重庆綦江白垩系夹关组遗迹化石及其沉积环境
2017-03-28胡旭峰王荀仟张海萍周羽漩王丰平谢显明
熊 璨,代 辉,陈 昀,胡旭峰,王荀仟,张海萍,周羽漩,王丰平,谢显明
(1.重庆市地质矿产勘查开发局208水文地质工程地质队,重庆 400700;2.重庆市云阳县文物保护管理所,重庆 404500;3.重庆市綦江区国土资源和房屋管理局,重庆 401420)
引言
遗迹化石是指地质历史时期的生物在沉积物及其他底质表面或者内部产生的各种生命活动记录,遗迹化石种类繁多,包括潜穴、爬痕、脊椎动物足迹、粪化石、卵化石以及一些微生物成因的沉积构造[1-2]。遗迹化石通常为未经搬运地原地埋藏,由于埋藏环境的差异,在这些地层中往往又缺乏实体化石,这就造就了遗迹化石在古环境、古气候和古生态等方面研究的重要性。不仅如此,遗迹化石在能源和其他沉积矿产资源的研究与勘探方面也有着独特的应用价值[3-6]。最早的遗迹化石研究主要是脊椎动物足迹的研究,可以追溯到19世纪初[7]。发展至今,遗迹化石的研究已经在众多分支领域取得了长足的进展[8-9]。龚一鸣等(2009)将中国的遗迹化石研究分为3个阶段:零星阶段(1929-1978)、系统阶段(1978-2004)、创新阶段(2004 至今)[5]。
重庆綦江位于四川盆地的东南缘与华蓥山帚状褶皱束山脉的南倾伏部,地处黔北山区和四川盆地过渡区。2007年,邢立达等对该研究区的恐龙足迹进行了详细调查描述,命名了甲龙类恐龙足迹中国綦江足迹(Qijiangpus sinensis)、兽脚类恐龙足迹敏捷舞足迹(Wupus agilisichnogen)、鸟脚类恐龙足迹炎热老瀛山足迹(Laoyingshanpus torridusichnogen)和莲花卡利尔足迹(Caririchnium lotus)[10]。该地区的古脊椎动物足迹见有多次补充报道,包括翼龙足迹、古水鸟足迹(2007年被描述为兽脚类恐龙足迹)和蜥脚类恐龙足迹幻迹等[10-12]。本文研究区无脊椎动物的遗迹化石尚未见报道。
本文描述了重庆綦江国家地质公园老瀛山园区莲花保寨中白垩系夹关组中的遗迹化石,初步探讨了遗迹化石的组成特征及其沉积环境,为阐明该地区夹关组沉积演化提供了新的遗迹化石证据。
1 研究区位置及地质背景
重庆市綦江国家地质公园位于綦江区境北部,园区跨古南镇、三角镇、石角镇、三江镇和永新镇的部分地域。本文的研究区位于三角镇红岩村陈家湾后山的莲花保寨,地理坐标为东经106°23′至106°43 ′,北纬28°26 ′至29°1 ′(图1)。本文的遗迹化石产于莲花保寨凹岩腔中的白垩系夹关组地层中。该段地层下部为灰黄色厚层状砾岩,向上逐渐变为含砾砂岩,中部为紫红色石英砂岩与泥质粉砂岩互层,顶部为砖红色块状含砾长石石英砂岩,遗迹化石产于泥质粉砂岩底部和石英砂岩顶部(图2)。
夹关组属于下白垩统还是上白垩统目前尚有争议。1972年,四川二区域地质测量队将夹关组和下灌口组划分到下白垩统,将上灌口组划分到上白垩统;1977年,四川省航空区域地质调查队直接将夹关组划分到上白垩统,而灌口组划分到了下白垩统;1983年,李玉文等通过对四川嘉定群介形类进行研究,将夹关组分为下中上部,其中夹关组下部划入到下白垩统,中部划入到中白垩统,上部划分到上白垩统;1995年,李元林对四川盆地大溪砾岩夹关组碎屑中的石英颗粒采取ESR(电子自旋共振)方法,测得夹关组年龄位于117Ma到85Ma(Aptian-Santonian)之间[13];2001 年,苟宗海和赵兵对四川大邑、崇州地区的夹关组同样采取ESR方法,测得夹关组年龄位于 129Ma到 91Ma(Valanginian-Santonian)之间[14];2009 年,陈海霞在川西雅安市夹关组剖面中采取了丰富的孢粉化石,通过研究建立孢粉组合带,将夹关组地层划分到下白垩统Barremian-Albian,也就是145Ma到100.5Ma之间[15]。为避免争议,本文将夹关组仅划分到白垩系。

图1 研究区地理位置图Fig.1 Geographical location of the study area
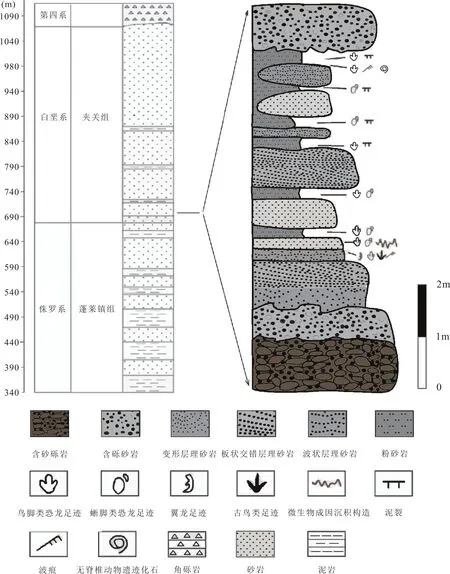
图2 研究区地层柱状图Fig.2 Stratigraphic column through the Jiaguan Formation in Qijiang,Chongqing
2 遗迹化石鉴定与描述
按遗迹化石结构构造和造迹生物的行为习性分类,经仔细辨认和鉴定,綦江国家地质公园老瀛山园区夹关组地层中已发现的无脊椎动物遗迹化石经鉴定包括1大类3属3种:Scoyenia gracilis(纤细斯柯茵迹)、Beaconites antarcticus(南极似塔形迹)和Planolites beverleyensis(贝弗利漫游迹),全为觅食迹。现将上述3种遗迹化石的详细特征描述如下:
遗迹属ScoyeniaWhite,1929
遗迹种Scoyenia gracilisWhite,1929
描述:潜穴为单一平直-微弯曲形底生迹(hypichnia),呈管状,直径13mm,粗细较均一,延伸长度约14cm,平行层面分布;潜穴表面可见不显著的纵向或斜向纹饰,潜穴内填充有半月形回填构造,填充物颜色与围岩相同,均为肉红色细砂岩,但填充物颗粒粒径较围岩粗(图3箭头所指)。
讨 论:KeckiaGlocker,LaminitesGhentet Henderson,MuenstreiaVonSternberg,PlagiogmusRoedel,TaenidiumHeer,和NeoneritesSeilacher 等都具有半月形充填构造,它们与Scoyenia的区别是均无外表上的纵向纹饰。根据该遗迹化石的直径大小、形态及内部的半月形回填构造和表面纵向纹饰在此将标本归为Scoyenia gracilis。
Frey和Pemberton(1984)根据潜穴外表的条纹推测Scoyenia gracilis的造迹动物不是Polychaete(多毛类)就是节肢动物,或者是寡毛类(主要是非海相环节动物),考虑到淡水多毛类潜穴稀少,其潜穴内部构造所知甚少,因此倾向于造迹动物可能是节肢动物[16]。根据潜穴内部的回填构造可以推测Scoyenia应当是觅食构造(fodinichnia)。
产地及层位:重庆市綦江区白垩系夹关组

图3 遗迹化石Scoyenia gracilis White,1929(比例尺为5cm)Fig.3 The trace fossil Scoyenia gracilis White,1929 (scale:5 cm)
遗迹属BeaconitesVialov,1962
遗迹种Beaconites antarcticusVialov,1962
描述:平直-微弯曲或弯曲的管状潜穴,不分枝,但可见相互交叉穿越或截断,整体有呈放射状向外伸展;潜穴长短不一,最长可达22cm,潜穴直径一般为8-13mm,但个体较小者也有4mm左右;潜穴具有薄的、清晰的衬壁,可见清晰的塔形或“U”形回填构造,回填纹宽度明显宽于潜穴直径;半浮痕或内迹保存,平行或倾斜于层面分布(图4)。
讨论:以往研究对具有新月形回填构造的潜穴,如Beaconites、Ancorichnus、Taenidium,一直存在着争议,争议在于回填构造的形态、潜穴壁的有无及其表面特征。Bromley(1996)认为Ancorichnus中心带具有“U”形回填纹并与侧壁回填纹的方向相反[17]。Keighley 和Pickerill(1994)对 遗 迹 属Beaconites进行了修订,将是否具有较厚的衬壁作为区分Beaconites和Taenidium的标志[18]。然而,很多学者也对这一提法提出质疑,主要是Vialov(1962)对Beaconites的描述并没有提到有清晰的衬壁[19],Retallack(1997)认为新月形回填构造就是鉴别他们的有效标志[20]。本文采用 Keighley和 Pickerill(1994)的区分方法。
Beaconites遗迹属的三个遗迹种:Beaconites antarcticus,Beaconites capronus和Beaconites coronus,也有其区分方法。Beaconites capronus回填构造呈“V”形;Beaconites antarcticus和Beaconites coronus回填构造均为“U”形,但前者回填纹的单个房室宽度大于潜穴直径,后者回填纹的单个房室宽度小于潜穴直径。这里遗迹化石为“U”形回填构造,且单个房室宽度大于潜穴直径,故归为Beaconites antarcticus。
产地及层位:重庆市綦江区白垩系夹关组

图4 遗迹化石Beaconites antarcticus Vialov,1962(比例尺为5cm)Fig.4 The trace fossil Beconites antarcticus Vialov,1962(scale:5 cm)
遗迹属PlanolitesNicholson,1873
遗迹种Planolites beverleyensisBilling,1862
描述:平直或微弯曲或弯曲管状潜穴,潜穴不分枝,但常互相穿插排列;潜穴长3~15cm,直径一般为8~9mm,但个体较小者也有5mm左右;潜穴表面光滑,无衬壁;潜穴部分有充填,充填物为浅灰色细砂岩,颜色与围岩不同。半浮痕或内迹保存,与层面平行或略微斜交(图5箭头所指)。
讨论:遗迹属Macaronichnus和Palaeophycus与Planolites形态特征相近。但Macaronichnus仅产自海滩较纯净的细砂沉积物中,潜穴壁较薄,而Palaeophycus发育在多种沉积环境中,潜穴壁较厚,Planolites则出现在多种较平静的沉积环境中,但一般无衬壁[21-22]。
依据潜穴的大小、弯曲度和潜穴纹饰,Pemberton和Frey(1982)把真正能代表Planolites的13个种归结为3个遗迹种,即Planolites montanus,Planolites beverleyensis和Planolites annularis[23]。其中,前两者可以根据潜穴直径加以区分,Planolites montanus的直径很少超过5mm,弯曲成蛇曲状;Planolites beverleyensis直径一般不小于8mm,潜穴直或微弯曲;Planolites annularis有明显的横纹或环状纹饰。研究区遗迹化石潜穴直径8~9mm,无环状纹饰,所以将其归为Planolites beverleyensis。
产地及层位:重庆市綦江区上白垩统夹关组
3 遗迹相及其沉积环境
生物群落的分布和发展被诸多因素所控制。对于海洋、湖泊和河流等水生底栖生物而言,水的深度、能量、含氧量、基底的性质(成分、粘结度等)和沉积速度等都对其有着很大的影响,这些影响因素是相互关联的,影响着生物群落的分异度、生物个体大小,以及生物活动量等,其变化通过造迹生物就能反映到遗迹化石方面[1]。
目前研究认为,Scoyenia遗迹相包括:Scoyenia、Beaconites、Ancorichnus、Taenidium、Planolites、Palaeophycus、Ancorichnus、Cruziana、Rusophycus、Diplichnites和Umfolozia等遗迹属[1,24]。本文遗迹化石均为近层面觅食迹,包含Scoyenia gracilis,Beaconites antarcticus和Planolites beverleyensis,故属于Scoyenia遗迹相。Bromley和Asgaard(1979)研究格陵兰东部三叠纪红层时,提出了Scoyenia和Rusophycus遗迹相代表了周期性泛滥的河流沉积环境[25]。Frey 等 (1984)专门研究了Scoyenia和Ancorichnus遗迹属,认为Scoyeniagracilis和Ancorichnus coronus是泛滥平原沉积中的特征遗迹化石[16]。胡斌和吴贤涛对四川峨眉夹关组遗迹化石群落进行了研究,提到Scoyenia遗迹化石组合代表了一种经常干旱的和周期性泛滥的河流环境(包括洪泛平原上蓄水极浅的小型湖泊环境)中的生态类型[26]。Spencer等(2011)对美国德克萨斯州中北部早二叠世晚期陆相遗迹群落研究时,提出Scoyenia遗迹相和Batrachichnus遗迹相代表了河漫滩和洪泛平原上短暂的浅水湖环境[27]。Hu等(2014)在总结中国陆相遗迹相和遗迹组合时,提到Scoyenia遗迹相发育在浅水到地表的低能过渡带,特别是河流的洪泛平原和湖滨地带,该地段间断的暴露于空气中,Scoyenia遗迹相常伴有泥裂和雨痕出现[28]。
从上述结论来看,本文Scoyenia遗迹相代表了:(1)该段地层沉积时水深较浅、水体能量低,沉积面间断性暴露地表;(2)造迹生物的活动区域为水道附近的漫滩或者洪泛平原上的浅水洼地环境。
4 结论
研究区发现的无脊椎动物遗迹化石均属觅食迹,包括3属 3种:Scoyenia gracilis(纤细斯柯茵迹)、Beconites antarcticus(南极似塔形迹)和Planolites beverleyensis(贝弗利漫游迹)。这些遗迹化石均为底生迹,平行于层面分布,少量与层面斜交。研究区已发现的遗迹化石均属于Scoyenia遗迹相,代表了间断性暴露地表的漫滩或者洪泛平原上的浅水洼地环境。
重庆綦江莲花保寨白垩系夹关组地层中的古脊椎动物足迹化石(恐龙足迹、翼龙足迹和古鸟类足迹)已经多次被报道。本文对研究区的无脊椎动物遗迹化石的首次报道丰富了该研究区的遗迹化石种类,为揭露该地区的古地理、古环境和古生态等提供了更加丰富的信息。
致谢:河南理工大学的胡斌教授和韩国遗迹化石专家Jeong Yul Kim对本文遗迹化石的鉴定和描述提供了宝贵的意见,重庆市綦江区国土房管局在野外工作期间提供了帮助,在此一并表示衷心的感谢。
参考文献:
[1] 杨式溥,张建平,杨美芳.中国遗迹化石[M].北京:科学出版社,2004.1-17.
[2] 童金南,殷鸿福.古生物学[M].北京:高等教育出版社,2007.16-17.
[3] Soren J,Mary L D,James G G.Trace fossil preservation and the earlyevolutionofanimals[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2005,220(1 -2):19 -29.
[4] 施振生,杨威,郭长敏,等.遗迹化石在层序地层学研究中的应用[J]. 地层学杂志,2008,32(1):79 -84.
[5] 龚一鸣,胡斌,卢宗盛,等.中国遗迹化石研究80年[J].古生物学报,2009,48(3):322 -337.
[6] Richard H,Nicolas G,Martin W,et al.New trace fossil evidence for an early recovery signal in the aftermath of the end-Permian massextinction[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2011,310(3 -4):216 -226.
[7] Cadee C C ,Goldring R.The Wadden Sea ,cradle of invertebrate ichnology[A].Miller W III.Trace Fossils:Concepts,Problems and Prospects[C].Amsterdam:Elsevier.2007.3-13.
[8] Miller W I.Trace fossils :concepts,problems,prospects[J].Ichnos-an International Journal for Plant& AnimalTraces,2009,16(4):286 -288.
[9] Seilacher A.Bathymetry of trace fossils[M].Marine Geology,2007,5:413 -428.
[10] 邢立达,王丰平,潘世刚,等.重庆綦江中白垩统夹关组恐龙足迹群的发现及其意义[J].地质学报,2007,81(11):1591-1602.
[11] Xing L D,Lockley M G,Wang F P,et al.Stone flowers explained as dinosaur undertracks:unusual ichnites from the Lower Cretaceous Jiaguan Formation,Qijiang District,Chongqing,China[J].Geological Bulletin of China,2015,34(5):885-890.
[12] Xing L D,Lisa G B,Richard T M,et al.Reanalysis of Wupus agilis (Early Cretaceous)of Chongqing,China as a Large Avian Trace:Differentiating between Large Bird and Small Non-Avian Theropod Tracks[J].PLOS ONE,2015,10(5):e0124039.
[13] 李元林.大溪砾岩及其年代归属[J].成都理工学院学报,1995,22(2):11 -14.
[14] 苟宗海,赵兵.四川大邑-崇州地区的白垩、第三系[J].地层学杂志,2001,25(1):28 -33.
[15] 陈海霞.川西雅安地区白垩纪古环境古气候研究 [D].成都:成都理工大学,2009.
[16] Frey R W,Pemberton S G,Fagerstrim J A.Morphologicalethological and environmental significance of the ichnogeneraScoyeniaandAncorichnus[J].Journal of Paleontology,1984,58:511 -528.
[17] Bromley R G.Trace fossils-biology,taphonomy and applications[M].London:Chapman & Hall,1996.
[18] Keighley D G,Pickerill R K.The ichnogenusBeaconitesand its distinction fromAncorichnusandTaenidium[J].Palaeontology,1994,37:305-337.
[19] Vialov O S.Problematica of the Beacon Sandstone at Beacon Heights West,Antarctica [J].New Zealand Journal of Geology and Geophysics,1962(5):718 -752.
[20] Retallack G J.Compaction of Devonian lycopsid stems from the Beacon Heights Orthoquatzite,southern Victoria Land [J].United States Antarctic Journal,1997,30(5):42 -44.
[21] 施振生,朱筱敏,王贵文等.塔里木盆地塔中地区志留系塔塔埃尔塔格组潮坪沉积中的遗迹化石[J].沉积学报,2005,23(1):91 -99.
[22] 胡斌,姜在兴,齐永安等.山东济阳坳陷古近系沙河街组深水湖沉积中的遗迹化石[J]. 古生物学报,2006,45(1):83-94.
[23] Pemberton S G,Frey R W.Trace fossil nomenclature and thePlanolites-Palaeophycusdilemma[J].Journal of Paleontology,1982,56:843-871.
[24] 吕希学,胡斌,姜在兴等.济阳济阳坳陷车镇和沾化凹陷古近系沙河街组遗迹群落及其沉积环境[J].古地理学报,2003,5(2):187 -194.
[25] Bromley R G,Asgaard U.Triassic freshwater ichnocoenoses from CarlsbergFjord, EastGreenland[J].Paleogeography,Paleoecology,Paleoclimatology,1979,28:39-80.
[26] 胡斌,吴贤涛.川西峨眉晚白垩世夹关期河流沉积中的痕迹化石群落[J]. 古生物学报,1993,32(4):478 -489.
[27] Spencer G L,Sebanstian V,Allan J L,et al.Late Early Permian continental ichnofauna from Lake Kemp,north-central Texas, USA[J].Palaeogeography, Palaeoclimatology,Palaeoecology,2011,308:395-404.
[28] Hu B,Wang Y Y,Song H B.The ichnofacies and ichnoassemblages in terrestrial deposits of China[J].Journal of Palaeogeography,2014,3(1):61 -73.
