埃氏小球藻对11种抗生素的敏感性
2017-04-14王亚君周广航季春丽薛金爱李润植
王亚君,周广航,季春丽,薛金爱,李润植
(山西农业大学分子农业与生物能源研究所,山西太谷030801)
埃氏小球藻对11种抗生素的敏感性
王亚君,周广航,季春丽,薛金爱,李润植
(山西农业大学分子农业与生物能源研究所,山西太谷030801)
为了探讨埃氏小球藻(Chlorella emersonii)株系SXND-12对11种抗生素的敏感性,采用分光光度法研究了氯霉素、潮霉素、红霉素、利福平霉素、卡那霉素、硫酸新霉素、头孢霉素、大庆霉素、氨苄霉素、羧卞霉素、链霉素等11种抗生素对埃氏小球藻生长的影响,从而来确定分离、纯化藻株时对藻细胞无害并能抑制伴生杂菌生长的抗生素浓度。结果表明,埃氏小球藻对头孢霉素、氨苄霉素、羧卞霉素、硫酸新霉素敏感性较弱,对利福平霉素较为敏感,200 μg/mL(致死剂量)即0.243 mmol/mL(致死浓度)时,在固体培养中抑制了埃氏小球藻的生长;对潮霉素、氯霉素、红霉素、链霉素、大庆霉素、卡那霉素非常敏感,在固体培养中的致死剂量分别达到20 μg/mL(0.101 mmol/mL),50 μg/mL(0.155 mmol/mL),30 μg/mL(0.041 mmol/mL),50 μg/mL(0.086 mmol/mL),50 μg/mL(0.105 mmol/mL),100 μg/mL(0.206 mmol/mL)。研究可为筛选出小球藻基因工程藻株的筛选试剂、诱变筛选的初筛试剂、建立无菌体系的选择试剂奠定基础。
埃氏小球藻;抗生素;敏感性;生长抑制
随着经济的不断发展和人口的不断增加,全球范围内的能源枯竭现象越来越严重,传统石化资源的开发和利用面临着前所未有的严峻形势:石油、天然气在50 a内将消耗殆尽,煤炭在100 a后也将枯竭[1],因此,发展新型的可再生生物能源——生物柴油已刻不容缓。生物柴油是一种可替代石化柴油的新型能源,而以动植物油脂、废弃油脂来制备生物柴油[2],原料成本占到70%以上,并且原料供给不稳定,产品质量无法保证。能源微藻是近年来发展起来的一种新型的产油材料,在缺氮的条件下油脂可大量积累,并且主要化学成分非常适合制备生物柴油,其被认为具有广阔的发展前景[3]。
微藻是一种光合自养型生物,同等条件下光合速率高于其他植物数倍,不仅具有较短的生长周期,而且在适应性、生物量等方面具有巨大的优势。小球藻可大规模培养[4],油脂含量又高,使其具备了制备一种新型油料物质的潜力[5]。但是目前自然界存在的藻种都无法满足大规模工业生产的需求。所以,当前获得优异工业藻株是微藻规模化培养和联产微藻燃油的一个关键制约因素。培育优异藻种通常有遗传转化和诱变2种主要方法。其中,遗传转化目标针对性强,诱变产生的突变株较多,短时间内就可能产生一个巨大的突变库。但是如何高效地筛选目标变异藻株是这2种方法面临的一个重要问题。在遗传转化体系中,外源基因成功导入受体细胞后,会使新的菌株带有某种特殊的性质,即抗生素阳性选择性标记,可为大规模的藻种鉴定提供强有力的帮助。而对于诱变产生的不定向突变体的初筛,工作量十分庞大。选取合适的抗生素[6]作为初筛试剂,可大大减少筛选工作量,提高筛选效率。
为筛选得到可用作埃氏小球藻基因工程和诱变产生突变体的选择试剂,本试验选取了常见的11种不同抗生素,研究埃氏小球藻对它们的敏感性,旨在为建立和完善埃氏小球藻突变体筛选体系提供参考和依据。
1 材料和方法
1.1 藻种
试验所用的埃氏小球藻(Chlorella emersonii)株系SXND-12由山西农业大学分子农业与生物能源研究所从山西省境内分离、鉴定获得,现保存于山西省科技重大专项微藻燃油基地实验室。
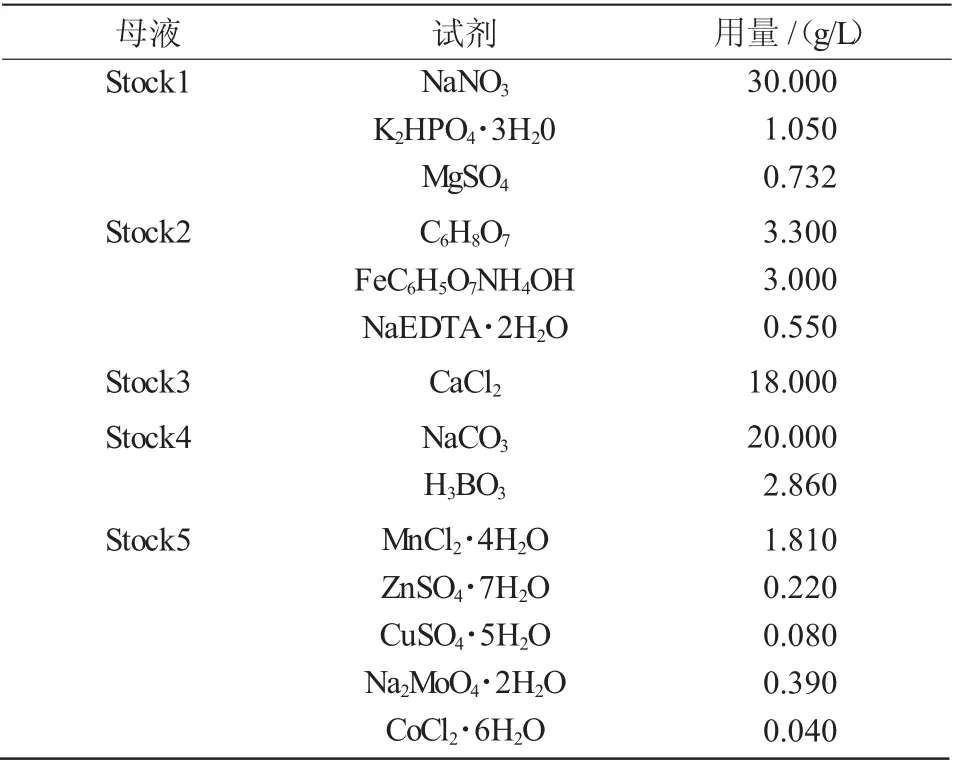
表1 BG11培养基(母液)配方
本试验主要以埃氏小球藻在含有不同种类和不同浓度的抗生素固体培养基上藻落长出的时间、一定时间内藻落存活的数量以及整个生长过程中的生长速度及状态,作为抗生素影响埃氏小球藻生长的指标,并计算出各条件下的致死率,进而筛选出适合遗传转化和化学物理诱变初筛的抗生素试剂。
1.2 培养基
BG11液体和固体培养基按照表1,2的配方进行配制,121℃高压灭菌20 min。灭菌前固体培养基中添加1.2%的琼脂。按照实验要求加入各种抗生素。

表2 BG11工作液配方
1.3 主要试剂
链霉素(Streptomycin)、氯霉素(Chlorampheni col)购自华美生物工程公司,卡那霉素(Kanamycin)、氨苄霉素(Ampicillin)、羧苄霉素(Carbenicillin)、潮霉素(Hvgromvcin)购自广州博衍生物科技有限公司,头孢霉素(Cefotaxime)、庆大霉素(Gentamycin)购自郑州豫新制药公司,利福平霉素(Rifampicin)、红霉素(Erythromycin)、硫酸新霉素(Neomycin)购自Invitrogen公司。其中,潮霉素购买的是100 mg/mL母液,其他的均是粉末,母液的配制参照文献[7]进行,并用0.22 μm/mL滤膜过滤除菌。
1.4 培养方法
1.4.1 接种物的制备 在平板上挑取体积较大、生长状态良好的埃氏小球藻藻落,利用接种针将其接种于含有一定量新鲜培养液的三角烧瓶中,7 d后转接到体积为250 mL的三角烧瓶进行扩大培养,长至指数期离心,转速为6 000 r/min,时间5 min,倒掉上清液,无菌水洗涤2~3次,用一定量新鲜的灭菌培养液悬浮,用于接种。
1.4.2 液体培养 将悬浮液转接到含有一定量培养液的100 mL三角烧瓶中,利用纱布将瓶口包裹若干层,防止细菌侵入。设置培养条件为光照强度5 000 lx,湿度控制在50%,温度控制在(25± 0.5)℃,光暗比12 h∶12 h,pH值8.5。
1.4.3 固体培养 取一定量对数期的藻液,取100μL涂布于BG11固体平板上,设置培养条件为光照强度5 000 lx,湿度控制在50%,温度控制在(25± 0.5)℃。
1.5 埃氏小球藻对11种抗生素敏感性测试
将埃氏小球藻进行固体培养并按抗生素的种类分为11组试验,每组6~8个梯度,每个梯度做3个平行,其中,将抗生素浓度为0的试验组作为对照,以消除试验误差。根据预试验结果,各种抗生素的最终浓度设置列于表3。每天观察各平板上藻落生长情况,30 d时统计长出的藻落数目,计算致死率。

式中,X0表示对照组平板上长出的藻落数,X1表示各种抗生素平板上长出的藻落数目。

表3 11种抗生素的最终质量浓度梯度 μg/mL
2 结果与分析
2.1 埃氏小球藻对氨苄霉素、羧苄霉素、硫酸新霉素和头孢霉素敏感性固体试验结果
在固体培养中,在设定的试验浓度范围内,Amp(氨苄霉素),Car(羧苄霉素),NM(硫酸新霉素),Cefo(头孢霉素)等4种抗生素基本不影响小球藻的正常生长。光照培养箱内适宜条件下培养7 d左右,对照组和试验组均能长出大量的藻落,10 d左右时,平板上的藻落数目趋于稳定。图1~图4是各试验组在相同条件下培养10 d时的图片。统计发现,图1~图4每个平板上的藻落数分别大约为3 500,1 500,1 800,2 200个,每个试验组内各个浓度平板上的藻落数量基本一致,误差在±5%左右。因此,从藻落长出的时间和数目这2个方面进行判断可知,在试验范围内这4种抗生素对小球藻的生长没有影响。



2.2 埃氏小球藻对氯霉素敏感性固体试验结果
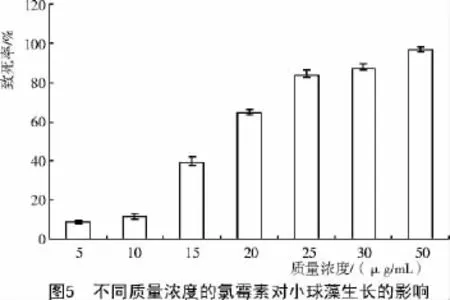
氯霉素通过阻碍蛋白质合成,抑制细胞生长,导致细胞死亡,对很多藻类的生长均有一定的抑制作用[8-12]。黄健等[8]研究表明,新月菱形藻(Nitzschia closterium Ehr.)、牟勒氏角刺藻(Chaetoceros muelleri Lemm.)、三角褐指藻(Phaeodactylum tricornutum Bohl.)、绿光等鞭金藻8701(Isochrysis galbana Parke 8701)、亚心形扁藻(Platymonas subcordiformis(Wille)Hazen)、小球藻(Chlorella vulgaris Beij.)等6种海洋微藻对氯霉素普遍敏感。曹军平等[10]对金藻的研究也得出了相似的结论。本试验的结果也证实了这一点。由图5可知,质量浓度在5~10 μg/mL范围内,氯霉素对小球藻的生长起到了一定的延缓作用,因为与对照组相比藻落在固体培养基上长出的时间延迟了5 d。当氯霉素达到50μg/mL时,小球藻的致死率达到96.23%,与对照相比,在30 d的培养时间内仅长出了少许藻落,且生长速率极慢,藻落也较小。本研究表明,小球藻对氯霉素的致死质量浓度为50 μg/mL(0.155 mmol/mL),与其他研究结果[7]较为吻合。但是用氯霉素作为遗传转化的选择性标记时需要注意,氯霉素的溶剂是乙醇,相关研究表明,乙醇对某些藻的生长具有一定的影响,所以,选择氯霉素时要考虑乙醇对试验的影响。
2.3 埃氏小球藻对红霉素敏感性固体试验结果
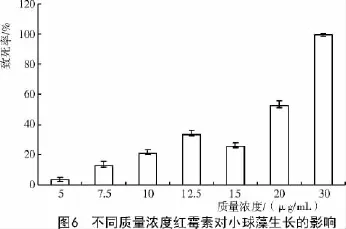
红霉素主要通过抑制mRNA的移位阻碍转肽作用,使核蛋白体上延伸的肽链解离,不再形成正常功能的蛋白质,从而抑制蛋白质的生物合成。刘滨扬等[11]关于红霉素对微藻敏感性的研究证实,红霉素对某些微藻在很低的浓度时就会有较强的抑制作用。小球藻对红霉素非常敏感,在很低的浓度下就完全抑制了小球藻的生长,它的抑制能力甚至超过了报道中常提到的潮霉素和氯霉素[12]。从图6可以看出,在5~20 μg/mL范围内,随着抗生素质量浓度的不断增加,小球藻对其敏感性也呈增强趋势,当红霉素质量浓度高于20 μg/mL时,小球藻的致死率急剧增加,从20 μg/mL的51.75%骤增至30 μg/mL的100%。
红霉素自发现以来广泛的应用于临床医学,在小球藻性基因方面应用的比较少,但鉴于小球藻对其敏感性,红霉素具有作为阳性筛选标记基因的潜在价值。
2.4 埃氏小球藻对利福平霉素敏感性固体试验结果
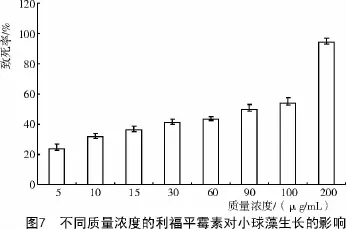
利福平霉素是通过阻碍DNA正常转录过程而达到杀菌目的的一种抗生素,但在微藻基因工程方面关于利福平作为抗生素的报告非常少,本试验的研究较好地填补了这个空缺。由图7可知,质量浓度为5~100 μg/mL时,小球藻的生长只是在一定程度上受到影响;质量浓度为200 μg/mL时,致死率达到了94.1%,主要表现为经过30 d的固体培养平板上长出少数几个藻落,且其生长速率极慢,可能是由抗生素破坏了小球藻的生理结构所致。另外,在研究中还发现,与其他抗生素相比,利福平霉素会使小球藻的生长产生严重的滞后现象,在整个试验浓度范围内这种现象都非常明显。当利福平霉素的质量浓度为5 μg/mL时,致死率仅为23.66%,但是它与对照相比藻落长出的时间却推迟了7 d左右;在0~100 μg/mL范围内,30 d时统计发现,在数目上各试验组基本没有差别,但是藻落长出的时间却随着抗生素浓度的增加不断地延迟,有的甚至20 d时才长出藻落;当质量浓度达到200 μg/mL时,统计发现,不仅最终存活藻落数大量减少,其生长的速度也明显迟缓。
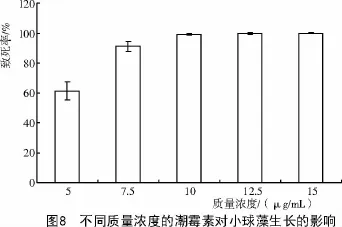
2.5 埃氏小球藻对潮霉素敏感性固体试验结果
潮霉素是小球藻基因工程中常用的抗生素[13],主要是通过磷酸转移酶的磷酸化作用,改变分子结构,致使细菌死亡[14]。王长海等[14]研究表明,潮霉素对海水小球藻的抑制性较低,但是淡化培养时5 μg/mL的质量浓度几天后就会造成小球藻死亡。另外,大量研究表明[15],海带、节旋藻、螺旋藻[16]、小球藻、裙带、盐藻等均对潮霉素有较高的敏感性。从图8可以看出,质量浓度为5 μg/mL时,小球藻的致死率达到了61.59%,与相关研究的结果基本一致;质量浓度为7.5 μg/mL时小球藻死亡率高达94.92%,严重抑制了小球藻的生长;质量浓度为15 μg/mL时,小球藻全部死亡,主要表现在在规定的培养时间内(30 d)最终存活的藻落数为0。表明埃氏小球藻对潮霉素非常敏感。
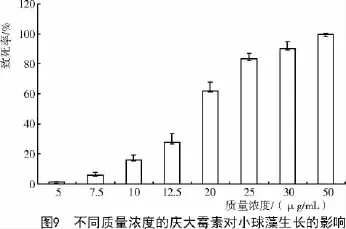
2.6 埃氏小球藻对庆大霉素敏感性固体试验结果
庆大霉素与氯霉素的抑菌原理相近,都是通过抑制蛋白质的合成起到杀菌的作用。臧晓南等[15]研究表明,Arthrospira341对庆大霉素比较敏感,在较低的浓度下,培养7 d左右就会发现有大量的藻丝死亡。王逸云[17]研究表明,海水小球藻对庆大霉素敏感性极低,几乎不影响小球藻的生长。由图9可知,当庆大霉素质量浓度为20 μg/mL时就已对小球藻的生长造成重大影响,经统计发现,在培养30 d后的固体培养基中,与对照相比藻落的数目减少约60%,个体较小、颜色呈淡黄色;当庆大霉素质量浓度达到30 μg/mL时,对小球藻的致死率达到91.04%;在50 μg/mL的固体培养基上,培养20 d仅仅有少许几个藻落长出,并且这几个藻落颜色呈土黄色,藻落如同针尖,生长非常缓慢。表明埃氏小球藻对庆大霉素比较敏感。
2.7 埃氏小球藻对卡那霉素敏感性固体试验结果
目前,卡那霉素抗性基因在遗传转化体系中应用非常广泛,是一种常见的选择性标记基因[18-20],主要是通过干扰蛋白质的合成抑制叶绿体的正常功能来杀死植物细胞。雒淑珍等[21]用卡那霉素作为选择性标记获得了具有良好性状的野罂粟转基因植株。臧晓南等[15]研究卡那霉素对Spiulina351生长影响时发现,50 μg/mL即可对其生长造成明显的影响,主要表现为微藻生长缓慢,藻体颜色变黄下沉。王长海等[14]以海水小球藻为材料进行研究,发现在卡那霉素高达500 μg/mL时,长时间培养小球藻时其依然表现出良好的长势。由图10可知,淡水小球藻对卡那非常敏感,在50 μg/mL的固体平板上培养30 d长出的藻落数目约占对照数目的0.1%,这与相关报道有较大的差别,可能是由海水与淡水离子浓度种类的差异性所造成的,离子的不同造成了抗生素表达活性的差异。

2.8 埃氏小球藻对链霉素敏感性固体试验结果
Str是链霉素的标记性基因,它在植物的遗传转化中应用比较广泛,其主要作用原理是通过无功能蛋白质的合成或失去合成蛋白的功能,致使细胞膜破裂,内溶物流出,细胞大量死亡。黄健等[8]研究表明,在其选取的5种抗生素中,亚心形扁藻对链霉素最不敏感,随抗生素浓度的提高其相对增长率变化不明显。耿德贵等[9]关于微藻对抗生素敏感性的研究表明,盐藻在较高的链霉素浓度下经过超过20 d的培养仍能健康地生长,并表现出良好的生长态势。王长海等[14]研究表明,小球藻不但对链霉素不敏感,而且在一定的浓度范围内还会起到一定的促进作用。从图11可以看出,小球藻对链霉素比较敏感,链霉素对小球藻的生长影响较大,30 μg/mL时小球藻的致死率达到了93.45%,基本上抑制了小球藻的生长;在50 μg/mL的质量浓度下培养30 d,平板上长出的藻落数目不到对照组的0.02%,说明其严重抑制了埃氏小球藻的生长。这与其他相关报道的结果相差较大,可能是这株小球藻在培养过程中,由于某些特殊的原因其生理性状发生了改变,或者是因为抗生素在一定的离子浓度下才起作用,本试验培养基所用水属于硬水,钙离子含量很高,可能影响了抗生素的某些特性,使小球藻对链霉素敏感性增加。

2.9 7种抗生素致死剂量的确定
在微藻的正常培养中,小球藻细胞壁上极易吸附细菌,这些细菌与小球藻形成共生体系,不易除去[22]。杂菌的大量繁殖会造成大量营养物质的消耗,进而影响微藻的生长。本试验通过测试微藻培养体系中埃氏小球藻藻体对抗生素的敏感性发现,氨苄、羧卞、头孢、硫酸新霉素这4种抗生素在试验浓度范围内对埃氏小球藻的生长没有影响,而细菌对抗生素却比较敏感,因此,可以根据埃氏小球藻藻体和细菌对抗生素敏感性的差异,选取这些抗生素清除微藻培养过程中杂菌,以求建立埃氏小球藻无菌培养体系,并对该体系中埃氏小球藻生长环境条件进行优化。
虽然小球藻已经可以大规模地培养,但是要真正达到工业要求的标准还需要进一步进行藻种的改良。改良藻种常用的方法是基因工程和诱变,这2种方法都存在目标藻株筛选困难的情况,前人通过研究发现,抗生素在突变株的初筛和目标藻株的筛选方面具有良好的效果。本试验研究了对埃氏小球藻比较敏感的7种抗生素(表4)以便在埃氏小球藻的育种筛选工作中更好地选择所需要的抗生素,从而为后续的试验提供参考。
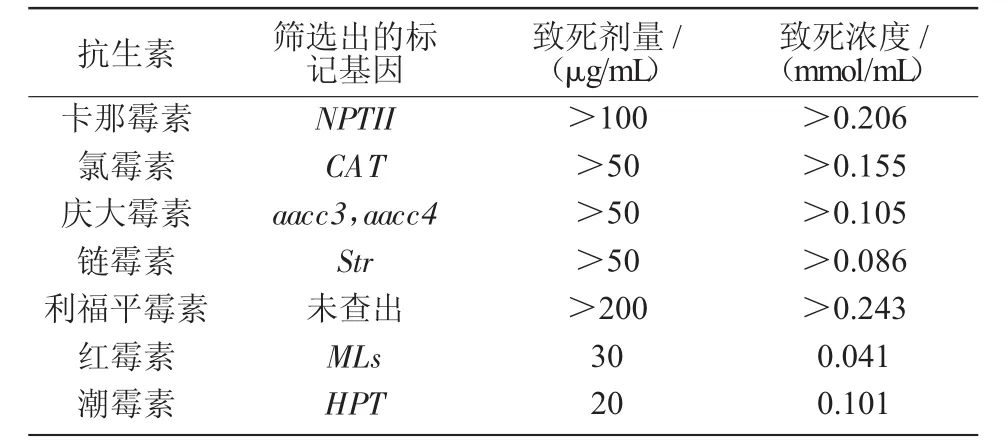
表4 7种抗生素致死浓度分析
3 结论与讨论
抗生素(antibiotics)是由微生物(包括细菌、真菌、放线菌属)或高等动植物在生活过程中所产生的具有抗病原体或其他活性的一类次级代谢产物,能干扰其他生活细胞发育功能的化学物质。
在遗传转化体系中,外源基因成功导入受体细胞后,会使新的菌株带有某种特殊的性质,即抗生素阳性选择性标记,可为大规模的藻种鉴定提供强有力的帮助。建立一个完整的转化体系,首先需要选择一个合适的阳性筛选标记,它可以在后续的转基因藻株的筛选工作中起到非常重要的作用,使筛选更加快速、高效。本试验的研究发现,埃氏小球藻对氯霉素、红霉素、卡那霉素、链霉素、庆大霉素、潮霉素等6种抗生素非常的敏感,并且这6种抗生素的标记基因已经被广泛研究和应用;潮霉素、氯霉素、卡那霉素在高等植物和藻类的遗传转化体系中已广泛使用,并取得了一些良好的效果;链霉素、红霉素在基因工程中应用较少,特别是红霉素相关的报道几乎没有,本试验研究结果可为遗传转化的阳性筛选标记提供技术参考。
诱变是改良藻种的另一种常用的方法,因为其在短时间内就可能产生一个巨大的突变库,但是诱变产生的不定向突变体的初筛,工作量十分庞大。选取合适的抗生素作为初筛试剂,可大大减少筛选工作量,提高筛选效率。抗生素抗性筛选是基于微生物对抗生素产生耐药性发展起来的菌株选育改良技术,实践证明,操作简单,效果明显,在微生物药物领域得到广泛应用[23]。微生物的某些抗生素抗性突变会直接影响其次生产物的代谢调控系统[24],从而改变突变株代谢产物的生产水平和能力,因此,抗性筛选可用于有用产物生产优良菌株的选育和改良,在微生物药物领域抗生素抗性筛选技术原本主要用来筛选获取高产菌株[25],但近来发现,微生物的抗生素抗性突变还可赋予突变株新生次级产物的代谢生产能力。由于微藻和微生物具有相似的繁殖方式及代谢机制,所以,可以利用诱变和抗生素筛选相结合的方法,对微藻突变株进行筛选。关于抗生素抗性筛选提高产能的生物学机制的研究在链霉素和利福平霉素抗性突变株的筛选中具有良好的效果[26]。
抗生素通过不同的作用机理可以抑制或杀死细菌、真菌等不同的病原微生物,因此,可作为一种建立微藻无菌体系的试剂。倘若某种抗生素对微藻的生长没有影响,同时又具有良好的除菌效果,那么它将成为建立无菌体系的最佳途径。本研究发现,氨苄霉素、羧卞霉素、头孢霉素、硫酸新霉素等4种抗生素在试验浓度范围内不影响小球藻的生长。因此,可将其用于建立埃氏小球藻的无菌体系,考虑到病原菌的耐药性及更佳的除菌效果,可将几种抗生素混合使用,以达到最佳的使用效果。
研究抗生素对微藻生长的影响,在微藻的藻种改良及培养工作中都具有非常重要的作用,可为获得适合微藻规模化培养并联产微藻燃油的工业藻株提供参考。
[1]刘飞翔,刘伟平.基于能源安全与环境思考的生物质能产业发展[J].科技与产业,2009,9(10):26-28.
[2]薛智权,吕蕊,李宏,等.一株富油微藻的鉴定及其脂肪酸成分分析[J].天津农业科学,2016,22(8):1-5.
[3]赵宗保,华艳艳,刘波.中国如何突破生物柴油产业的原料瓶颈[J].中国生物工程杂志,2005,25(11):1-6.
[4]李翠芬,吴小情,尹爱国.小球藻产油脂培养条件的优化[J].河南农业科学,2014,43(10):156-159.
[5]郑金明.生物柴油原料评价及其混合性能改善研究 [D].北京:中国地质大学,2012.
[6]郑国清,王宝艾,杨自军,等.抗生素等药物添加剂的应用[J].河南农业科学,1991(3):28-29.
[7]钱文倩.微生物对异养小球藻生长及代谢产物影响的研究[D].上海:上海交通大学,2008.
[8]黄健,唐学玺,宫相忠,等.海洋微藻基因工程的选择标记[J].植物学报,2000,42(8):841-844.
[9]耿德贵,王义琴,李文彬,等.杜氏盐藻基因工程选择标记的研究[J].生物技术,2001,11(5):1-3.
[10]曹军平,费志清,刘必谦,等.金藻基因工程选择标记的研究[J].海洋科学,2001,25(7):6-8.
[11]刘滨扬.红霉素、环丙沙星和磺胺甲噁唑对羊角月牙藻的毒性效应及其作用机理[D].广州:暨南大学,2011.
[12]李杰,刘颖,黄洁虹,等.G418和氯霉素作为转基因盐藻的抗生素筛选标记[J].生物技术,2003,13(3):22-23.
[13]CHA T S,YEE W,AZIZ A.Assessment of factors affecting agrobacterium-mediated genetic transformation of the unicellular green alga,Chlorella vulgaris[J].World Journal Microbiology Bio-technology,2012,28(4):1771-1779.
[14]王长海,王逸云.小球藻基因工程选择标记研究[J].大连理工大学学报,2007(7):512-513.
[15]臧晓南,张学成,王高歌,等.节旋藻和螺旋藻对7种抗生素敏感性的比较研究[J].海洋学报,2004(5):85-86.
[16]李翠英,陈美兰,缪丽梅.螺旋藻在水产养殖中的应用[J].内蒙古农业科技,2003(Z2):174-175.
[17]王逸云.小球藻外源基因转化系统的建立及其表达植酸酶的研究[D].大连:大连理工大学,2005.
[18]GRUBBER M Y.In vitro expression of a cyanobacterical plasmid [J].Current Microbiology,1987,15:265-268.
[19]杨元,吴兴杰,陈建月.抗生素对两种竹芋组织培养的影响[J].天津农业科学,2012,18(6):121-124.
[20]齐红莉,杨广,刘金兰.抗卡那霉素单克隆抗体细胞株的筛选及间接 ELISA法的建立 [J].华北农学报,2010,25(2):121-124.
[21]雒淑珍,魏玉杰,赵继荣,等.利用卡那霉素筛选野罂粟转基因植株的技术研究[J].广东农业科学,2010,22(9):170-171.
[22]林伟,刘秀云.海洋微藻除菌及除菌与自然带菌微藻生长特点比较[J].海洋与湖沼,2000,31(6):647-652.
[23]孙玉雯,崔承彬.抗生素抗性筛选在微生物菌株选育中的作用[J].国际药学研究杂志,2008,35(3):213-217.
[24]SHINA L,TAKASE S,TAKAHASHI,et al.Novel detoxification ofthe trichothecenemycomxin deoxynivalenol by a soil bacterium i-solated by enrichnent culhzre [J].ApplEnviron Microbiol,1997,63(10):3825-3830.
[25]HOSOYA YOSHIKO,OKAMOTO SUSUMU,MURAMATSU H,et al.Acquisition of cer-tain streptanycin-resistant(str)mutations enhances antibiotic production in bacteria[J].Antinicrob Agents Chenoer,1998,42(8):2041-2047.
[26]INAOKA T,TAKAHASHI K,YADA H,et al.RNA polymerase mutation activates the production of a doanant antibiotic 3,3' -neotrehalosadianine via an autoinduction mechanisn in Bacillus sabtilis[J].J Biol Chen,2004,279(5):3885-3892.
Study on the Sensitivity of Chlorella emersonii to Eleven Kinds of Antibiotics
WANGYajun,ZHOUGuanghang,JI Chunli,XUE Jin'ai,LI Runzhi
(Institute ofMolecular Agriculture and Bioenergy,Shanxi Agricultural University,Taigu 030801,China)
To investigate the sensitivity of strain SXND-12(Chlorella emersonii)to 11 antibiotics,the study was conducted by spectrophotometry to evaluate the effects of 11 antibiotics(chloramphenicol,hygromycin,erythromycin,rifampicin,kanamycin,neomycin sulfate,cephalosporins,daqingmycin,ampicillin,carboxomycin and streptomycin etc.)on growth of Chlorella emersonii to determine concentration of antibiotics in isolation and purification of algae,which was harmless to algal cells and could inhibit the growth of associated bacteria.The results showed that Chlorella emersonii was less sensitive to cephalosporin,ampicillin,carboxymycin and neomycin sulfate,was more sensitive to rifampicin(concentration of 200 μg/mL(lethal dose)as well as 0.243 mmol/mL(lethal concentration)in the solid culture which could inhibit the growth of Chlorella emersonii,and was very sensitive to hygromycin, chloramphenicol,erythromycin,streptomycin,daqingmycin,kanamycin(lethal concentration in the solid culture reached 20 μg/mL(0.101 mmol/mL),50 μg/mL(0.155 mmol/mL),30 μg/mL(0.041 mmol/mL),50 μg/mL(0.086 mmol/mL),50 μg/mL(0.105 mmol/mL), 100 μg/mL(0.206 mmol/mL,respectively).This study laid a foundation for screening reagents for chlorella genetic engineering, mutagenesis screening early screening reagents,and establish a selection reagent for sterile systems.
Chlorella emersonii;antibiotics;sensitivity;growth inhibition
S949.93
A
1002-2481(2017)03-0379-07
10.3969/j.issn.1002-2481.2017.03.15
2016-12-06
国家“948”项目(2014-Z39);山西省煤基重点科技攻关项目(FT-2014-01)
王亚君(1990-),女,山西忻州人,在读硕士,研究方向:生物固碳与生物质综合利用。李润植为通信作者。
