茂兰自然保护区四药门花群落结构与物种多样性研究
2017-03-14陈晓熹申长青洪文君曾思金周玉洁黄久香
陈晓熹,申长青,2,洪文君,3,曾思金,周玉洁,黄久香
(1.华南农业大学 中国南方石灰岩植物研究中心,华南农业大学林学与风景园林学院,广东 广州 510642;2.乳源瑶族自治县林业局,广东 韶关 512700;3.三亚市林业科学研究院,海南 三亚 572023)
四药门花Loropetalum subcordatum为金缕梅科Hamamelidaceae檵木属Loropetalum常绿灌木或小乔木[1]。四药门花首次发现于香港,之后的近10 a曾一度被认为是香港的特有种。直到1957年,才相继在广西壮族自治区龙州、贵州省茂兰和广东省五桂山发现极为稀少的植株[2]。因其数量少、分布区狭小,自然更新困难,已被IUCN[1]列为易危种,也是我国Ⅱ级珍稀濒危保护植物[3];四药门花在解剖学及形态学上被普遍认为是金缕梅亚科中最为原始的代表,对研究金缕梅亚科的系统演化有特别重要的意义[4],具有较高的保护和利用价值。
对极小种群植物的研究是近几年珍稀濒危植物研究热点之一,其中对四药门花的研究也是植物学家的关注焦点。施苏华等[5]和章群等[6]根据ITS序列探讨了四药门花的系统位置;顾垒等[1]对广东五桂山的四药门花种群授粉进行了研究;Gong等[7]和李柏君[8]对不同地区的四药门花群体进行了遗传结构分析;陈真权等[9]和黄柔柔等[10]对广东五桂山四药门花种群进行了生态学研究;刘华[4]和何妙坤等[2]分别开展了四药门花的扦插繁殖研究;孙红梅等[11]分析了不同激素对四药门花形成愈伤组织和萌芽培养的影响;申长青等[12-14]研究了四药门花对光照和水分胁迫的光合生理的响应机制,以及修剪和多效唑对其生长的影响;洪文君等[15]比较了四药门花、檵木L.chinense和红花檵木L.chinense var.rubrum三种檵木属植物的根、茎、叶的解剖特征,结果表明四药门花植株在解剖特征上表现出对荫凉或湿生生境具有较好的适应性。这些研究为四药门花的保护和发展提供了宝贵的资料。四药门花茂兰种群的遗传多样性最高,是最重要的保护群落[7-8],有人已对茂兰自然保护区植物群落的结构和物种多样性进行了研究[16-23],但针对该地四药门花种群的研究尚未见相关报导。群落特征不仅体现群落中物种的生长情况,也反映它们与环境之间的相互适应关系,对四药门花开展种群结构和多样性研究,了解群落的物种组成和更新动态,可以为四药门花的保护提供科学依据[24-25]。
1 材料和方法
1.1 研究地概况
贵州省茂兰国家级自然保护区处于贵州省南部黔南布依族苗族自治州荔波县东南部,107°52'10''~108°05'40''E,25°09'20''~25°20'50''N,是保护完整的综合自然生态系统以及珍稀孑遗动植物资源为目的的科学保护区[[26]。1984年成立,1988年经国务院批准为国家级自然保护区。喀斯特森林为主体,属中亚热带季风温润气候,年平均气温为15.3℃,气温年均差18.3℃,1月平均气温为5.2℃,7月平均气温为23.5℃,年均降水量1 752 mm[27]。茂兰自然保护区喀斯特地貌十分典型,海拔430~1 078.6 m[17],土壤略碱性,多为黑色石灰土,有机质和全氮含量丰富[28]。
1.2 调查方法
在踏查的基础上,采用典型格子样方法,于2015年10月在保护区内四药门花分布较丰富的地段设置了4个20 m×20 m的样地,总面积1 600 m2。每个样地划分为4个10 m×10 m的小样方,记录每个小样方内所有胸径(diameter at breast height,DBH)≥2.0 cm的树木种类、树高、胸径、冠幅及株数;在每个小样方分别选取1个5 m×5 m和1 m×1 m的样方,分别记录灌木层和草本层各植物的种名、株数、株高和盖度[29]。同时,记录所有四药门花植株的胸径、树高和冠幅。根据树高(height,H)和胸径大小,将四药门花植株划分为5个等级[30]:Ⅰ级幼苗H<33 cm,DBH<2.5 cm;Ⅱ级小树H≥33 cm,DBH<2.5 cm;Ⅲ级幼树2.5 cm≤DBH<7.5 cm;Ⅳ级中树7.5cm≤DBH<22.5 cm;Ⅴ级大树DBH≤22.5 cm。
1.3 数据处理
1.3.1 重要值计算[31]
乔木层重要值(IV)=相对多度(RA)+相对频度(RF)+相对显著度(RD)
灌木层重要值(IV灌)=相对多度(RA)+相对频度(RF)
草本层重要值(IV草)=相对盖度(RC)+相对频度(RF)
1.3.2 丰富度、多样性和均匀度指数计算[22]
Margalef丰富度指数(dMa):

Simpson多样性指数(D):

Shannon-Wiener多样性指数(H’):
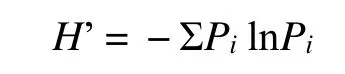
Pielou均匀度指数(J):
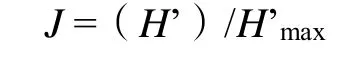
上述式中,S为物种数,N为植株数,Pi为物种i的个体数占群落中全部个体数的比例。
1.3.3 数据统计
本文数据统计、作图均在Excel 2007中完成。
2 结果与分析
2.1 四药门花群落的物种组成
本次调查共记录到维管植物142种,隶属于60科104属。其中蕨类植物6科7属9种,裸子植物1科1属1种,单子叶植物4科6属6种,双子叶植物49科90属126种,详见表1。
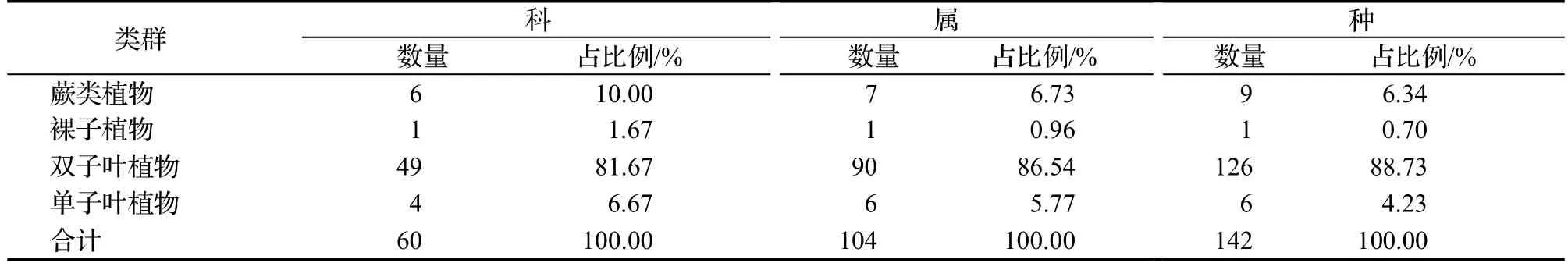
表1 茂兰自然保护区四药门花群落的物种组成Table 1 Species composition of L.subcordatum communities in Maolan National Nature Reserve
在样地中,5种以上的科共7科,其中以樟科Lauraceae,山茶科Theaceae,桑科Moraceae,大戟科Euphorbiaceae和茜草科Rubiaceae的植物最为丰富,7种以上;其次是蔷薇科Rosaceae和紫金牛科Myrsinaceae,分别为6种和5种。优势科的分布区类型均为泛热带分布型和世界分布型[32](见表2)。

表2 四药门花群落优势科及其属种数Table 2 Genera and species of dominant families at studied communities
2.2 四药门花群落的优势种分析
在样地乔木层中,共记录到69种,隶属36科54属,重要值排前10的树种如表3所示。其中四药门花和香叶树Lindera communis在四个样地均有较高的相对多度和相对显著度,表明这2个树种在群落中有占绝对优势和较高的密度;马尾松Pinus massoniana和枫香树Liquidambar formosana在群落中都有大树,但相对多度较低,反映了这两个树种在群落中幼苗较少,更新困难。
灌木层72种,隶属37科55属,重要值排前10的树种见表4。该层物种数最为丰富,其中香叶树占绝对优势;杜茎山Maesa japonica和红鳞蒲桃Syzygium hancei在4个样地中都有出现,且有较高的相对多度,表明这2个树种在群落中有较强的自然更新能力。

表3 四药门花群落乔木层优势种及其重要值(前10种)Table 3 Dominant species and their importance values at tree layer(The top 10 species)

表4 四药门花群落灌木层优势种及其重要值(前10种)Table 4 Dominant species and their importance values at shrub layer(The top 10 species)
草本层14种,隶属11科12属,重要值排前10的树种见表5。该层中物种数最少,其中翠云草Selaginella uncinata优势度最高,薄叶卷柏S.delicatula较为丰富。在1 m×1 m的样方中还记录到香叶树,石岩枫,粗叶木Lasianthus chinensis,硃砂根Ardisia crenata,地果Ficus tikoua,杜鹃Rhododendron simsii,狭叶润楠Machilus rehderi等木本植物的小苗。此外,小样方中主要的层外植物有石岩枫,天门冬Asparagus cochinchinensis,常春藤Hedera sinensis,乌蔹莓Cayratia japonica,地果,络石Trachelospermum jasminoides,鸡矢藤Paederia scandens等。

表5 四药门花群落草本层优势种及重要值(前10种)Table 5 Dominant species and their importance values at herb layer(The top 10 species)
2.3 四药门花群落种子植物属的地理成分分析
依照吴征镒[32]对中国种子植物属分布类型的划分标准,除蕨类植物外,记录到的97属共有12种地理成分。以热带分布属为主,占非世界分布属的67.02%,其中以泛热带分布最为丰富,占22.34%;中国特有分布1属(表6)。

表6 四药门花群落种子植物属的分布类型Table 6 Generic geographical elements of the spermatophytes of the studied plot
2.4 四药门花群落的物种多样性分析
对四药门花群落物种多样性进行分析,结果表明(表7),群落中物种丰富度指数由高到低排序为灌木层>乔木层>草本层,说明群落中乔、灌木的物种较丰富,草本层物种较匮乏。Simpson多样性指数反映个体的优势度,数值越小优势种的优势地位越明显;Shannon-Wiener多样性指数反映个体的集中度,数值越大,物种集中度越低,分布越均匀[33]。Simpson多样性指数和Shannon-Wiener多样性指数表现出一致性,由高到低顺序为灌木层>乔木层>草本层,这是乔木层中的四药门花和香叶树、草本层中的翠云草和薄叶卷柏明显的优势地位造成的。Pielou均匀度指数由高到低排序为灌木层>草本层>乔木层,说明灌木层物种较多且分布均匀。

表7 四药门花群落物种的多样性Table 7 Species diversity of L.subcordatum communities
2.5 与不同地区植物群落多样性比较
将茂兰自然保护区四药门花群落与5个不同地区不同植物群落乔木层的多样性指数进行比较,结果见表8。由表8可以看到,茂兰自然保护区四药门花群落乔木层的物种多样性指数与广东五桂山四药门花群落相近,高于大浩湖山黄麻Trema tomentosa群落和白湾石灰岩风水林,远低于茂兰自然保护区的顶级群落常绿落叶阔叶混交林、青冈Cyclobalanopsis glauca-黄樟Cinnamomum parthenoxylon群落。根据龙翠玲等[16]、王周平等[34]和何永涛等[35]的相关研究,茂兰四药门花群落的乔木层多样性指数较低、稳定性差、灌木层结构相对发达,表明正处于演替中期向顶级群落发展。

表8 茂兰自然保护区四药门花群落和其他群落乔木层物种多样性比较Table 8 Comparison of species diversity at tree layer among L.subcordatum communities in Maolan National Nature Reserve and other communities in different locations
2.6 四药门花种群径级结构
在4个样地中,共记录到四药门花165株,Ⅲ级幼树数量最多,共96株,占总个体的58%;Ⅰ,Ⅱ,Ⅳ级个体数分别为21,32和16株,未记录到Ⅴ级以上的大树。该种群缺少Ⅰ级幼苗和Ⅱ级小树,自然更新能力弱,径级结构呈纺锤形,表现为衰退型种群(图1)。
3 结论与讨论
茂兰四药门花种群主要分布于海拔700~900 m的常绿阔叶林群落中,在调查中发现樟科、山茶科和桑科等地带性植物种类丰富,在种子植物属的分布区类型上以热带分布为主,占非世界分布属的67.02%,略高于茂兰青冈-黄樟群落(60.71%)和茂兰喀斯特常绿落叶阔叶混交林群落(65.31%),3个群落的优势科均为樟科,表明该地四药门花群落的植物区系与热带亚洲植物区系以及泛热带植物区系关系密切,有喀斯特常绿阔叶林的特征。

图1 茂兰自然保护区四药门花的径级分布Figure 1 Diameter distribution of L.subcordatum in Maolan National Nature Reserve
从群落的物种多样性分析来看,茂兰四药门花群落灌木层的多样性指数最高,与刘济明等[23]和徐瑞晶等[37]的研究结果相似。出现该结果的原因可能是四药门花和香叶树相比乔木层的其他树种更适应该地的气候和土壤条件,抢占了更多的资源;灌木层则在林冠层的保护下,林下生境逐渐改善,树种多样性得到发展;而草本层因为光照条件相对其他两个层次较差,物种多样性也相对较少。随着群落的发展,林分郁闭度不断增大,灌木和草本物种数和个体数开始不断减少,乔木层树种逐渐占据优势。
与同一地区青冈-黄樟群落[23]和常绿落叶阔叶混交林群落[18]相比,茂兰四药门花群落物种数量不及其他两地,多样性指数更低且优势科与优势种也不同。出现该结果的原因一是茂兰自然保护区为喀斯特地貌,生境复杂多变,不同的小生境对物种组成、群落结构都会构成影响;二是其他两地为顶级群落,结构稳定,而茂兰四药门花群落正处演替中期,结构相对简单,稳定性和多样性较低;还有可能是因为调查设置的样地大小不一,造成调查结果的差异。
与广东省五桂山四药门花群落相比,两地的优势种仅四药门花,且茂兰四药门花群落物种丰富度更高,种类更多,物种组成差异较大。同一植物群落在不同生境下的差异,反映了土壤养分的空间异质性对群落结构有显著影响[38-39]。茂兰自然保护区为典型的喀斯特地貌,土壤以黑色石灰土为主;而五桂山四药门花群落则分布于低山丘陵乱石山地,土壤以山地赤红壤为主,进一步验证了土壤性质一定程度上影响到植物群落结构[22],不同的土壤肥力使群落的物种丰富度产生较大差异[40]。此外,人为因素的影响也是导致两地群落结构产生差异的原因之一,茂兰四药门花群落保护较好,而广东五桂山四药门花群落曾遭受过较严重的人为破坏,现存的母树多为砍伐后的萌生株[9]。从径级结构来看,茂兰四药门花群落共记录到四药门花植株165株,以Ⅲ级幼树为主,占总数的58%;广东五桂山四药门花群落共记录到四药门花植株56株,以Ⅲ级幼树为优势,占总数的58.9%。两地的四药门花群落径级结构相似,缺乏幼苗,呈纺锤形,均为衰退型种群。在调查过程中,发现保护区空旷的山路旁有较多的四药门花幼苗,而在郁闭度较高的林内则较少或没有,说明四药门花种子萌发需要一定的光照,而洪文君等[15]的研究表明四药门花植株喜荫凉、湿生的生境,种子和植株间生态特性的矛盾可能是导致四药门花易危的原因之一。调查发现四药门花在原生群落中根部萌发普遍。顾垒等[1]的研究结果表明,四药门花为自花传粉的植物,自然结实率较低,是其种群自然更新困难主要原因之一;Gong等[7]和李柏君[8]的研究表明四药门花种群遗传多样性较低、群体遗传分化较大,种群中存在无性克隆,这可能与其分布区隔离及其自身的繁殖特性关系密切。四药门花种群缺乏幼苗,自然更新困难,为衰退型种群;而群落中香叶树种群的幼苗数量较大,更新良好,为稳定增长型种群。随着群落的演替,四药门花可能会被更新能力更强的树种取代,有灭绝的危险。因此建议进行适当地疏林、开林窗改善林下生境以提高四药门花幼苗的存活率。
致谢:感谢熊志斌高级工程师以及贵州茂兰国家级自然保护区管理局对调查工作提供的帮助与支持!
[1]顾垒,张奠湘.濒危植物四药门花的自花授粉[J].植物分类学报,2008,46(5):651-657.
[2]何妙坤,黄久香,黄川腾,等.四药门花扦插繁殖技术研究[J].林业实用技术,2013(11):50-53.
[3]于永福.中国野生植物保护工作的里程碑——《国家重点保护野生植物名录(第一批)》出台[J].植物杂志,1999(5):3.
[4]刘华.四药门花扦插繁殖技术[J].现代园艺,2013(12):33-34.
[5]施苏华,黄椰林,章群,等.四药门花属及其近缘植物ITS区序列分析和系统学意义[J].云南植物研究,1999,21(1):89-95.
[6]章群,施苏华,黄椰林,等.金缕梅亚科ITS序列分析及其系统学意义[J].武汉植物学研究,2001(6):445-448.
[7]Gong W,Gu L,Zhang D.Low genetic diversity and high genetic divergence caused by inbreeding and geographical isolation in the populations of endangered species Loropetalum subcordatum(Hamamelidaceae)endemic to China[J].Conserv Genet,2010,11(6):2281-2288.
[8]李柏君.基于SRAP分子标记的四药门花种群遗传多样性分析[D].广州:华南农业大学,2016.
[9]陈真权,庄雪影,黄久香,等.广东五桂山四药门花种群生态学研究[J].广东林业科技,2009,25(5):41-45.
[10]黄柔柔,叶子霖,张倩媚,等.四药门花(Loropetalum subcordatum)的生态生物学特性研究[J].生态科学,2016,35(3):52-55.
[11]孙红梅,陈彦,梁银兴,等.濒危植物四药门花的组织培养研究初探[J].现代园艺,2015(2):12-13.
[12]申长青.濒危植物四药门花对光照和水分胁迫的适应性研究[D].广州:华南农业大学,2016.
[13]申长青,陈晓熹,周玉洁,等.多效唑对四药门花生长、叶绿素及叶绿素荧光的影响[J].林业科技通讯,2017(5):55-59.
[14]申长青,洪文君,黄久香,等.修剪和多效唑对四药门花矮化效应初探[J].广东园林,2016,38(1):46-48.
[15]洪文君,黄久香,申长青,等.3种檵木属植物结构解剖及环境适应性评价[J].南京林业大学学报(自然科学版),2016,40(5):61-66.
[16]龙翠玲,余世孝,熊志斌,等.茂兰喀斯特森林林隙的植物多样性与更新[J].生物多样性,2005,13(1):43-50.
[17]龙翠玲.不同地形部位喀斯特森林物种多样性的比较研究——以贵州茂兰自然保护区为例[J].中国岩溶,2007,26(1):55-60.
[18]胡蕖,魏鲁明,杨成华,等.茂兰喀斯特峰丛中部常绿落叶阔叶混交林群落特征及多样性研究[J].福建林业科技,2009,36(1):43-48.
[19]兰洪波,冉景丞,蒙惠理,等.茂兰自然保护区生物物种多样性及其保护[J].山地农业生物学报,2009,28(2):119-125.
[20]玉屏,兰洪波,冉景丞,等.茂兰自然保护区生物多样性现状及保护对策[J].现代农业科技,2011(15):233-236.
[21]周凤娇,丁访军,杨成华,等.茂兰自然保护区旅游景点植物群落结构及多样性[J].贵州农业科学,2012,40(2):8-12.
[22]郎华林,龙翠玲.茂兰自然保护区喀斯特森林物种多样性与土壤因子的关系[J].湖北农业科学,2012,51(18):3987-3990.
[23]刘济明,文爱华,池馨,等.茂兰青冈栎-黄樟群落特征[J].湖北农业科学, 2014,53(14):3326-3331.
[24]杜虎,彭晚霞,宋同清,等.桂北喀斯特峰丛洼地植物群落特征及其与土壤的耦合关系[J].植物生态学报,2013,37(3):197-208.
[25]郭华,张桂萍,铁军,等.太行山南段油松群落物种多样性研究[J].植物科学学报,2015,33(2):151-157.
[26]令狐克鸿,覃龙江,陈正仁,等.茂兰自然保护区珍稀濒危植物石山木莲的资源现状及保护[J].中国农学通报,2012,28(28):100-103.
[27]杨瑞,喻理飞,安明态.喀斯特区小生境特征现状分析:以茂兰自然保护区为例[J].贵州农业科学,2008,36(6):168-169.
[28]郑振宇,龙翠玲.茂兰喀斯特森林不同地形部位木本植物种间联结性分析[J].林业资源管理,2014(4):78-84.
[29]洪文君,魏依娜,黄川腾,等.广州南沙黄山鲁白楸次生林的群落特征研究[J].湖北民族学院学报(自然科学版),2013,31(4):370-376.
[30]苏志尧,吴大荣,陈北光.粤北天然林优势种群结构与空间格局动态[J].应用生态学报,2000,11(3):337-341.
[31]李贵详,柴勇,邵金平,等.大树杜鹃群落结构特征研究[J].浙江林业科技,2016,36(3):1-7.
[32]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991,13(S4):1-139.
[33]吴毅,周国英,祁承经,等.湖南紫金山马尾松的群落特征及物种多样性分析[J].中南林业科技大学学报,2011,31(11):120-124.
[34]王周平,李旭光,石胜友,等.缙云山森林林隙与非林隙物种多样性比较研究[J].应用生态学报,2003,14(1):7-10.
[35]何永涛,李贵才,曹敏,等.哀牢山中山湿性常绿阔叶林林窗更新研究[J].应用生态学报,2003,14(9):1399-1404.
[36]马晓东,饶显龙,王发国,等.佛山市大浩湖山黄麻群落特征及物种多样性研究[J].广东农业科学,2014(21):153-158.
[37]徐瑞晶,庄雪影,莫惠芝,等.清远白湾石灰岩山区村落风水林植物物种多样性研究[J].华南农业大学学报,2012,33(4):513-518.
[38]苏松锦,刘金福,何中声,等.格氏栲天然林土壤养分空间异质性[J].生态学报,2012,32(18):5673-5682.
[39]张忠华,胡刚,祝介东,等.喀斯特森林土壤养分的空间异质性及其对树种分布的影响[J].植物生态学报,2011,35(10):1038-1049.
[40]张萍,冯志立.西双版纳热带雨林次生林的生物养分循环[J].土壤学报,1997,34(4):418-426.
