increased by 33.82% after this fermentation process. The amino acid analysis showed that there were 15 kinds of amino acids increased differently beside lysine and arginine. The metabolic test results ofMaganggeese showed that the crude protein utilization of banana stems and leaves powder after fermentation significantly increased by 52.66% comparing with the banana stems and leaves powder without fermentation (P<0.01), additionally, the energy and metabolic energy utilization were increased slightly as well. In conclusion, the nutritional value of banana stems and leaves powder is improved after this fermentation process, and also promote the digestion and absorption of nutrient of geese.[ChineseJournalofAnimalNutrition, 2017, 29(4):1283-1293]
banana stems and leaves powder; solid state fermentation; protein; nutrient utilization rate;Maganggeese;Aspergillusoryzae;Candidautilis
10.3969/j.issn.1006-267x.2017.04.025
2016-10-09
公益性行业科研专项(201303143)
王增煌(1990—),男,福建松溪人,硕士研究生,从事非常规饲料资源研究。E-mail: 397421559@qq.com
*通信作者:杨 琳,教授,博士生导师,E-mail: ylin@scau.edu.cn
S816.34
A
1006-267X(2017)04-1283-11
