枇杷叶熊果酸对大鼠肝星状细胞增殖抑制作用及对PPAR-γ、TGF-β1 表达的影响
2017-04-14张扬武罗伟生谭全肖王仕衍禤传凤
张扬武,罗伟生,陈 姗,黄 瑞,谭全肖,王仕衍,张 夏,禤传凤
(广西中医药大学第一附属医院脾胃科,广西 南宁 530001)
枇杷叶熊果酸对大鼠肝星状细胞增殖抑制作用及对PPAR-γ、TGF-β1 表达的影响
张扬武,罗伟生,陈 姗,黄 瑞,谭全肖,王仕衍,张 夏,禤传凤
(广西中医药大学第一附属医院脾胃科,广西 南宁 530001)
目的 研究枇杷叶熊果酸对大鼠肝星状细胞增殖的抑制作用及对PPAR-γ、TGF-β1表达的影响,探讨枇杷叶熊果酸抗肝纤维化的可能作用机制。方法 体外培养HSC-T6细胞,随机分为空白组,罗格列酮对照组,枇杷叶熊果酸低、中、高浓度组,分别干预24、48、72 h后,运用CCK-8法检测各组细胞的增殖情况;ELISA法检测各组细胞培养上清液中的Ⅰ型胶原蛋白含量;Real-time PCR法检测各组PPAR-γ、TGF-β1 mRNA的表达;免疫细胞化学法检测各组PPAR-γ、TGF-β1蛋白的表达。结果 CCK-8结果显示,随着熊果酸作用时间的延长,药物对细胞的增殖的抑制率增高(P<0.01);ELISA结果显示,随着熊果酸药物浓度的增加,上清中的Ⅰ型胶原蛋白含量下降(P<0.01);Real-time PCR结果显示,随着熊果酸药物浓度增加,PPAR-γ mRNA的表达量增加,TGF-β1 mRNA的表达量下降(P<0.01);免疫细胞化学结果显示,随着熊果酸药物浓度增加,PPAR-γ 蛋白的表达增强,TGF-β1 蛋白的表达减弱(P<0.01)。以上作用具有剂量-效应关系,高浓度组作用较对照组强。结论 枇杷叶熊果酸能够抑制HSC-T6细胞增殖,降低细胞外基质分泌,该机制可能与上调PPAR-γ 表达,下调TGF-β1表达有关。
枇杷叶熊果酸;肝星状细胞;肝纤维化;胶原;细胞外基质;细胞因子
肝纤维化是由于多种致病因素形成的慢性肝实质炎症、坏死导致肝脏纤维结缔组织的过度沉积、异常增生的一种慢性肝脏疾病。肝纤维化的进程中,肝星状细胞(hepatic stellate cell,HSC)的活化和细胞外基质(extracellular matrix,ECM)的分泌起着关键作用。熊果酸(ursolic acid,UA)是枇杷叶的功能物质之一,而且相对其它含有UA的中草药含量丰富。前期研究表明[1],UA具有降低转化生长因子-β1(transforming growth factor-β1,TGF-β1)的mRNA及蛋白表达,抑制TGF-β1与其受体的结合[2],诱导HSC-T6细胞凋亡并降低Bcl2/Bax比值,激活天冬氨酸特异的半胱氨酸蛋白酶3(cysteinyl aspartate-specific proteinase-3,Caspase-3)相关蛋白[3]的作用。过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor-γ ,PPAR-γ)是存在于HSC里的一种配体激活转录因子。PPAR-γ通过阻断TGF-β1/TβR-Ⅰ信号,抑制Smad磷酸化,进一步抑制Smad靶基因的转录,从而抑制ECM生成[4]。UA是否也可通过激活PPAR-γ而抑制TGF-β1及其信号通路下游因子的表达而发挥抗肝纤维化的作用?因此,本实验通过体外培养HSC-T6细胞,运用CCK-8法检测UA对HSC-T6细胞的增殖情况,ELISA法检测各组细胞培养上清液中的(Collagen Ⅰ,Col Ⅰ)含量,Real-time PCR法检测各组PPAR-γ、TGF-β1 mRNA的表达,免疫细胞化学法检测各组PPAR-γ、TGF-β1蛋白的表达,探讨UA抗肝纤维化的可能作用机制。
1 材料
1.1 细胞系 HSC-T6细胞购自中国科学院昆明动物研究所细胞库。
1.2 药物与试剂 枇杷叶熊果酸(批号:160623)购自上海融禾医药科技发展有限公司;罗格列酮(批号:HY-17386)购自MCE上海皓元生物医药科技有限公司;FBS、DMEM高糖培养基购自加拿大维森特公司;CCK-8试剂盒购自日本Dojindo公司;Col-Ⅰ ELISA试剂盒购自基尔顿(上海)生物科技有限公司;总RNA提取试剂盒、反转录试剂盒、荧光定量PCR试剂盒、PPAR-γ、TGF-β1、GAPDH内参引物均购自宝生物工程 (大连) 有限公司;PPAR-γ、TGF-β1第一抗体购自美国Abcam公司;兔SP检测试剂盒、DAB显色剂购自北京中杉金桥生物技术有限公司。
1.3 主要仪器 二氧化碳培养箱(371-184L,苏州);低速离心机(DL-5M,湖南);恒温水浴槽(SY-1220,美国);倒置显微镜(IX71,日本);生物安全柜(BSC-1600IIA2,苏州);超微量紫外可见分光光度计(UV1901PC,上海);酶标仪(Epoch Biotek,美国);实时荧光定量PCR仪(PIKOREAL,美国)。
2 方法
2.1 细胞培养 大鼠肝星状细胞HSC-T6运用10%胎牛血清的高糖DMEM培养基置于5% CO2、37℃的培养箱中培养,每3~4天传代1次,取处于对数期生长期的细胞用于实验。实验分组为:空白组、罗格列酮对照组、UA低、中、高浓度组。
2.2 CCK-8法检测HSC-T6细胞增殖 取处于对数生长期的细胞消化进行细胞计数,调整细胞密度为2×107·L-1,吹悬打匀后以每孔100 μL接种于96孔培养板,设置空白对照组(无药培养基)、罗格列酮对照组[4](15 μmol·L-1罗格列酮) 、UA低浓度组(10 μmol·L-1UA)、UA中浓度组(25 μmol·L-1UA)、UA高浓度组(40 μmol·L-1UA),每组设定5个复孔。细胞接板24 h完全贴壁后弃去旧培养基,分别加入不同药物的工作液,干预24、48、72 h后弃培养基,加入新的完全培养基后每孔加入10 μL的CCK-8溶液,置于37℃培养箱孵育1 h,多功能酶标读数仪上测定450 nm处的OD值。按以下公式计算不同药物浓度对HSC-T6细胞的增殖抑制率:抑制率(IR)/%=[(空白对照组OD值-实验组OD值)/(空白对照组OD值-空白孔OD值)]×100%,根据不同药物浓度对细胞增殖的能力,选择后续实验药物最佳作用时间。
2.3 ELISA法检测细胞培养上清液中Col-Ⅰ含量 取处于对数期细胞以10×107·L-1密度,每孔2.5 mL接种于6孔板中,每组设3个复孔,细胞贴壁后按CCK-8法中的各组药物浓度干预细胞72 h后收集细胞培养上清液。先在各小试管中稀释标准品,然后设置空白孔、待测样品孔进行加样,温育30 min,用30倍浓缩液稀释后进行洗涤,每孔加入酶标试剂后再进行温育30 min,重复洗涤后加入显色剂显色15 min,加入终止液终止反应,以空白孔调零于450 nm波长依次测量各孔的吸光度(OD值)。以标准物的浓度为横坐标,OD值为纵坐标,在坐标纸上绘出标准曲线,根据样品OD值由标准曲线查出相应的浓度。
2.4 Real-time PCR法检测细胞内PPAR-γ、TGF-β1 mRNA的表达 细胞按10×107·L-1密度,每孔2.5 mL接种于6孔板中,每组设3个复孔,细胞贴壁后按CCK-8法中的各组药物浓度干预72 h后TRIzol法提取总RNA,逆转录合成cDNA,SYBR®Premix Ex TaqTM Ⅱ说明书进行Real -time PCR检测,以GAPDH引物为内参。GAPDH上游5′-ATGGGAAGCTGGTCATCAAC-3′,GAPDH下游5′-GTGGTTCACACCCATCACAA-3′;PPAR-γ上游5′-CTGTTTTATGCTGTTATGGGT-3′;PPAR-γ下游5′-GTCAAAGGAATGGGAGTGGTC-3′;TGF-β1上游5′-CTGGGGCCCTGCCCCTACAT-3′,TGF-β1下游5′-CTTGGGCTTGCGACCCACGT-3′。两步法反应条件如下:95℃ 30 s;95℃ 5 s,60℃ 30 s,44个循环。读取Ct值,以2-△△Ct表示目的基因mRNA的表达水平(-△△Ct计算公式:-△△Ct XY=Ct XY-Ct GAPDH Y-Ct X对照,其中X代表基因,Y代表处理因素)。
2.5 免疫细胞化学法检测细胞内PPAR-γ、TGF-β1蛋白的表达 采用免疫组化SP法,将对数生长期的HSC-T6细胞接种于放有盖玻片的6孔板中,每组设2个复孔,待细胞贴壁生长后,不同药物分组干预。药物与细胞共孵育72 h后,取出盖玻片置于载玻片上,用4%多聚甲醛固定,3% H2O2消除内源性过氧化氢酶,正常血清封闭后分别加入鼠抗人PPAR-γ、TGF-β1单克隆抗体,4℃孵育过夜,随后加入生物素二抗工作液及辣根酶标记链霉卵白素工作液,经DAB显色,脱水、封片,光学显微镜下观察。结果判读:在每组的2 片细胞盖玻片中随机选取6个有意义的高倍镜视野进行观察,可见细胞质呈特异性棕褐色阳性染色,阳性表达率/%=(500个细胞内阳性细胞数/500个细胞数)×100%[5]。
3 结果
3.1 HSC-T6细胞的增殖情况 不同药物浓度分别干预5组细胞24、48、72 h后CCK-8分析发现,UA具有剂量-效应关系抑制HSC-T6细胞的增殖,在72 h抑制率达到最大;与对照组比较,UA高浓度组抑制作用较强,但差异无统计学意义(P>0.05),中、低浓度组的抑制率较低(P<0.05)。后续实验采用72 h干预时间。见Tab 1。
3.2 各组细胞培养上清液Col-Ⅰ含量比较 药物干预细胞72 h后,与空白组比较,Col-Ⅰ含量明显低于空白组(P<0.01);与对照组比较,UA高浓度组Col-Ⅰ含量低于对照组(P<0.01),低、中浓度组Col-Ⅰ含量高于对照组(P<0.01)。见Tab 2。
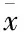
Tab 1 Effect of different group on OD value of HSC-T6 cells(± s,n=5)
*P<0.05,**P<0.01vsblank;#P<0.05,##P<0.01vscontrol
Tab 2 Comparison of content of Col-Ⅰ in different groups(±s,n=3)
**P<0.01vsblank;##P<0.01vscontrol
3.3 各组细胞PPAR-γ、TGF-β1 mRNA的表达比较 药物干预细胞72 h后,与空白组比较,药物干预组中PPAR-γ mRNA的表达量均明显增加(P<0.01),TGF-β1 mRNA的表达量均降低(P<0.05);与对照组比较,UA高浓度组PPAR-γ mRNA的表达量明显高于对照组(P<0.01),低浓度组明显低于对照组(P<0.01),中浓度组表达量低于对照组,但差异无统计学意义(P>0.05);与对照组比较,UA低浓度组TGF-β1 mRNA的表达量明显增加(P<0.01),UA中浓度组TGF-β1 mRNA的表达量增加,但差异无统计学意义(P>0.05),高浓度组表达量降低,但差异无统计学意义(P>0.05)。见Fig 1、Fig 2。
3.4 各组细胞PPAR-γ、TGF-β1蛋白的表达差异 药物干预72 h后,不同组别细胞中均出现棕褐色染色,与空白组比较,药物干预组PPAR-γ蛋白的表达量均增加,TGF-β1蛋白的表达量均减少(P<0.01);与对照组比较,UA低、中浓度组PPAR-γ蛋白的表达量均低于对照组,UA高浓度组高于对照组(P<0.01);与对照组比较,UA低、中浓度组TGF-β1蛋白的表达量均高于对照组,UA高浓度组低于对照组(P<0.01)。见Tab 3、Fig 3、4。
**P<0.01vsblank group;##P<0.01vscontrol group

Fig 2 Comparison of TGF-β1 mRNA expression in different groups(±s,n=3)
*P<0.05,**P<0.01vsblank group;##P<0.01vscontrol group
Tab 3 Percentage of PPAR-γ protein and TGF-β1 protein expression in different groups(±s,n=6)
**P<0.01vsblank;##P<0.01vscontrol
4 讨论
肝纤维化是各种肝病发展成肝硬化的必经病理过程。在我国常以乙型或丙型病毒性肝炎迁延不愈所致,30%慢性丙型肝炎患者最终出现严重的肝纤维化或肝硬化病变。肝病的治疗重点转移到如何控制肝病的进展,防止肝纤维化以致肝硬化的发生、发展已成为国内外生物医学研究的热点之一[6]。肝纤维化的特征表现为以胶原为主的ECM的降解减少,合成增多,沉积于肝内过度表达从而造成假小叶的形成与肝小叶的结构重建,而HSC的活化与增殖是ECM产生的主要来源,在肝纤维疾病的进程中起着绝对作用[7]。肝纤维化状态下ECM的质和量会发生明显变化,Ⅰ型胶原在肝脏中的含量比正常值高3~8 倍。所以我们可以认为HSC 细胞的分泌功能以Ⅰ 型胶原表达水平的测定来直接地反映[8]。前期实验研究表明,UA在体内具有抗菌、抗炎、抗病毒、抗癌、免疫调节、镇静、肝损伤保护等多种药理作用[9],但在抗肝纤维化的报道还比较少。本实验中,UA对HSC-T6细胞的抑制率随着浓度升高而升高,与对照组比较,UA高浓度干预组抑制率更高,说明UA具有明显的抑制HSC-T6细胞增殖的作用,并呈剂量-效应的关系。ELISA测定各组细胞上清液Ⅰ型胶原含量实验中发现,UA能够使Ⅰ型胶原降解,在UA高浓度组中该作用最为明显,说明UA能够抑制HSC-T6细胞中的胶原分泌。

Fig 3 Expression of PPAR-γ protein in different groups(×200)

Fig 4 Expression of TGF-β1 protein in different groups(×200)
肝纤维化的形成是一个多因素、多细胞参与的复杂过程。肝纤维化过程中,HSC的激活是其中心环节。PPAR-γ在配体作用下通过启动或参与复杂的信号通路发挥众多生物效应,但目前对PPAR-γ与TGF-β-Smad信号交联及其对下游因子在肝纤维化细胞信号传导通路中作用知之甚少。有研究表明,PPAR-γ活化后对c-Ski始发于转录水平的上调,并提示c-Ski可能是PPAR-γ的又一靶基因[10]。c-Ski是TGF-β的smad途径的阻遏子,其可以通过阻止smad2、smad3磷酸化,以及阻止磷酸化smad3与smad4结合形成有活性的smad复合物及募集大量辅抑制子而阻断TGF-β/smad途径对靶基因的转录调控[11]。TGF-β激活能够促进HSC增殖、产生胶原,同时抑制细胞外基质的降解,加速肝纤维化发展。罗格列酮可作为PPAR-γ的特异性诱导剂,通过上调PPAR-γ表达,下调TGF-β表达而抑制HSC细胞的活化,实现肝纤维化的逆转[12]。本实验研究发现,UA能够使HSC-T6细胞中的PPAR-γ基因和蛋白表达水平上调,下调TGF-β1基因和蛋白的表达,并且存在剂量依赖性,这可能是UA抗肝纤维的分子机制之一。
综上所述,UA能够抑制HSC-T6细胞增殖,降解ECM的沉积,说明UA具有抗肝纤维化的作用,其中机制可能与上调PPAR-γ表达,下调TGF-β1表达有关。肝纤维化的调控机制非常复杂,UA抗肝纤维化的作用机制有待进一步研究,为UA抗肝纤维化提供实验依据。
(致谢:本研究全部在广西中医药大学第一附属医院分子生物学实验室完成,特此感谢!)
[1] Murakami S, Takashima H, Sato-Watanabe M, et al. Ursolic acid, an antagonist for transforming growth factor(TGF)-beta1[J].FEBSLett,2004,566(1-3):55-9.
[2] 欧阳灿晖,朱 萱,张焜和,等.熊果酸对肝纤维化大鼠肝组织TGF-β1和α-SMA表达的影响[J]. 世界华人消化杂志,2009,17(22):2237-43.
[2] Ouyang C H, Zhu X, Zhang K H, et al.Effects of ursolic acid on the expression of transforming growth factor-β1 and alphasmooth muscle actin in fi brotic liver in rats[J].WorldChinJDigestol,2009,17(22):2237-43.
[3] 申月明,朱 萱,张昆和,等.熊果酸对肝星状细胞增殖与凋亡的影响[J]. 中华肝脏病杂志,2008,16(4): 298-301.
[3] Shen Y M, Zhu X, Zhang K H, et al. Effect of ursolic acid on proliferation and apoptosis of hepatic stellate cellsinvitro[J].ChinJHepatol,2008,16(4): 298-301.
[4] 蔡玲燕,刘 菲. PPARγ对肝纤维化中肝星状细胞相关信号通路的影响[J].国际内科学杂志,2009,36(7):417-9.
[4] Cai L Y, Liu F. Effect of PPARγ on signal transduction of hepatic stellate cells in liver fibrosis[J].IntJInternMed,2009,36(7):417-9.
[5] 谢金玲,赵 川,李俊萱,等. 白花丹醌对人肝癌细胞HepG2侵袭和凋亡的影响[J].中国药理学通报,2016,32(5):687-91.
[5] Xie J L, Zhao C, Li J X, et al.Effect of plumbagin on invasion and apoptosis of human hepatocellular carcinoma cell line HepG2[J].ChinPharmacolBull,2016,32(5):687-91.
[6] Garcia-Tsao G,Sanyal A J,Grace N D, et al. Prevention and management of gastroesophageal varices and variceal hemorrhage in cirrhosis[J].Hepatology, 2007,46(6): 922-38.
[7] Xu T, Ni M M, Xing-Li, et al.NLRC5 regulates TGF-β1-induced proliferation and activation of hepatic stellate cells during hepatic fibrosis[J].BiochemCellBiol,2016,1(70):92-104.
[8] Friedman S L. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury[J].JBiolChem,2000,275(4):2247-50.
[9] Kiran M S, Viji R I, Sameer Kumar V B, et al. Modulation of angiogenic factors by ursolic acid[J].BiochemBiophysResCommun,2008,371(3):556-60.
[10]李工博,李 军,曾益军,等.PPARγ对TGFβ/smad信号通路阻遏子c-Ski的上调[J].生理学报,2011,63(1):62-8.
[10]Li G B, Li J, Zeng Y J,et al. PPARγ up-regulates TGFβ/smad signal pathway repressor c-Ski[J].ActaPhysiolSin,2011,63(1):62-8.
[11]Macias-silva M, Li W, Leu J I,et al.Up-regulated transcriptional repressors SnoN and Ski bind Smad proteins to antagonize transforming growth factor-beta signals during liver regeneration[J].JBiolChem,2002,277(32): 28483-90.
[12]康 谊,曾艳丽,魏君锋,等. 罗格列酮抑制大鼠肝纤维化机制研究[J].中华实用诊断与治疗杂志,2013,27(07):676-9.
[12]Kang Y, Zeng Y L, Wei J F, et al. Mechanism of rosiglitazone inhibiting liver fibrosis in rats[J].JChinPractDiagnTher,2013,27(07):676-9.
Effects of ursolic acid from loquat leaves on proliferation inhibition and expression of PPAR-γ, TGF-β1 in rat hepatic stellate cells
ZHANG Yang-wu, LUO Wei-sheng, CHEN Shan, HUANG Rui,TAN Quan-xiao, WANG Shi-yan, ZHANG Xia, XUAN Chuan-feng
(DeptofStomachandSpleen,theFirstAffiliatedHospitalofGuangxiUniversityofChineseMedicine,Nanning530001,China)
Aim To investigate the effects of ursolic acid from loquat leaves on proliferation inhibition and expression of PPAR-γ 、TGF-β1 in rat hepatic stellate cells, so as to explore the mechanism of anti-hepatic fibrosis of UA.Methods HSC-T6 cells were randomly divided into blank group, rosiglitazone control group, the low, medium and high concentration of UA group to detect the cell proliferation inhibition by CCK-8 after 24,48,72 h. The content of type collagen Ⅰ in cell culture supernatant of each group was examined by ELISA. The expression of PPAR-γ mRNA, TGF-β1 mRNA in HSC-T6 cells exposured were examined by real-time quantitative PCR. Effects on HSC-T6 PPAR-γ and TGF-β1 protein from each group were detected by immunocytochemical method.Results CCK-8 results showed that the inhibitory rate of UA on cell proliferation increased with the prolongation of drug action time(P<0.01). ELISA results showed that with the increase of the concentration of UA, the content of type Ⅰ collagen content decreased(P<0.01). Real-time PCR results showed that with the increase of the concentration of UA, and the expression of PPAR-γ mRNA increased, the expression of TGF-β1 mRNA decreased(P<0.01). The results of immunocytochemistry showed that the expression of PPAR-γ protein was increased, and the expression of TGF-β1 protein was decreased with the increase of the concentration of UA(P<0.01).All effects mentioned above were dose-dependent. Moreover, the effects in the high concentration groups were stronger than those in control group.Conclusion UA can inhibit the proliferation of HSC-T6 cells, Which may be associated with the up-regulation of PPAR-γ expression and the down-regulation of TGF-β1 expression.
ursolic acid from loquat leaves;hepatic stellate cells; liver fibrosis; collagen;extracellular matrix; cytokine
时间:2017-3-13 8:38
http://kns.cnki.net/kcms/detail/34.1086.R.20170324.1247.028.html
2016-12-20,
2017-01-14
国家自然科学基金地区资助项目(No 81360530);国家自然科学基金地区资助项目(No 81660779);广西中医药大学硕士研究生科研创新重点项目(No YJS201617)
张扬武(1987-),男,硕士生,研究方向:消化系统疾病的临床和基础,E-mail:1013194142@qq.com; 罗伟生(1959-),男,博士,教授,主任医师,研究方向:消化系统疾病的临床和基础,通讯作者,E-mail:4011188@qq.com
10.3969/j.issn.1001-1978.2017.04.014
A
1001-1978(2017)04-0517-05
R-332;R284.1;R322.47;R329.24;R392.12;R575.202.2;R977.6
