猪IGF2基因多态性研究
2017-03-30武建亮卢纪和孟庆利马小军周海深王丽丽
武建亮,徐 利,卢纪和,孟庆利,马小军,周海深,王丽丽
(北京养猪育种中心,北京 100194)
猪IGF2基因多态性研究
武建亮,徐 利,卢纪和,孟庆利,马小军,周海深,王丽丽
(北京养猪育种中心,北京 100194)
该研究分析了猪胰岛素样生长因子2(IGF2)基因内含子3的3 072位点在大白、长白、杜洛克3个品种中的多态性,并分析不同基因型与100 kg体重日龄和背膘厚的关系。结果表明,在3个品种中均存在A、G基因型,且A基因型为优势基因型。3个品种中A基因型个体背膘厚显著低于G基因型个体(P<0.05),而不同基因型间100 kg体重日龄无显著差异。结果提示IGF2可作为脂肪沉积的候选基因应用于标记辅助选择。
猪;IGF2基因;多态性;背膘厚
胰岛素样生长因子2基因(Insulinlike Growth Factor-2,IGF2),是最早发现的内源性印迹基因,其表达、调控不遵循孟德尔遗传规律。IGF2是一种多功能细胞增殖调控因子,对细胞的分化、增殖、胚胎的生长发育具有重要作用。IGF2是一种促细胞分裂多肽,也被称为生长调节素A,参与生长激素发挥作用,刺激培养细胞生长,在哺乳动物的生长发育过程中起着重要作用。有研究表明,在欧洲野公猪与大白猪的杂交后代以及皮特兰与大白猪的杂交后代中存在一个影响猪肌肉生长和脂肪沉积的QTL,为父源表达,并定位在猪2号染色体短臂末端IGF2基因座上[1-4]。近来发现,IGF2基因内含子3的3 072位点的G→A单碱基突变正好位于高度保守的调控区域,影响猪骨胳中IGF2的表达,并且对出生后的肌肉形成起重要作用,该G→A突变与瘦肉率存在相关性,使得猪瘦肉量增加3%~4%[5]。许多研究结果发现,IGF2基因还与猪的初生重、生长速度、背膘厚、瘦肉率等重要经济性状相关[6-8]。本研究以大白、长白、杜洛克为研究对象,分析IGF2基因不同基因型与生长速度、背膘厚的关系,旨在为猪生长和肉质性状标记辅助选择提供理论依据。
1 材料与方法
1.1 实验材料
在北京养猪育种中心的下属猪场采集了大白、长白、杜洛克3个品种共600头母猪的耳组织,放入75%的酒精中于-20 ℃保存。同时收集猪只的背膘厚和100 kg体重日龄等生长性状相关指标数据。DNA提取试剂盒、PCR相关试剂、内切酶、Marker、琼脂糖、TAE缓冲液等购自TAKARA公司。
1.2 DNA提取
按照DNAiso Reagent试剂盒说明书提取DNA,-20 ℃保存。
1.3 PCR反应
根据GenBank中检索到的猪IGF2基因(序列号:AY242109)DNA序列,用Primer 3.0软件在IGF2 intron3 G3072A位点附近设计引物。IGF2引物序列:F-5’-CGGACCGAGCCAGGGACGA-3’;R-5’-GCCGGCTGGAAGGGAGGAA-3’。
IGF2基因PCR扩增采用的25 μL反应体系为:2×Taq PCR Mastermix 12.5 μL,引物各0.5 μL(10 μM),DNA 1.0 μL和水。PCR扩增的反应条件: 94 ℃变性4 min,35次循环(94 ℃,30 s;54.5 ℃,30 s;72 ℃,30 s),72 ℃延伸5 min。
1.4 多态性检测
PCR产物经琼脂糖凝胶电泳检测合格后直接测序,序列通过MEGA v4.0软件进行比对,确定检测个体IGF2的基因型。
1.5 数据分析
100 kg体重日龄和背膘厚指猪只在85~135 kg范围内实际称重并用ALOKA 500型B超仪测量的倒数第三、第四肋骨间的背膘厚,根据《全国种猪遗传评估方案(试行)》提供的校正公式校正后的100 kg体重日龄和背膘厚。
IGF2第三内含子3 072位点不同基因型对背膘厚和100 kg体重日龄的影响用SPSS 17.0软件中的一般线性模型(GLM)进行统计分析,GLM为:
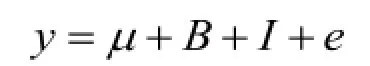
2 结果与分析
2.1IGF2基因多态性检测及基因型频率
根据序列比对结果,在所检测的个体中在IGF2基因第三内含子3 072位点存在A和G两种基因型。根据IGF2基因不同基因型在各品种中的检出个数,计算其基因型频率,结果见表1。

表1 IGF2第三内含子3 072位点不同基因型频率
2.2IGF2第三内含子3 072位点不同基因型与背膘厚的关系
IGF2第三内含子3 072位点不同基因型对背膘厚的影响见表2。IGF2第三内含子3 072位点不同基因型的背膘厚在大白、长白和杜洛克3个品种中均表现为G型显著高于A型(P<0.05)。就同一基因型而言,大白和长白、长白和杜洛克间均无显著差异,大白和杜洛克间差异显著(P<0.05)。

表2 IGF2第三内含子3 072位点不同基因型对背膘厚的影响 mm
2.3IGF2第三内含子3 072位点不同基因型与100 kg体重日龄的关系
IGF2第三内含子3 072位点不同基因型对100 kg体重日龄的影响见表3。IGF-2第三内含子3 072位点不同基因型的100 kg体重日龄在大白、长白和杜洛克3个品种中均无显著差异。就IGF2第三内含子3 072位点同一基因而言,大白和长白猪间的100 kg体重日龄差异不显著,杜洛克与其他两个品种的100 kg体重日龄差异显著(P<0.05)。

表3 IGF2第三内含子3 072位点不同基因型对100 kg体重日龄的影响 d
2.4 猪100 kg体重日龄和背膘厚的差异性分析
猪100 kg体重日龄和背膘厚在不同品种、IGF-2不同基因型之间存在一定的差异,该研究通过建立一般线性模型进行方差分析剖析了差异的来源,结果见表4。建立的一般线性模型对于猪不同品种、IGF-2不同基因型之间的100 kg体重日龄和背膘厚的分析是适宜的(P<0.01),其校正的R2统计量分别为0.012和0.131,品种差异显著(P<0.05),IGF2不同基因型对猪100 kg体重日龄没有显著效应,而对背膘厚有显著效应(P<0.05)。说明猪背膘厚的差异主要来源于不同基因型和品种的差异。

表4 猪100 kg体重日龄和背膘厚的差异性分析
3 讨论
3.1IGF2基因多态性分析
IGF2基因多态性研究已经成为猪育种实践应用中关注的焦点。Knoll等[9]通过连锁分析,确定了猪IGF2基因内含子2中有G→A转换的单核苷酸多态性,这一转换导致了NciⅠ限制性酶切位点的改变。有研究进一步分析了这一单核苷酸多态性在长白、大白、杜洛克猪和皮特兰4个品种中的分布情况[2,10]。刘桂兰等[11]对大白×梅山猪F2代资源家系群体的IGF2基因第8内含子的研究中,发现两个等位基因频率差异较大的NciⅠ酶切位点。薛慧良等[12]对猪IGF2基因外显子8进行了PCR-SSCP检测,发现单核苷酸C→T转换。有研究发现猪IGF2基因内含子3的3 072位点的存在G→A单碱基突变[5]。本研究在大白、长白和杜洛克3个品种中发现在第3内含子3 072位点均存在A和G两种基因型,且A等位基因为优势等位基因。这一结果与虞德兵等[13]、Yang等[14]分析这一多态性在长白、大白、长×大杂交猪及二花脸等中国地方品种中的分布情况相符,即A等位基因在长白和大白猪群体中为优势等位基因。
3.2IGF2基因多态性与100 kg体重日龄的关系
本研究结果表明IGF2第3内含子G3 072A多态性在大白、长白和杜洛克3个品种的100 kg体重日龄均无显著相关。而林长光等[7]研究表明大白和长白猪中该位点AA基因型个体比GA和GG基因型个体生长速度快,等位基因A对猪的生长具有正效应。Kolaríková等[10]对大白猪群体IGF2基因内含子2的研究结果表明,不同基因型对日增重无显著相关性。而薛慧良等[12]、Magri等[15]和Grrard[16]等的研究表明在IGF2基因外显子8上检测到的多态性位点基因型间初生重差异显著。
3.3IGF2基因多态性与背膘厚的关系
在诸多研究中均表明IGF2基因中不同位点的多态性与猪的背膘厚、瘦肉率等肉质性状存在相关性。如IGF2基因内含子8上NciI多态性位点的不同基因型间脂肪沉积和瘦肉量存在差异显著[11]。IGF2基因内含子7上多态性位点的基因型间背膘厚及瘦肉率有着显著差异显著[17]。IGF2基因外显子8上检测到的多态性位点不同基因型间6月龄背膘厚的差异显著[12]。IGF2基因内含子3的3 072位点G→A多态性与猪瘦肉率存在相关性,A型个体的瘦肉率比G型个体高3%~4%[5]。同时有研究表明这一变异对梅山×欧洲白猪杂交后代个体的背膘厚有显著影响[18]。相关研究还表明A基因型个体的在臀部、胸腰部和第6~7肋处的膘厚比G型个体要薄[13]。本研究分析表明IGF-2基因第3内含子G3 072A多态性与在大白、长白、杜洛克3个品种的背膘厚间存在显著差异,且A基因型个体背膘厚显著低于G基因型个体,这与Laere等、Jungerius等、虞德兵等研究结果相一致。表明在选种、育种实践过程中,IGF2基因可作为脂肪沉积的候选基因应用于标记辅助选择,选择带有A等位基因的个体有望降低后代的背膘厚、提高胴体瘦肉率,改善猪肉品质,加快育种进程。
[1] JEON J T,CARLBORG ,T RNSTEN A,et al.A paternally expressed QTL affecting skeletal and cardiac muscle mass in pigs maps to the IGF2 locus[J].Nature genetics,1999,21(2):157-158.
[2] NEZER C,MOREAU L,BROUWERS B,et al.An imprinted QTL with major effect on muscle mass and fat deposition maps to the IGF2 locus in pigs[J].Nature Genetics,1999,21(2):155—156.
[3] ANDERSSON L,HALEY CS,ELLEGREN H,et al.Genetic mapping of quantitative trait loci for growth and fatness in pigs[J].Science,1994,263(5154):1771—1774.
[4] ALEXANDER L J,TROYER D L,ROHRER G A,et al.Physical assignments of 68 porcine cosmid and lambda clones containing polymorphic microsatellites[J].Mammalian Genome,1996,7(5):368-372.
[5] V A N L A E R E A S,N G U Y E N M,BRAUNSCHWEIG M,et al.A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig[J].Nature,2003,425(6960):832—836.
[6] 孙亿,费思清,唐辉,等.猪IGF2基因G3072A位点多态性及其与大白猪初生重和早期生长的关系[J].畜牧兽医学报,2007,38(12):1306-1310.
[7] 林长光,刘亚轩,吴悌霖,等.IGF2基因多态性检测与生长性状相关性分析[J].福建农林大学学报(自然科学版),2011,40(6):624-627.
[8] STINCKENS A,MATHUR P,JANSSENS S,et al.Indirect effect of IGF2 intron3 g.3072G> A mutation on prolificacy in sows[J].Animal genetics,2010,41(5):493-498.
[9] KNOLL A,PUTNOVA L,DVO K J,et al.A Ncii PCR RFLP within intron 2 of the porcine insulin like growth factor 2b (IGF2) gene[J].Animal genetics,2000,31(2): 150.
[10] KOLA KOV O,PUTNOV L,URBAN T,et al.Associations of the IGF2 gene with growth and meat efficiency in Large White pigs[J].J.Appl.Genet,2003,4(44):509-513.
[11] 刘桂兰,蒋思文,熊远著,等.IGF2基因PCR-RFLP多态性与脂肪沉积相关性状的关联分析[J].遗传学报,2003,30(12):1107—1112.
[12] 薛慧良,徐来祥,XUEHui-Liang,等.猪IGF2基因的遗传多态性及其遗传效应分析[J].遗传,2008,30(2):179-184.
[13] 虞德兵,何宗亮,张伟峰,等.猪IGF2基因内含子3变异的遗传效应分析[J].遗传,2008,30(1):87-93.
[14] YANG G C,REN J,GUO Y M,et al.Genetic evidence for the origin of an IGF2 quantitative trait nucleotide in Chinese pigs[J].Animal genetics,2006,37(2): 179-180.
[15] MAGRI K A,BENEDICT M R,EWTON D Z,et al.Negative feedback regulation of insulin-like growth factor-II gene expression in differentiating myoblasts in vitro[J].Endocrinology,1994,135(1):53-62.
[16] Gerrard D E,Okamura C S,Ranalletta M A,et al.Developmental expression and location of IGF-I and IGFII mRNA and protein in skeletal muscle[J].Journal of Animal Science,1998,76(4):1004-1011.
[17] VYKOUKALOVA Z,KNOLL A,DVO K J,et al.New SNPs in the IGF2 gene and association between this gene and backfat thickness and lean meat content in Large White pigs[J].Journal of Animal Breeding and Genetics,2006,123(3):204-207.
[18] JUNGERIUS B J,VAN LAERE A S,TE PAS M F W,et al.The IGF2-intron3-G3072A substitution explains a major imprinted QTL effect on backfat thickness in a Meishan×European white pig intercross[J].Genetical research,2004,84(2):95-101.
2016-12-26)
北京三元种业科技股份有限公司自立课题,课题编号:SYZYZ20130015
武建亮,男,博士,从事种猪生产与管理工作,E-mail:wwwjl1617@163.com
