过表达MicroRNA-199a促进TNFα诱导的脂肪细胞凋亡
2017-04-06齐仁立王琪吴永江王敬黄金秀杨飞云
齐仁立 王琪 吴永江 王敬 黄金秀 杨飞云
(1. 重庆市畜牧科学院,重庆 402460;2. 农业部养猪科学重点实验室,重庆 402460,3. 西南大学荣昌校区,重庆 402460)
过表达MicroRNA-199a促进TNFα诱导的脂肪细胞凋亡
齐仁立1,2王琪1吴永江3王敬1黄金秀1,2杨飞云1,2
(1. 重庆市畜牧科学院,重庆 402460;2. 农业部养猪科学重点实验室,重庆 402460,3. 西南大学荣昌校区,重庆 402460)
旨为探明microRNA-199a(miR-199a)对脂肪细胞凋亡的影响及核转录因子κB(nuclear factor κB,NF-κB)在其中的作用。培养3T3-L1脂肪细胞,使用肿瘤坏死因子α(tumor necrosis factor α,TNFα)诱导细胞凋亡。在此基础上分别使用NF-κB阻断剂PDTC和miR-199a mimic处理细胞,判断过表达miR-199a对TNFα诱导的脂肪细胞凋亡的影响及NF-κB在其中的作用。使用流式细胞仪分析细胞凋亡率,试剂盒检测Caspase3/7酶活,双荧光素酶报告系统检测NF-κB活性,qRT-PCR 检测miR-199a的表达水平,Western blotting检测NF-κB相关蛋白变化。结果显示,TNFα可以诱导分化的脂肪细胞凋亡并同时激活NF-κB,激活的NF-κB在TNFα诱导的脂肪细胞凋亡中发挥负调控作用。在脂肪细胞过表达miR-199a明显抑制NF-κB的激活进而显著促进TNFα诱导的凋亡。miR-199a通过调控NF-κB活性参与TNFα诱导的脂肪细胞凋亡。
MiR-199a;脂肪细胞;凋亡;TNFa;NF-κB
白色脂肪组织被视为哺乳动物的“燃料库”,脂肪细胞的数量、尺寸和状态对于机体能量平衡具有重要的影响[1]。脂肪的过度积累会导致肥胖症、Ⅱ型糖尿病、胰岛素抵抗等多种代谢疾病。此外,脂肪组织还是一种重要的分泌器官,它能分泌产生瘦素、抵抗素和脂联素等多种蛋白因子,调控机体不同的代谢反应[2]。当生长环境剧烈改变或者受到刺激时(射线、药物、细胞因子等)细胞会启动凋亡—由多基因调控的细胞程序性死亡[3]。关于脂肪细胞凋亡的分子机制研究有助于我们更好地了解脂肪的生长和发育机制,也为肥胖症等疾病的治疗提供新的视角。
肿瘤坏死因子α(Tumor necrosis factor α,TNFα)能够诱导包括脂肪细胞在内的多种细胞凋亡[4-6]。TNFα作为配体与受体蛋白TNFR结合,激活死亡受体途径并传递凋亡信号,最终激活Caspase凋亡蛋白级联引起细胞凋亡。核因子κB(nuclear factor κB,NF-κB)是一种重要的核转录因子,细胞静息状态下NF-κB位于胞浆中没有活性。在受到药物、放射线、或者TNFα等蛋白因子的刺激时,NF-κB会被激活并移入核内调控靶基因的转录[7]。激活的NF-κB对细胞凋亡具有重要影响,多数情况下发挥负调控作用,但是某些情况下也会促进凋亡[8,9]。
MicroRNA(miRNA)是一类长度为18-25 nt的非编码RNA,在真核生物体内广泛表达[10]。通常情况下,miRNA通过与靶基因3'UTR区互补结合,引导沉默复合体降解靶mRNA或抑制其翻译[11]。一个miRNA通常可以调控多达数百个靶基因,进而参与调控细胞的增殖、分化、激活和凋亡等不同生命事件。
最近的研究发现miR-199a对于脂肪细胞增殖和分化具有调控作用[12],并且参与了一些癌细胞的凋亡过程[13]。但是miR-199a是否参与了脂肪细胞凋亡的产生和发展还不清楚。此外,miR-199a与NF-κB通路也有一定作用关系。脂多糖(LPS)激活癌细胞中的NF-κB并抑制miR-199a的表达,而使用阻断剂PDTC抑制NF-κB激活则补偿性提高miR-199a的表达,说明NF-κB与miR-199a之间存在互作关系[14]。本实验试图探明miR-199a对脂肪细胞凋亡的影响并分析NF-κB在其中扮演的角色,旨在对miRNA功能的研究和脂肪细胞生长发育调控的研究提供参考。
1 材料与方法
1.1 材料
3T3-L1前体脂肪细胞购自于中科院上海细胞库,TNFα购自于Sigma-Aldrich公司(美国),PDTC购自于北京碧云天生物技术公司,miR-199a mimic由广州锐博生物技术公司设计合成,TUNEL细胞凋亡检测试剂购自于Life technology公司(美国),RNA提取试剂和qRT-PCR反应试剂盒购自于TaKaRa公司(日本),NF-κB luciferase reporter plasmid和Casepase3/7活性检测试剂盒购自于Promega公司(美国)。
1.2 方法
1.2.1 细胞培养和处理 接种并培养3T3-L1前体脂肪细胞,待细胞密度达到70%时开始生脂诱导分化。分化6 d的细胞使用TNFα(终浓度20 ng/mL,)处理6 h然后检测细胞凋亡率,空白处理作对照。为了检测NF-κB对凋亡的影响,使用NF-κB通路阻断剂 PDTC处理细胞1 h,再用TNFα处理6 h,然后检测NF-κB活性和细胞凋亡率。为了检测miR-199a对凋亡的影响,使用特异性miR-199a mimic(60 nmol/L)转染细胞,过表达细胞内miR-199a。mimic转染24 h后再用TNFa处理,检测NF-κB活性和细胞凋亡率。
1.2.2 细胞凋亡分析 TUNEL(TdT-mediated dUTP nick end labeling)可以特异性结合细胞DNA断裂,是细胞凋亡的重要标志。收集5×104个细胞,1%多聚甲醛固定5 min,PBS漂洗,70%乙醇通透细胞4 h,然后使用TUNEL反应液孵育细胞1 h,最后使用PI染色20 min并马上用流式细胞仪(贝克曼公司,美国)检测凋亡细胞,统计TUNEL阳性细胞(凋亡细胞)占总细胞的比例。
1.2.3 Caspase3/7 活性检测 Casepase3/7是一种关键的凋亡蛋白,是细胞凋亡通路的终端执行因子。细胞中Caspase3/7的酶活性通过试剂盒检测,具体操作根据生产商的说明进行。细胞转染48 h后,弃去培养基,PBS溶液漂洗3次,离心收集细胞,加入一定体积的Caspase-Glo®3/7反应液室温条件下孵育2 h,然后使用多功能荧光酶标仪(Biotek公司,美国)检测光密度。
1.2.4 RNA提取和qRT-PCR MiR-199a的表达水平使用实时荧光定量PCR(qRT- PCR)检测 使用small RNA试剂提取细胞中的总RNA,使用1%琼脂糖凝胶电泳检测RNA完整性,使用核酸定量分析仪分析RNA纯度。合格的RNA使用PrimeScriptTMRT reagent Kit试剂盒反转录为cDNA。使用Step One荧光定量PCR仪(AB公司,美国)进行PCR反应,反应条件:95℃预变性30 s;(95℃ 5 s 60℃ 35 s)40个循环。U6为参照基因,2-△△CT法计算基因的相对表达量。
1.2.5 Western blotting收获细胞并使用RIPA裂解液提取总蛋白 BCA法蛋白检测试剂盒(康为公司,中国)检测蛋白浓度。12%的SDS-PAGE胶进行蛋白分离,然后转印到0.22 μm PVDF膜(Millipore公司,美国)上。转印完的PVDF膜用5%的脱脂牛奶溶液室温封闭2 h。使用相应的一抗和二抗溶液孵育PVDF膜。最后用ECL化学发光试剂(Millipore公司,美国)和自动发光曝光仪(Bio-Rad公司,美国)进行拍照分析。试验用的抗体购自于CST公司(美国),Tublin作为内参抗体。
1.2.6 NF-κB活性检测 使用双荧光素酶报告系统检测NF-κB活性。双荧光素酶检测试剂盒和NF-κB luciferase reporter plasmid购自于Promega公司(美国),具体操作步骤参考说明书进行。
1.2.7 数据统计分析 使用SPSS19.0分析软件包(IBM公司,美国)对试验数据进行单因素方差分析(one-way ANOVA)或者student's t-test,P < 0.05表示差异显著,P < 0.01表示差异极显著。
2 结果
2.1 抑制NF-κB激活增强TNFα诱导的脂肪细胞凋亡
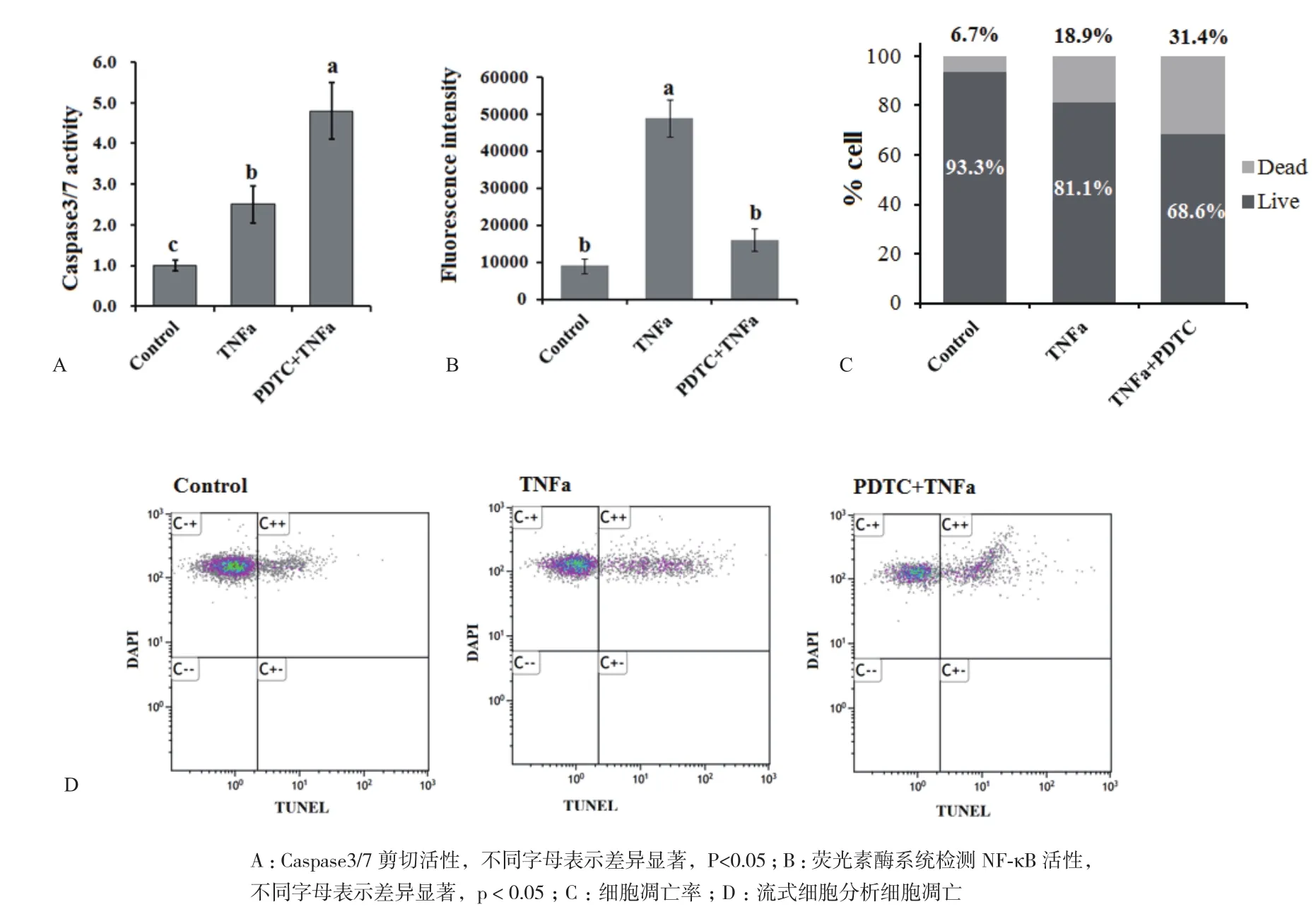
图1 抑制NF-κB活性促进TNFα诱导的脂肪细胞凋亡
图1 显示TNFα处理6 h能够诱导分化的脂肪细胞凋亡,但是诱导作用较为温和,同时TNFα能够快速激活细胞内的NF-κB。PDTC是一种NF-κB阻断剂,它显著抑制了TNFα对 NF-κB的激活作用(图1-B)。使用PDTC处理细胞1 h后再用TNFα处理细胞,Caspase3/7酶活性显著提高(图1-A),凋亡细胞数显著增加(图1-C和1-D)。这说明抑制NF-κB活性能够促进TNFa诱导的脂肪细胞凋亡。
2.2 表达miR-199a增强TNFα诱导的脂肪细胞凋亡定量PCR结果显示mimic转染使脂肪细胞内miR-199a的表达水平增加了42.8倍(图2-A)。转染mimic 24 h后再用TNFa处理细胞,凋亡细胞数和caspase3活性均显著高于只用TNFα处理的细胞(图2-B,2-C和2-D)。与空白对照组相比,单独转染mimic对于脂肪细胞凋亡没有显著影响。表明过表达miR-199a能够增强TNFα诱导的脂肪细胞凋亡。
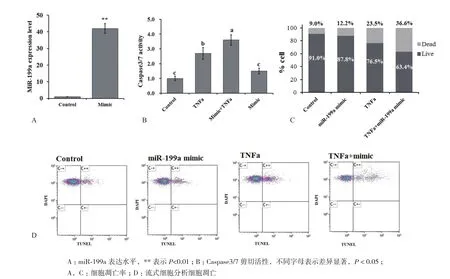
图2 过表达miR-199a促进TNFα诱导的脂肪细胞凋亡
2.3 过表达miR-199a抑制TNFα激活NF-κB
我们推测miR-199a调控TNFα诱导的脂肪细胞凋亡可能与NF-κB激活有关。使用双荧光素酶系统检测NF-κB活性(图3-A)及Western blotting 检测NF-κB蛋白水平(图3-B),结果表明miR-199a mimic 转染显著抑制了NF-κB的激活,明显地减少磷酸化的NF-κB抑制物激酶(inhibitor of nuclear factor kappa-B kinase α,IKKα)蛋白和磷酸化的P65蛋白。
3 讨论
凭借对众多靶基因的调控,microRNA在真核细胞的不同生命活动中扮演了重要的调控角色。在脂肪细胞上,已经有数十个miRNA的表达和调控功能被探明,如miR-143和Let-7等[15-18]。并且随着研究方法和技术的不断进步,会有更多涉及脂肪细胞生长和发育的microRNA被发现和确认。
miR-199a在哺乳动物的多个组织都有表达,涉及免疫反应、信号传导、肌肉发育、癌细胞生长等不同分子事件[19-21]。最近的研究表明,miR-199a对脂肪细胞增殖和分化也由明显的调控作用[12]。在猪前体脂肪细胞上过表达miR-199a能够促进细胞增殖,抑制其向成熟脂肪细胞分化。我们前期实验研究发现miR-199a对肌细胞生脂也有影响,过表达miR-199a显著抑制了骨骼肌细胞中脂肪沉积(数据未发表)。在一些肿瘤细胞上,miR-199a还显示了细胞凋亡调控的能力。然而,miR-199a是否参与脂肪细胞凋亡的形成和发展还不清楚。本实验结果确认过表达miR-199a能够通过抑制NF-κB活性促进TNFa诱导的脂肪细胞凋亡。相似的研究发现激活的NF-κB 能够抑制miR-199a表达;反之,高水平的miR-199a通过靶向调控IKKα影响NF-κB的活性[22]。IKKα是NF-κB的上游激酶控制着P65的磷酸化,本实验同样发现过表达miR-199a明显降低了IKKα的磷酸化水平继而影响了P65的磷酸化和NF-κB的活性。这些实验结果显示miR-199a和NF-κB之间存在负反馈调节机制。
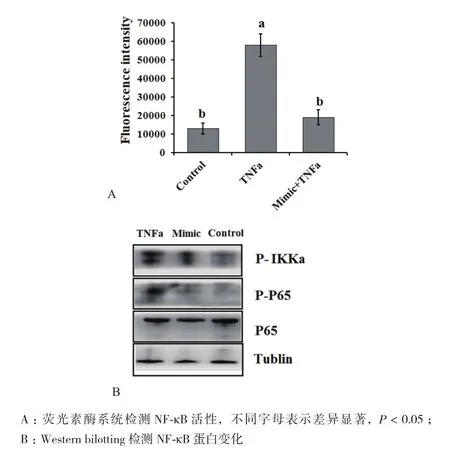
图3 过表达miR-199a抑制TNFα激活NF-κB
多数研究者认为 NF-κB 具有凋亡抑制作用,敲除 RelA 基因会显著提高胚胎成纤维细胞对于 TNFα介导的凋亡的敏感度。TNFα 能促使RelA-/-小鼠出现肝细胞大量凋亡,并最终导致小鼠死亡[23,24]。但是也有实验报道了NF-κB的促凋亡作用。例如,Ryan等[25]实验发现抑制或者减少 NF-κB 的活性,则会阻断 P53 引起的凋亡。本实验确认过表达miR-199a能够抑制脂肪细胞中NF-κB的活性,增加脂肪细胞对TNFα介导的凋亡的敏感度。TNFα快速激活脂肪细胞中的NF-κB,但是活化的NF-κB反馈抑制了TNFα诱导的脂肪细胞凋亡。近年来,一些miRNA陆续被探明涉及NF-κB的活性调控[26],这些miRNA也有较大可能性参与细胞凋亡的形成和发展,但是还需要更多的实验验证。
4 结论
核转录因子NF-κB在TNFa诱导的脂肪细胞凋亡中扮演负调控角色,而miR-199a可以通过调控NF-κB的活性促进TNFα诱导的脂肪细胞凋亡。
[1]Rosen ED, Macdougald OA. Adipocyte differentiation from the inside out[J]. Nature Reviews Molecular Cell Biology, 2006, 7:885-896.
[2]Harp JB, Franklin D, Vanderpuijea AA, Gimbleb JM. Differential expression of signal transducers and activators of transcription during human adipogenesis[J]. Biochemical and Biophysical Research Communications, 2001, 281:907-912.
[3]Hu Q, Wu D, Chen W, et al. Molecular determinants of caspase-9 activation by the Apaf-1 apoptosome[J]. PNAS, 2014, 111:16254-16261.
[4]Ebmeyer J, Leichtle A, Hernandez M, et al. TNFa deletion alters apoptosis as well as caspase 3 and 4 expression during otitis media[J]. BMC Immunology, 2011, 12:12.
[5]林亚秋, 陈国柱, 卢建雄, 等. TNFa对培养大鼠脂肪细胞凋亡的影响[J]. 畜牧兽医学报, 2005, 36(8):823-827.
[6]Haake SM, Dinh CT, Chen SB, et al. Dexamethasone protects auditory HCs against TNF alpha-initiated apoptosis via activation of PI3K/Akt and NF-kappa B signaling[J]. Hearing Research, 2009, 255(1-2):22-32.
[7]Gilmore TD. Introduction to NF-κB:players, pathways, perspectives[J]. Oncogene, 2006, 25:6680-6684.
[8]Beg AA, Baltimore D. An essential role for NF-B in preventing TNFα induced cell death[J]. Science, 1996, 274:782-784.
[9]Baichwal VR, Baeuerle PA. Apoptosis:Activate NF-kB or die?[J]. Curr Biol, 1997, 7(2):94-96.
[10]Zeng Y, Yi R, Cullen BR. MicroRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms[J]. PNAS, 2003, 100:9779-9784.
[11]Friedman RC, Farh KK, Burge C B, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs[J]. Genome Res,2009, 19:92-105.
[12]Shi XE, Li YF, Jia L, et al. MicroRNA-199a-5p affects porcine preadipocyte proliferation and differentiation[J]. Int J Mol Sci, 2014, 15:8526-8538.
[13]Tsukigia M, Bilima V, Yuukia K, et al. Re-expression of miR-199a suppresses renal cancer cell proliferation and survival by targeting GSK-3β[J]. Cancer Letters, 2012, 315:189-197.
[14]Duan QL, Wang XX, Gong W, et al. ER stress negatively modulates the expression of the mir-199a/214 cluster to regulates tumor survival and progression in human hepatocellular cancer[J]. PLoS One, 2012, 9:e31518.
[15]Kajimoto K, Naraba H, Iwai N. MicroRNA and 3T3-L1 preadipocyte differentiation[J]. RNA, 2006, 12:1626-1632.
[16]Sun TW, Fu MG, Bookout AL, et al. MicroRNA let-7 regulates 3t3-l1 adipogenesis[J]. Mol Endocrinol, 2009, 23:925-931.
[17]Esau C, Kang XL, Peralta E, et al. MicroRNA-143 regulates adipocyte differentiation[J]. The journal of Biological Chemistry, 2004, 279:52361-52365.
[18]Arner P, Kulyté A. MicroRNA regulatory networks in human adipose tissue and obesity[J]. Nature Reviews Endocrinology, 2015, 11:276-288.
[19]Song XW, Li Q, Lin L, et al. MicroRNAs are dynamically regulated in hypertrophic hearts, and miR-199a is essential for the maintenance of cell size in cardiomyocytes[J]. Journal of Cellular Physiology, 2010, 225:437-443.
[20]Lee YB, Bantounas L, Lee DY, et al. Twist-1 regulates the miR-199a/214 cluster during development[J]. Nucl Acids Res, 2009. 37:123-128.
[21]Hashemi Gheinani A, Burkhard FC, Rehrauer H, et al. MicroRNA MiR-199a-5p regulates smooth muscle cell proliferation and morphology by targeting WNT2 signaling pathway[J]. J Biol Chem, 2015, 290(11):7067-7086.
[22]Chen R, Alvero AB, Silasi DA, et al. Regulation of IKKβ by miR-199a affects NF-κB activity in ovarian cancer cells[J]. Oncogene, 2008, 27:4712-4723.
[23]Beg A, Sha W, Bronson R, et al. Embryonic lethality and liver degeneration in mice lacking the RelA component of NF-κB[J]. Nature, 1995, 376:167-170.
[24]Doi TS, Marino MW, Takahashi T, et al. Absence of tumor necrosis factor rescues RelA-deficient mice from embryonic lethality[J]. PNAS, 1999, 96:2994-2999.
[25]Ryan KM, Ernst MK, Rice NR, et al. Role of NF-κB in p53-mediated programmed cell death[J]. Nature, 2000, 404(20):892-897.
[26]齐仁立, 黄金秀, 龙定彪, 等. MicroRNA与NF-κB调控的细胞凋亡[J]. 生物技术通报, 2015, 31(5):27-31.
(责任编辑 狄艳红)
Over-expression of MicroRNA-199a Intensifies the TNFα-induced Apoptosis of Adipocytes
QI Ren-li1,2WANG Qi1WU Yong-jiang3WANG Jing1HUANG Jin-xiu1,2YANG Fei-yun1,2
(1.Chongqing Academy of Animal Sciences,Chongqing 402460;2.Key Laboratory of Pig Industry Sciences,Ministry of Agriculture,Chongqing 402460;3.Southwest University Rongchang Campus,Chongqing 402460)
The aim of this study is to explore the effects of MicroRNA-199a(miR-199a)on adipocytes apoptosis and the role of nuclear factor κB(NF-κB). Tumor necrosis factor α(TNFα)was used to induce the apoptosis of cultured 3T3-L1 adipocytes. Based on this work,adding PDTC of NF-κB blocker and miR-199a mimic,the effect of overexpression of miR-199a on TNFα-induced apoptosis and the role of NF-κB in it were determined. The flow cytometry was used to analyze the cell apoptosis,kit Caspase3/7 to detect the enzymatic activities,dual luciferase reporter assay to detect the activity of NF-κB,qRT-PCR to detect the expression of miR-199a,and Western blotting to detect the changes of relevant proteins. As results showed,TNFα induced the apoptosis of differentiated adipocytes,concurrently activated NF-κB. The activated NF-κB played a negative regulator role in the apoptosis of adipocytes mediated by TNFα. Over-expressed miR-199a significantly suppressed the activation of NF-κB,and then remarkably promoted the apoptosis induced by TNFα. Conclusively,miR-199a involves in the TNFα-induced apoptosis of adipocytes through regulating NF-κB activation.
miR-199a;adipocyte;apoptosis;TNFα;NF-κB
10.13560/j.cnki.biotech.bull.1985.2017.03.026
2016-06-28
国家自然科学基金项目(31470117,31302055),重庆市畜牧科学院基本科研业务费项目(14403)
齐仁立,男,副研究员,研究方向:动物脂肪代谢的基因调控,E-mail:qirenli1982@163.com;王琪为共同第一作者
杨飞云,男,研究员,E-mail:yfeiyun@yeha.net
