不同形态锑对水稻生长及根系形态特征的影响研究
2017-03-16雷蕾吴倩桦吕海琴王瑞刚丁永祯徐应明范稚莲莫良玉冯人伟
雷蕾,吴倩桦,吕海琴,王瑞刚,丁永祯,徐应明,范稚莲,莫良玉,冯人伟,4,*
(1.农业部环境保护科研监测所,天津300191;2.中国农科院农田重金属污染修复创新团队,天津 300191;3.广西大学农学院,南宁530004;4.华中农业大学资源与环境学院,武汉 430070)
不同形态锑对水稻生长及根系形态特征的影响研究
雷蕾1,2,3,吴倩桦1,2,3,吕海琴1,2,4,王瑞刚1,2,丁永祯1,徐应明1,2,范稚莲3,莫良玉3,冯人伟1,2,3,4,*
(1.农业部环境保护科研监测所,天津300191;2.中国农科院农田重金属污染修复创新团队,天津 300191;3.广西大学农学院,南宁530004;4.华中农业大学资源与环境学院,武汉 430070)
通过溶液培养试验(采用埃斯皮诺营养液),研究不同形态锑[Sb(Ⅲ)和Sb(Ⅴ)]对常规籼稻(丰美占)生长、根系形态以及不同部位Sb含量的影响。结果表明:Sb(Ⅲ)和Sb(Ⅴ)均对水稻生长有抑制作用,且Sb(Ⅲ)的毒性较Sb(Ⅴ)高。随Sb(Ⅲ)和Sb(Ⅴ)处理浓度增加,地上部分和根系Sb含量均显著增加,且大量集中在根系。水稻对Sb(Ⅲ)的转运能力较Sb(Ⅴ)强,导致Sb(Ⅲ)处理下水稻地上部分Sb含量高于Sb(Ⅴ)处理,根系Sb含量则相反。Sb(Ⅲ)的加入显著抑制大多数根系形态参数,包括根长、根面积、根体积、根尖数以及根分叉数;而仅20 mg·L-1Sb(Ⅴ)能显著抑制水稻根面积、根直径、根分叉数。上述结果表明:Sb(Ⅲ)和Sb(Ⅴ)处理下,水稻能通过降低根面积和分叉数降低对Sb的吸收。随着Sb(Ⅲ)处理浓度增加,水稻细根比例从52.1%降低至41.5%,中等根比例从45.2%增加至55.2%,表明Sb(Ⅲ)处理下水稻能通过增加根系粗度来降低对Sb的吸收;而随着Sb(Ⅴ)处理浓度增加,水稻细根比例在52.1%~56.8%之间波动,中等根比例在41.7%~45.2%之间波动。
不同形态锑;水稻;毒性;根系形态;吸收
中国是全球最主要的锑(Sb)储量及生产大国[1],其Sb矿资源主要分布于湖南、贵州、广西、云南等南方省区[2]。由于Sb矿开采及含Sb化合物的广泛使用,我国环境中Sb的污染问题越来越严重。齐翠翠[3]对湖南锡矿山Sb矿区的调查发现,矿区表层土壤中Sb浓度高达5949 mg·kg-1,远超我国土壤Sb背景值(0.38~2.98 mg·kg-1)[4];He等[5]研究发现,锡矿山冶炼厂周边生长的水稻(Oryza sativa L.)体内富集了大量Sb,水稻根部、茎叶部及籽粒中Sb含量分别为225.34、18.78 mg·kg-1和5.79 mg·kg-1。环境中高含量的Sb能通过食物链进入人体,进而对人体产生毒害作用。刘碧君等[6]调查发现,锡矿山地区居民头发中Sb含量高达15.9 mg·kg-1。过量Sb会引发人体多种疾病,如:皮肤病、呼吸系统疾病、肺部疾病、心脑血管疾病、神经疾病以及癌症[7]。
Sb不是植物必需营养元素,但植物能大量吸收Sb[8]。目前有关动物、微生物等生物体内Sb的吸收、代谢及毒性研究已有所开展,但关于植物对Sb吸收、代谢及毒害作用的机理研究尚显不足。Sb对植物的毒害作用主要表现在以下几方面:(1)影响植物的生长发育,降低作物产量[9];(2)抑制植物根系生长[10];(3)诱导植物体内产生氧化胁迫[11];(4)影响植物对必需营养元素的吸收[12]。
根系是植物吸收矿质营养和水分的主要器官。根系形态的变化受植物遗传特性和环境因子的共同影响,其又能直接影响作物的生长及产量[13]。在重(类)金属胁迫下,植物能通过改变根系形态特征来降低对重(类)金属的吸收富集,从而降低重(类)金属对植物的毒性[14]。因此,在重(类)金属胁迫下,研究植物根系的形态变化能在一定程度上阐明重(类)金属的胁迫程度,以及植物对重(类)金属的解毒机制。但是,现有与植物根系形态相关的研究仅见于三价锑[Sb(Ⅲ)],而关于五价锑[Sb(Ⅴ)]对植物根系形态特征影响的研究较少[10]。基于以上原因,本研究以水稻为供试植物,利用水培实验,比较研究不同形态Sb对水稻生长及根系形态特征的影响,试图阐明不同形态Sb对水稻毒性的差异。
1 材料与方法
1.1 试验幼苗的培育及处理过程
本研究以常规籼稻(丰美占,由广东省农业科学院水稻研究所提供)为研究对象。水稻幼苗的培育过程如下:挑选颗粒饱满的种子于2%的NaClO溶液消毒20 min。去离子水清洗干净后,播种于润湿的培养基质(珍珠岩∶蛭石=1∶1,体积比)中。待水稻幼苗长出两片真叶,将其移至20 L塑料盆中适应3周,采用埃斯皮诺营养液培养,此营养液组分构成为:Ca(NO3)2· 4H2O 0.089 g·L-1、MgSO4·2H2O 0.250 g·L-1、(NH4)2SO40.049 g·L-1、KH2PO40.034 g·L-1、FeCl20.030 g·L-1、H3BO22.86 mg·L-1、ZnSO4·7H2O 0.22 mg·L-1、MnCl2· 2H2O 1.81 mg·L-1、CuSO4·5H2O 0.08 mg·L-1、H2MoO4· 4H2O 0.02 mg·L-1。
适应3周后,选取长势一致的水稻幼苗,去离子水清洗幼苗根系,移至1 L避光塑料桶中(1株·桶-1,埃斯皮诺营养液培养),同时加入不同浓度Sb处理。Sb(Ⅲ)和Sb(Ⅴ)溶液分别由分析纯的酒石酸锑钾和焦锑酸钾配制而成。试验分为7个处理,分别为:对照(CK,0 mg·L-1),5 mg·L-1Sb(Ⅲ),10 mg·L-1Sb(Ⅲ),20 mg·L-1Sb(Ⅲ),5 mg·L-1Sb(Ⅴ),10 mg·L-1Sb(Ⅴ),20 mg·L-1Sb(Ⅴ),每个处理3次重复。培养期间,每3 d换1次营养液,期间用0.1 mol·L-1的NaOH和HNO3调节pH为5.5。光照时间设置为16 h/ 8 h(昼/夜),温度为20℃/25℃(黑夜/白天),光照强度为100 μmol·m-2·s-1,相对湿度为60%~70%。Sb处理2周后收获植物。
1.2 样品的分析
收获时,将水稻分为根系和地上部,用滤纸将根系及地上部表面水分吸干后称量鲜重。采用爱普生平板式扫描仪(Epson Expression 10000XL 1.0)分析水稻根系形态,该扫描仪配备有根系图像分析系统(Win-RHIZO)。根系扫描获取如下参数:根长、根体积、根面积、平均根直径、根尖数和分叉数。根系参数获取后,将地上部分和根系样品剪碎分别混匀后在70℃的烘箱中烘干至恒重(48 h)。
根系、地上部分采用莱伯泰科DigiBlock ED54(北京莱伯泰科仪器股份有限公司)电热消解仪消解。过程如下:称取植物样品约0.2 g至消煮管中,加入10 mL浓硝酸,静置一夜。消煮程序为:先以80℃加热1.5 h,再升温至120℃加热1.5 h,最后以150℃加热3 h,冷却后揭盖,温度调至180℃,蒸干管内液体至约1 mL,冷却、用超纯水定容至50 mL容量瓶中。消解后样品的Sb含量用赛默飞电感耦合等离子体质谱测定。
1.3 数据处理及分析
采用单因素并结合多重比较法(Tukey检验)比较各处理间的显著性(P<0.05)。本试验中如不另外说明,所有的数据均为3次重复的平均值。数据统计分析采用SPSS 18.0软件,并用Sigmaplot 10.0软件作图。
2 结果与分析
2.1 不同形态Sb对水稻幼苗生物量的影响
由图1可以看出,与对照相比,除10mg·L-1Sb(Ⅴ)处理未对地上部生物量和5 mg·L-1Sb(Ⅲ)未对根系生物量有显著影响外,其他含Sb处理均显著降低了水稻地上部和根系生物量。同一处理浓度下,Sb(Ⅲ)处理水稻地上部生物量明显低于Sb(Ⅴ)处理,例如:5 mg·L-1处理时,Sb(Ⅴ)处理下地上部分生物量为Sb(Ⅲ)处理下的2.4倍;Sb(Ⅲ)处理下的根系生物量为Sb(Ⅴ)处理下的1.7倍。而在10、20 mg·L-1处理时,Sb(Ⅲ)处理下的根系生物量与Sb(Ⅴ)处理下的根系生物量相似。
2.2 不同形态Sb对水稻根系和地上部分Sb含量的影响
如图2所示,水稻地上部和根系Sb含量均随Sb处理浓度的增加而显著增加,且大部分Sb被富集在水稻根系,向地上部转移较少[Sb(Ⅲ)处理下Sb的转运系数在0.23~0.13之间,Sb(Ⅴ)处理下Sb的转运系数在0.05~0.06之间,数据未列出]。同一处理浓度下,Sb(Ⅲ)处理水稻地上部分Sb含量均高于Sb(Ⅴ)处理,例如:在10 mg·L-1处理下,Sb(Ⅲ)处理水稻地上部分Sb含量为Sb(Ⅴ)处理的3.1倍(数据未显示)。与地上部分Sb含量相反,Sb(Ⅴ)处理水稻根系Sb的含量均高于Sb(Ⅲ)处理。
2.3 不同形态Sb对水稻根系形态特征的影响
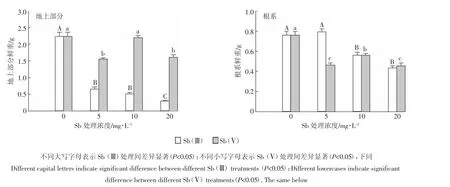
图1 不同形态Sb对水稻地上部分和根系生物量的影响Figure 1 Effects of different forms of Sb on the shoot and root biomass of rice plants
如表1所示,不同形态Sb均对大多数水稻根系形态特征指标产生不利影响。与对照相比,20 mg·L-1Sb(Ⅲ)处理显著降低水稻根长、根面积、根体积、根尖数、分叉数,分别达81.9%、70.0%、59.6%、81.9%和80.4%。但Sb(Ⅲ)处理未显著影响水稻根直径。Sb(Ⅴ)处理对水稻根系形态特征相关参数的影响较Sb(Ⅲ)处理的小。与对照相比,≤10 mg·L-1Sb(Ⅴ)处理仅显著降低了水稻根直径,未对其他指标有显著影响。添加20 mg·L-1Sb(Ⅴ)显著降低了水稻根面积、根直径和分叉数,分别达20.1%、22.2%和40.6%,未显著影响水稻根长、根体积和根尖数。

图2 不同形态Sb处理下水稻地上部分和根系的Sb含量Figure 2 Concentration of Sb in the shoots and roots of rice plants subjected to different forms of Sb

表1 不同形态Sb对水稻根长、根面积、根体积、平均根直径、根尖数和分叉数的影响Table 1 Effects of different forms of Sb on the length,area,volume,average diameter,tip number and fork number of rice roots
在处理浓度为10、20 mg·L-1时,Sb(Ⅴ)处理的根长、根面积、根体积、根尖数以及根分叉数均要高于Sb(Ⅲ)处理的。如10 mg·L-1处理下,Sb(Ⅴ)处理水稻根长、根面积、根体积、根尖数及根分叉数分别为Sb(Ⅲ)处理的1.96、2.22、1.59、2.43倍和2.33倍。
2.4 不同形态Sb对水稻根系组成比例的影响
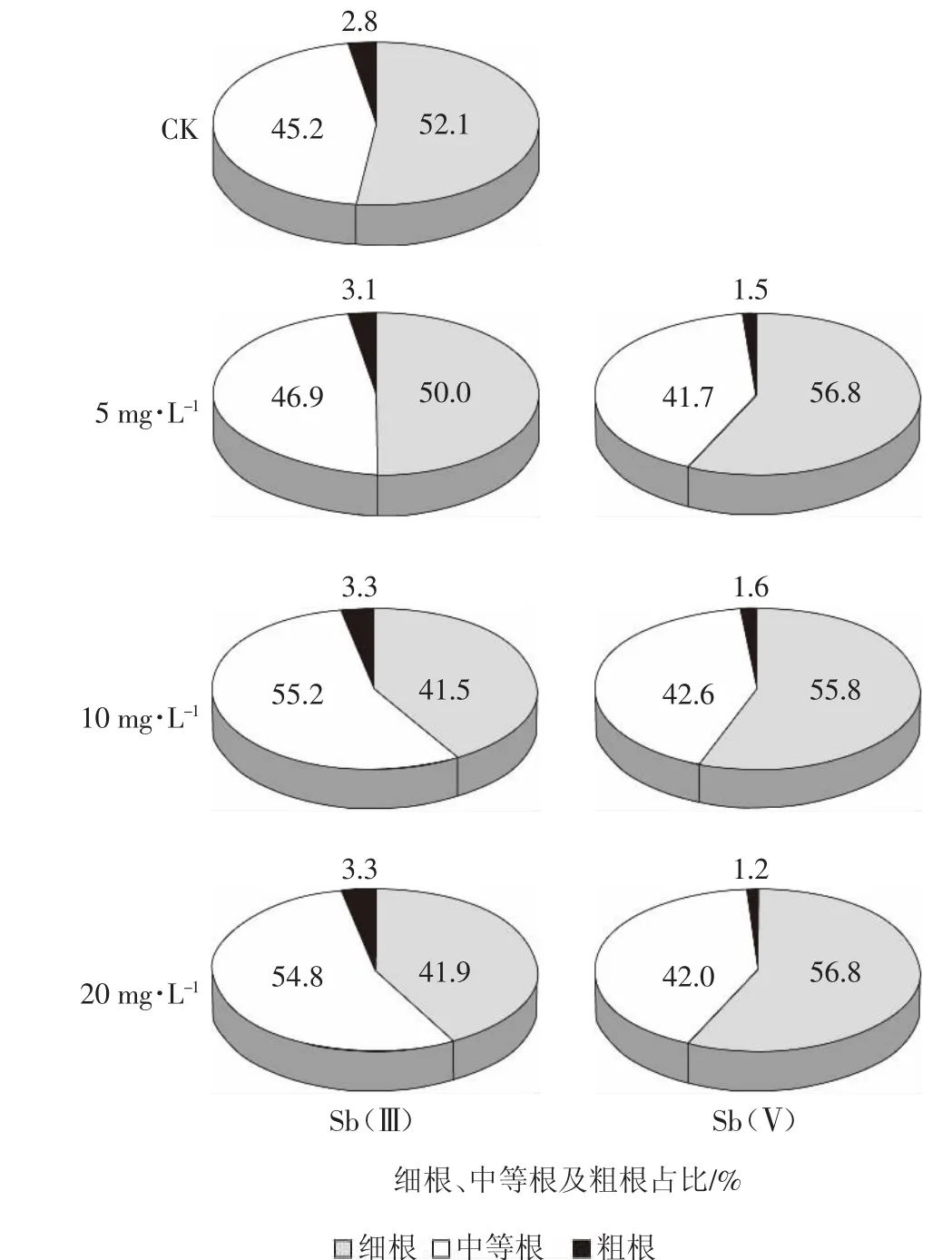
如图3所示,与对照相比,随Sb(Ⅲ)处理浓度增加,粗根比例从2.8%增至3.3%,中等根比例从45.2%增至54.8%,而细根比例从52.1%降至41.9%。随Sb(Ⅴ)处理浓度增加,粗根比例从2.8%降至1.2%,中等根比例从45.2%降至42%,而细根比例从52.1%增至56.8%。
3 讨论
研究表明,Sb在土壤中的植物有效性较低,但植物仍能大量吸收Sb,从而影响其生长[15]。本研究中,虽然两种形态Sb均显著抑制了水稻地上部分和根系生物量,但是不同形态Sb对水稻的毒性作用存在显著差异。例如:Sb(Ⅲ)处理下水稻地上部分生物量显著低于Sb(Ⅴ)处理下的,表明Sb(Ⅲ)对水稻的毒性较Sb(Ⅴ)高,此结果与前人报道结果相似[16]。
本研究中,水稻吸收的Sb大部分富集在其根系,向地上部转移较少,这一结果与Ren等[17]研究结果相似。但值得注意的是:本研究中Sb(Ⅲ)处理水稻地上部Sb含量高于Sb(Ⅴ)处理,而根系则相反,表明丰美占这一水稻品种对Sb(Ⅲ)的转运能力较Sb(Ⅴ)的高。Ren等[17]的研究发现,水稻对Sb(Ⅴ)的转运系数最高为Sb(Ⅲ)转运系数的3~4倍,这与本研究的结果不一致,可能与水稻的品种有关。Huang等[18]的研究表明:Sb(Ⅲ)和Sb(Ⅴ)处理下,三个水稻品种(Yangdao 6、Nongken 57、Jiahua 1)表现出不同的Sb富集及转运能力。Shtangeeva等[19]的研究结果也表明:不同植物对Sb的吸收能力存在差异,他们发现在75mg·L-1水平下,Sb(Ⅲ)处理黑麦(Secale cereal L.)各部位Sb含量低于Sb(Ⅴ)处理,而小麦(Triticum astevium L.)各部位Sb含量则呈现相反的结果。
根系是植物与土壤环境进行物质交换的重要器官。根系的生长发育能影响植物对营养物质的吸收及有害物质的排泄,其形态特征能够说明植物受重(类)金属毒害程度,植物能通过改变根系形态特征来降低重(类)金属胁迫程度[20-21]。有报道表明:Sb(Ⅲ)和Sb(Ⅴ)均能抑制水稻根系的生长[6]。本研究发现:Sb(Ⅲ)和Sb(Ⅴ)均显著抑制了水稻根系的生长发育,相对于Sb(Ⅲ),Sb(Ⅴ)处理对水稻根系形态各指标的影响较小。当处理浓度小于10 mg·L-1时,Sb(Ⅴ)基本未对根系形态指标有显著性影响;而在20 mg·L-1处理水平下,Sb(Ⅴ)处理根系形态指标(除根直径外)均明显高于Sb(Ⅲ)处理。但在10 mg·L-1和20 mg·L-1处理水平下,Sb(Ⅴ)和Sb(Ⅲ)处理对水稻根系生物量的影响相似。He等[5]研究发现,在水稻种子(Yuhong No.1)萌发15 d时,与对照相比,1000 mg·L-1Sb(Ⅲ)处理干物质转换率为38.79%,而1000 mg·L-1Sb(Ⅴ)处理干物质转换率约为20%(此数据为作者根据该文所示图估算出)。上述结果表明:Sb(Ⅲ)处理水稻干物质转换效率高于Sb(Ⅴ)处理,导致Sb(Ⅲ)处理水稻根系鲜重与Sb(Ⅴ)处理相似,而Sb(Ⅴ)处理下根系形态特征参数明显高于Sb(Ⅲ)处理。
报道表明,植物能通过减少细根数量、根系表面积及分叉数来降低根与重(类)金属的接触面积,从而降低对重(类)金属的吸收[22-23]。本研究中Sb(Ⅲ)处理显著降低了大多数根系形态参数,增加了水稻中等根和粗根比例、降低了细根比例;Sb(Ⅴ)处理下对水稻根形态参数及各种根比例的影响较小。上述结果表明Sb(Ⅲ)对水稻的毒性较Sb(Ⅴ)高,且水稻根系粗度及形态参数的变化与不同形态Sb对水稻的毒害程度有关。
4 结论
(1)Sb(Ⅲ)和Sb(Ⅴ)均对水稻生长有害,且Sb(Ⅲ)毒性较Sb(Ⅴ)高。
(2)水稻根系富集了大多数Sb,Sb(Ⅲ)处理水稻地上部Sb含量高于Sb(Ⅴ),而根系中则相反。
(3)与对照相比,10 mg·L-1和20 mg·L-1Sb(Ⅲ)均显著降低绝大多数根系形态参数的数值;Sb(Ⅴ)仅20 mg·L-1处理水平显著抑制了水稻根面积、根直径和根分叉数。
(4)Sb(Ⅲ)处理增加水稻根系的粗度,而Sb(Ⅴ)处理对水稻根系粗细程度的影响较小。
[1]张亚平,张婷,陈锦芳,等.水、土环境中锑污染与控制研究进展[J].生态环境学报,2011,20(8/9):1373-1378.
ZHANGYa-ping,ZHANGTing,CHENJin-fang,et al.Research progress on present situation and countermeasure of antimony pollution in water and soil environment[J].Ecology and Environmental Sciences,2011,20(8/9):1373-1378.
[2]王淑玲.中国锑资源现状及可持续发展问题探讨[J].中国金属通报, 2002,47:6-10.
WANG Shu-ling.Current situation and sustainable development of antimony resources in China[J].China Metal Bulletin,2002,47:6-10.
[3]齐翠翠.锑在中国煤及典型矿区中的环境地球化学研究[D].合肥:中国科学技术大学,2010.
QI Cui-cui.Environmental geochemistry of antimony in Chinese coals and two typical mines[D].Hefei:University of Science and Technology of China,2010.
[4]齐文启,曹杰山.锑(Sb)的土壤环境背景值研究[J].土壤通报,1991, 22(5):209-210.
QI Wen-qi,CAO Jie-shan.Study on the soil environmental background values of antimony(Sb)[J].Chinese Journal of Soil Science,1991,22(5):209-210.
[5]He M C,Yang J R.Effects of different forms of antimony on rice during the period of germination and growth and antimony concentration in rice tissue[J].The Science of the Total Environment,1999,243/244:149-155.
[6]刘碧君,吴丰昌,邓秋静,等.锡矿山矿区和贵阳市人发中锑、砷和汞的污染特征[J].环境科学,2009,30(3):907-912.
LIU Bi-jun,WU Feng-chang,DENG Qiu-jing,et al.Pollution characteristics of antimony,arsenic and mercury in human hair at Xikuangshan antimony mining area and Guiyang City,China[J].Environmental Science,2009,30(3):907-912.
[7]戈兆凤,韦朝阳.锑环境健康效应的研究进展[J].环境与健康杂志, 2011,28(7):649-653.
GE Zhao-feng,WEI Chao-yang.Environmental health effect of antimony:Areviewofrecentresearches[J].JournalofEnvironmentandHealth,2011,28(7):649-653.
[8]Baroni F,Boscagli A,Protano G,et al.Antimony accumulation in achillea ageratum,plantago lanceolata and silene vulgaris growing in an old Sb-mining area[J].Environmental Pollution,2000,109(2):347-352.
[9]Tschan M,Robinson B,Nodari M,et al.Antimony uptake by different plantspeciesfromnutrientsolution,agarandsoil[J].EnvironmentalChemistry,2009,6(2):144-152.
[10]黄益宗,胡莹,刘云霞,等.锑对水稻根生长的影响及锑的吸收动力学特性[J].环境化学,2010,29(4):640-643.
HUANG Yi-zong,HU Ying,LIU Yun-xia,et al.Effect of antimony on roots growth and kinetics of antimony uptake by rice roots[J].Environmental Chemistry,2010,29(4):640-643.
[11]蓝唯源,宋书巧,吴浩东,等.土壤三价锑污染对甜芥菜生长及品质的影响研究[J].环境科学与技术,2009,32(2):20-23.
LAN Wei-yuan,SONG Shu-qiao,WU Hao-dong,et al.Effect of soil antimony(Ⅲ)pollution on the growth and quality of sweet mustard[J].Environmental Science and Technology,2009,32(2):20-23.
[12]Shtangeeva I,Bali R,Harris A.Bio-availability and toxicity of antimony[J].Journal of Geochemical Exploration,2011,110(1):40-45.
[13]何俊瑜,任艳芳,王阳阳,等.不同耐性水稻幼苗根系对镉胁迫的形态及生理响应[J].生态学报,2011,31(2):522-528.
HE Jun-yu,REN Yan-fang,WANG Yang-yang,et al.Root morphological and physiological responses of rice seedling with different tolerance to cadmium stress[J].Acta Ecologica Sinica,2011,31(2):522-528.
[14]刘莹,盖钧镒,吕彗能.作物根系形态与非生物胁迫耐性关系的研究进展[J].植物遗传资源学报,2003,4(3):265-269.
LIU Ying,GAI Jun-yi,LÜ Hui-neng.Advances of the relationship between crop root morphology and tolerance to antibiotic stress[J].Journal of Plant Genetic Resources,2003,4(3):265-269.
[15]何孟常,万红艳.环境中锑的分布、存在形态及毒性和生物有效性[J].化学进展,2004,16(1):131-135.
HE Meng-chang,WAN Hong-yan.Distribution,speciation,toxicity and bio-availability of antimony in the environment[J].Progress in Chemistry,2004,16(1):131-135.
[16]向猛,黄益宗,蔡立群,等.外源钙对两种价态锑胁迫下水稻幼苗吸收积累锑和钙的影响[J].生态毒理学报,2015,10(3):153-160.
XIANG Meng,HUANG Yi-zong,CAI Li-qun,et al.Effect of calcium on uptake and accumulation of antimony and calcium by rice seedling in solution culture[J].Asian Journal of Ecotoxicology,2015,10(3):153-160.
[17]Ren J H,Ma L Q,Sun H J,et al.Antimony uptake,translocation and speciation in rice plants exposed to antimonite and antimonate[J].Science of the Total Environment,2014,475:83-89.
[18]Huang Y C,Chen Z,Liu W J.Influence of iron plaque and cultivars on antimony uptake by and translocation in rice(Oryza sativa L.)seedlings exposed to Sb(Ⅲ)or Sb(Ⅴ)[J].Plant and Soil,2012,352(1/2):41-49.
[19]Shtangeeva I,Steinnes E,Lierhagen S.Uptake of different forms of antimony by wheat and rye seedlings[J].Environmental Science and Pollution Research,2012,19(2):502-509.
[20]Li F,Li M Y,Pan X H,et al.Biochemical and physiological characteristics in seedlings roots of different rice cultivars under low phosphorus stress[J].Chinese Journal of Rice Science,2004,18(1):48-52.
[21]Zhang E H,Zhang X H,Wang H Z.Adaptable effects of phosphorus stress on different genotypes of faba-bean[J].Acta Ecologica Sinica, 2004,24(8):1589-1593.
[22]Das P,Samantaray S,Rout G R.Studies on cadmium toxicity in plants:A review[J].Environmental Pollution,1997,98(1):29-36.
[23]何俊瑜,王阳阳,任艳芳,等.镉胁迫对不同水稻品种幼苗根系形态和生理特性的影响[J].生态环境学报,2009,18(5):1863-1868.
HE Jun-yu,WANG Yang-yang,REN Yan-fang,et al.Effect of cadmium on root morphology and physiological characteristics of rice seedlings[J].Ecology and Environmental Sciences,2009,18(5):1863-1868.
Effects of different forms of antimony on the growth and root morphology of rice plant
LEI Lei1,2,3,WU Qian-hua1,2,3,LÜ Hai-qin1,2,4,WANG Rui-gang1,2,DING Yong-zhen1,XU Ying-ming1,2,FAN Zhi-lian3,MO Liang-yu3,FENG Ren-wei1,2,3,4,*
(1.Institute of Agro-Environmental Protection,The Ministry of Agriculture,Tianjin 300191,China;2.Innovation Team of Remediation of Heavy Metal Contaminated Farmland Soil,Chinese Academy of Agricultural Sciences,Tianjin 300191,China;3.Agricultural College,Guangxi University,Nanning 530004,China;4.College of Resources and Environment,Huazhong Agricultural University,Wuhan 430070,China)
This study was conducted to investigate the effects of different forms of antimony(antimonite[Sb(Ⅲ)]and antimonate[Sb(Ⅴ)])on Sb concentration in different organs of rice plant(Fengmeizhan),plant growth and plant root morphology using a hydroponic culture system containing Espino nutrient solution.The results show that the additions of Sb(Ⅲ)and Sb(Ⅴ)both showed a negative effect on the growth of rice plants,and the toxicity of Sb(Ⅲ)is higher than that of Sb(Ⅴ)to this rice plant.The supplementation of Sb(Ⅲ)and Sb(Ⅴ)to the solution significantly enhanced the Sb concentrations in all organs of this rice plant,and most uptake Sb was concentrated in the roots. This rice plants showed a weaker ability to transport Sb(Ⅴ)from the roots to the shoots relative to Sb(Ⅲ),which resulted in the shoot Sb concentration of this rice plant subjected to Sb(Ⅲ)exposure being higher,but root Sb concentration being lower than that subjected to Sb(Ⅴ).The addition of Sb(Ⅲ)significantly reduced most tested parameters of root morphology,including root length,root average area,root volume,root tip number and root fork number.However,only 20 mg·LSb(Ⅴ)remarkably reduced the root average area,root diameter and root fork number.Above results suggested that the rice plant might alter the root morphology to adapt the Sb exposure,and reduce the Sb uptake via reducing the root average area and root fork number.With the increasing Sb(Ⅲ)levels in the solution,the fine root proportion decreased from 52.1%to 41.5%,and the medium root proportion increased from 45.2%to 55.2%,indicating that the rice plant could reduce the Sb uptake through enhancing the coarseness of root exposing to Sb(Ⅲ).However,the addition of Sb(Ⅴ)showed a limited effect on the proportions of fine roots and medium roots,which fell into the range of 52.1%~56.8%and 41.7%~45.2%,respectively.
different forms of antimony;rice;toxicity;root morphology;absorption
S511
A
1672-2043(2017)03-0437-06
10.11654/jaes.2016-1212
雷蕾,吴倩桦,吕海琴,等.不同形态锑对水稻生长及根系形态特征的影响研究[J].农业环境科学学报,2017,36(3):437-442.
LEI Lei,WU Qian-hua,LÜ Hai-qin,et al.Effects of different forms of antimony on the growth and root morphology of rice plant[J].Journal of Agro-Environment Science,2017,36(3):437-442.
2016-09-18
雷蕾(1992—),女,山西运城人,硕士研究生,主要从事土壤污染与修复研究。E-mail:leilaxj@163.com
*通信作者:冯人伟E-mail:frwzym@aliyun.com
国家自然科学基金项目(41473114,41103075)
Project supported:The National Natural Science Foundation of China(41473114,41103075)
