封育和放牧对黄土高原典型草原芽库的影响
2017-03-02赵凌平赵芙蓉
崔 伟,赵凌平,2,赵芙蓉,2
(1.河南科技大学动物科技学院,河南 洛阳 471003;2.河南省饲草饲料资源开发与畜禽健康养殖院士工作站,河南 洛阳 471003)
封育和放牧对黄土高原典型草原芽库的影响
崔 伟1,赵凌平1,2,赵芙蓉1,2
(1.河南科技大学动物科技学院,河南 洛阳 471003;2.河南省饲草饲料资源开发与畜禽健康养殖院士工作站,河南 洛阳 471003)
在多年生草地生态系统中,芽库在干扰后的种群补充和更新中占据着重要地位。本研究采用单位面积挖掘取样法,比较分析了封育和放牧对黄土高原典型草原地上植被特征和芽库密度的影响。结果表明,黄土高原典型草原芽库以分蘖芽为主,密度为832~6 848芽·m-2。与放牧相比,短期封育显著提高了芽库总密度和禾草芽库密度,但显著降低了非禾草芽库密度(P<0.05)。封育和放牧对典型草原芽库密度的显著影响,主要来自根茎型、分蘖型和根蘖型牧草芽库的贡献。短期封育显著提高根茎型和分蘖型牧草的芽库密度,而显著降低根蘖型牧草的芽库密度(P<0.05)。禾草芽密度与地上生物量呈显著正相关,与地上茎分枝数呈显著负相关(P<0.05)。封育和放牧对黄土高原典型草原地上植被特征的影响可能是通过调控禾草芽库密度来实现的。在草地管理中,芽库对地上植被的发展有一定的指示作用,可以依靠芽库的测定来预测未来植被的产量、演替趋势和发展方向。
封育;芽库;典型草原;无性繁殖
多年生植物可以同时以有性繁殖和克隆繁殖进行种群的维持与更新[1]。在植物的繁殖库中,种子库和芽库分别是有性繁殖和无性繁殖的基础[2]。植物的繁殖库能够使植物在遭受破坏后迅速恢复,大大降低了灭绝的几率[3]。与种子库相比,芽库在多年生草地的种群更新中发挥着更重要的作用。例如,在北美高草草原,99%的地上枝条更新来自于地下芽库,而种子库在植被更新中的作用几乎可以忽略[4]。但是芽库在草地群落中的特征和作用研究相对贫乏,直到近几年芽库在草地生态系统中的特征、功能和作用才开始受到国内外学者的重视。主要是因为受到试验地点以及外界条件的严格限制,调查芽库十分困难,特别是调查地下芽库有时只能对草地进行破坏性试验。
芽库是营养繁殖的潜在来源,是所有潜在能进行营养繁殖的芽的集合[5]。不同植物种群的芽库动态变化能预测出群落的动态变化[6]。在草地生态系统中,芽库在植物种群的维持、群落生产力维持和对恶劣环境的恢复响应等方面发挥重要作用[4,7-9]。植物对芽休眠的调节是一个十分复杂的过程,可通过调节芽的输入和输出对干扰做出反应,进而影响着种群结构,群落的物种组成和动态变化。正确理解干扰调控芽库的机制对于探讨干扰影响机制,合理管理草地和解决生产实践问题是十分重要的。
多年来研究者对封育和放牧对草地生态系统的影响做过许多工作,其中包括封育和放牧管理措施下的植被特征、多样性变化和土壤理化性质的研究等[10-11]。但从繁殖库,特别是从芽库方面阐述封育和放牧对多年生草地的影响的研究鲜见报道。在黄土高原地区,有关种子库在草地生态系统中的特征和作用研究较多[12-15],而对芽库的研究较少。鉴于此,本研究分析封育和放牧对黄土高原典型草原芽库特征的影响,有助于预测典型草原植物群落动态变化和揭示干扰对芽库的影响机制,为退化草地经营管理和植被恢复提供理论依据。
1 材料与方法
1.1 研究区概况
试验设在宁夏回族自治区固原市东北部45 km处的黄土高原云雾山国家草原自然保护区(106°21′-106°27′ E,36°10′-36°17′ N)。该区平均海拔2 000 m,气候属于中温带半干旱气候,年平均降水量440.5 mm,且年内分布不均,降水主要集中在6月-9月。年平均气温为5 ℃,最热月为7月,气温为22~25 ℃,最冷月为1月,平均气温为-14 ℃左右。年平均蒸发量1 330~1 640 mm,≥0 ℃年积温2 370~2 882 ℃·d,年太阳总辐射量为523.35 kJ·cm-2,干燥度为1.5~2.0,全年无霜期137 d。保护区土壤类型简单,主要以淡黑垆土和黄绵土为主。植被区划为干草原区,以本氏针茅(Stipabungeana)、铁杆蒿(Artemisiasacrorum)、大针茅(Stipagrandis)、百里香(Thymusmongolicus)、星毛委陵菜(Potentillaacaulis)和冷蒿(Artemisiafrigida)等为优势种。
1.2 试验方法
在云雾山国家草原自然保护区中的实验区,选取放牧地和封育5年草地作为试验样地。放牧地为自然放牧状态,放牧强度为中度放牧(4.6羊单位·hm-2)。封育5年草地采用铁丝网围栏,完全排除家畜的采食和践踏。
芽库调查于2014年7月进行,这时植物正处于营养生殖高峰期。在封育5年草地和放牧地随机设置面积为50 m×50 m的5个小区,每个小区之间至少间隔50 m。芽库调查时,在每个小区内随机设置面积为25 cm×25 cm的6个样方,样方间隔至少5 m。芽库调查采用单位面积挖掘取样法。取样深度25 cm。取样时将样方内地上部分茎枝连同地下部分(根茎和根蘖等)一起挖出,用清水轻轻冲洗干净装入塑料袋带回实验室。注意保持地上植株与地下器官及全部营养芽的自然联系,以便鉴定与统计。参照参考文献[9]的方法进行芽库鉴定与统计。在解剖显微镜下,根据芽形态和芽所附着根系的形态鉴定芽库类型。本研究只统计明显的芽,可能形成根的分生组织不予统计。不同类型的植物需要不同的鉴定技术:对于游击型植物,通过肉眼即可辨认根茎上、根蘖上和匍匐茎上的芽;而需要借助解剖镜对位于丛生型植物基部的分蘖芽和根颈芽来鉴定芽的类型和数量。本研究中,根据植物芽所在的器官库进一步将芽分为分蘖芽、根蘖芽、根茎芽和根颈芽4类,忽略不计偶尔出现的其它芽类型。
2014年7月进行群落物种调查。在每个小区内另外再随机设置面积为50 cm×50 cm的4个样方,样方之间间距至少5 m。地上植被调查内容主要有群落植物种类和植被总盖度以及每种植物的个数、分枝数、盖度和地上生物量。
1.3 数据处理
物种多样性用Shannon-Wiener指数和物种丰富度指数来计算。
Shannon-Wiener指数(H)的计算公式为:
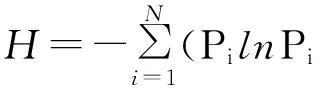
式中:N为物种总数,Pi为物种i的个数占所有物种总数的比例。
用变异系数(coefficientofvariation,CV)来衡量芽库密度的空间异质性。
变异系数的计算公式为:
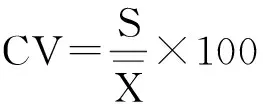
采用Excel2013对数据进行预处理,对封育地和放牧地地上植被特征和芽库密度之间的差异,采用t检验进行分析,用平均值和标准误表示测定结果,以P<0.05表示差异显著。用Pearson相关分析芽库密度与地上植被的密度、地上茎分枝数和地上生物量的相关性。以上所有分析均用SPSS16.0统计软件进行。
2 结果与分析
2.1 封育和放牧对地上植被特征的影响
在草地生态系统中,群落的结构与外貌常以优势种种类组成为特征。该区草地经过长期的演替发展,最终以本氏针茅为优势种的群落成为该区的主要群落。该群落在长期放牧利用下,形成了以本氏针茅、大针茅、百里香、冷蒿、铁杆蒿和星毛委陵菜等物种共同为优势种的群落,结构趋于简化,层次分化不明显,多为单层。在封育5年草地中,杂类草的地位下降,本氏针茅的优势地位明显提高,形成了以本氏针茅和大针茅为优势种的群落。
封育和放牧管理措施对黄土高原典型草原地上植被的密度、地上生物量、总盖度、地上茎分枝数、Shannon-Wiener多样性指数和丰富度均有显著影响(P<0.05)。与放牧地相比,封育5年草地显著增加了放牧地的地上生物量和总盖度,分别增长了56.8%和9.6%。但封育5年草地显著降低了地上植被密度、丰富度、Shannon-Wiener多样性指数和地上茎分枝数,分别下降了35.1%、25.3%、10.5%和21.7%(表1)。
2.2 封育和放牧对芽库密度的影响
在放牧地,芽库密度的变化范围是832~2 944芽·m-2,变异系数为27%;封育5年草地的芽库密度变化范围是1 040~6 848芽·m-2,变异系数为45%。可见,在放牧地,芽库密度的空间异质性较低。封育和放牧措施显著影响本氏针茅群落地下芽库总密度(P<0.05)。与放牧相比,短期封育显著增加了本氏针茅群落地下芽库的总密度,增长了74.1%(图1)。
在放牧地,禾草芽占到芽库的59.4%,非禾草芽占到芽库的40.6%;在封育5年草地,禾草芽占芽库的87.0%,非禾草芽占到芽库的13.0%。可见,短期封育保护了禾草芽免于动物的采食和践踏伤害,大大提高了禾草芽在芽库中的比例,降低了非禾草芽在芽库中的比例。封育和放牧措施对禾草和非禾草芽库密度均有显著影响(P<0.05)。与放牧地相比,封育5年草显著提高了禾草的芽库密度,而显著降低了非禾草的芽库密度(图2)。
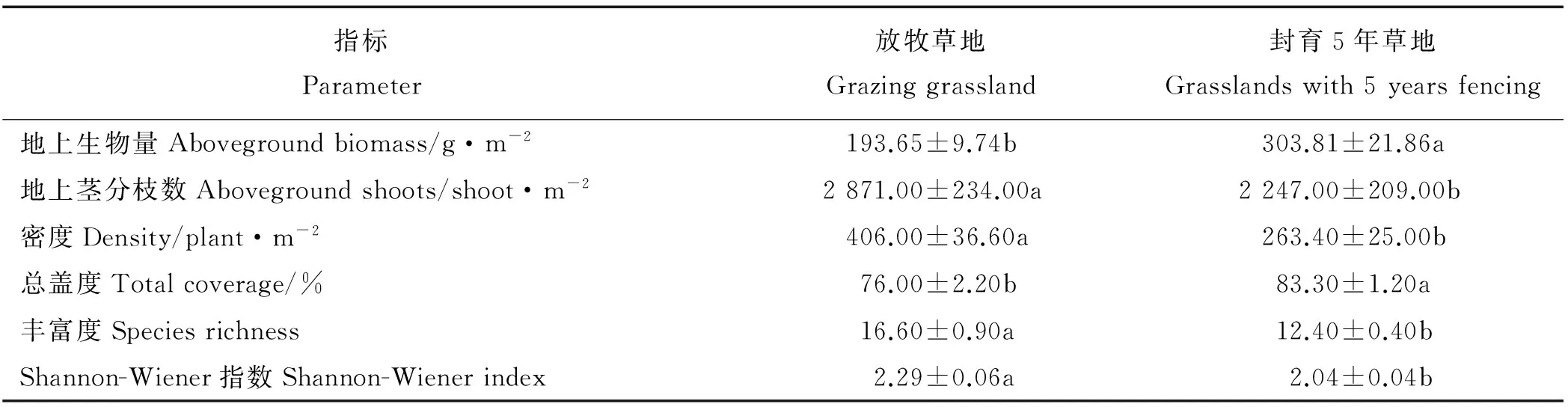
表1 封育和放牧措施下地上植被特征变化Table 1 Vegetation parameters changes under grazing and fencing management
注:同行不同小写字母表示两块样地间差异显著(P<0.05),下同。
Note: Different lower case letters in each row indicate significant difference between two sampling plots at the 0.05 level. The same below.
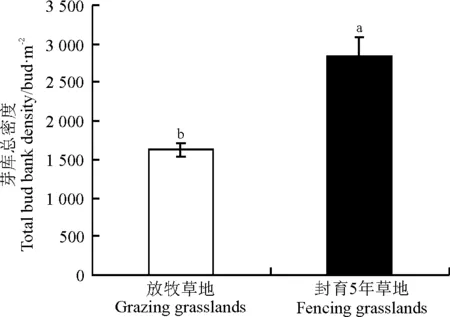
图1 封育和放牧对本氏针芽芽库总密度的影响Fig.1 Effects of grazing and feneing management on total bud bank of Stipa bungeana density
注:不同小写字母表示不同样地间差异显著(P<0.05)。
Note: Different lower case letters indicate significant difference between grazing and fencing grasslands at the 0.05 level.
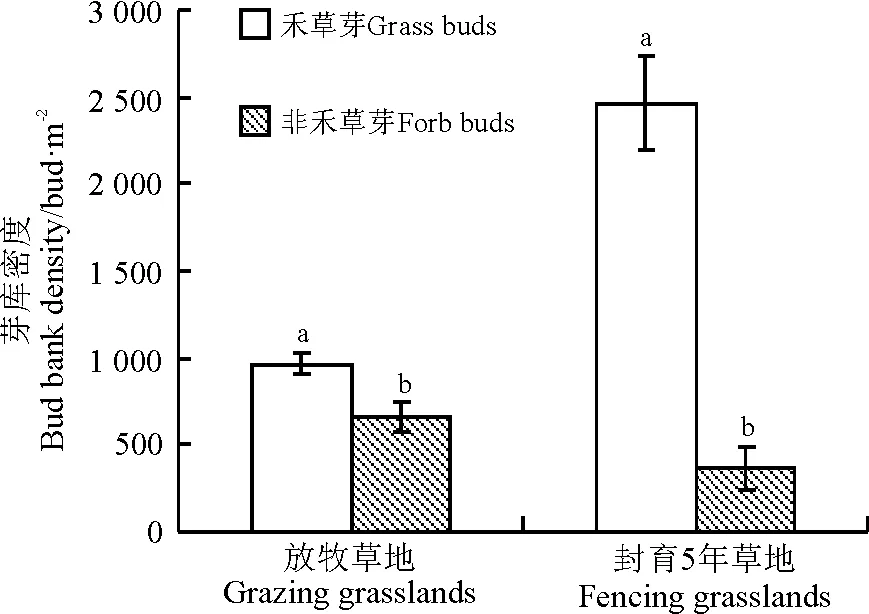
图2 封育和放牧对禾草和非禾草芽库密度的影响Fig.2 Effects of grazing and fencing management on the density of grass and forb bud bank
注:不同小写字母表示相同样地禾草芽与非禾草芽间差异显著(P<0.05)。
Note: Different lower case indicate significant difference between grass buds and forb buds in the same sample plot at the 0.05 level.
在黄土高原典型草原芽库中,分蘖芽占优势,其次为根颈芽和根茎芽,根蘖芽所占比例最小。封育和放牧草地间的根茎芽、分蘖芽和根蘖芽密度差异显著(P<0.05),而根颈芽密度差异不显著(P>0.05)。封育5年草地的根茎芽和分蘖芽密度均显著增加,而根蘖芽密度显著降低,根颈芽密度也降低,但差异不显著(P>0.05)(表2)。
2.3 芽库与地上植被的关系
用Pearson相关分析法分析芽库密度与地上植被的密度、地上茎分枝数和地上生物量的相关性,结果表明芽库总密度、非禾草芽库密度与植被的密度、地上茎分枝数和地上生物量之间相关性均不显著(P>0.05)。禾草芽库密度与地上生物量呈显著正相关(R2=0.223,P<0.05),即随着禾草芽密度的增加,植被的地上生物量增加(图3)。禾草芽库密度与地上茎分枝数呈显著负相关(R2=0.145 8,P<0.05),即随着禾草芽密度的增加,地上茎分枝数量减少(图4)。
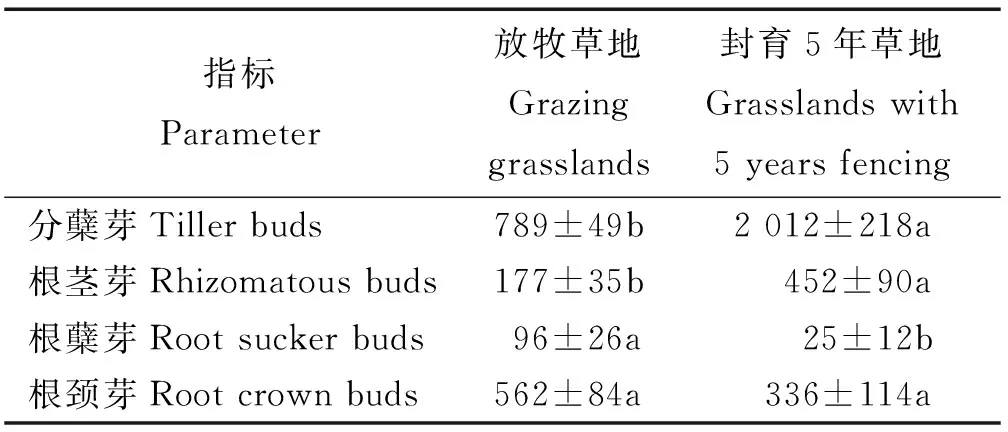
表2 封育和放牧对不同芽库类型密度(芽·m-2)的影响Table 2 Effects of grazing and fencing management on density(bud·m-2) of different bud bank types
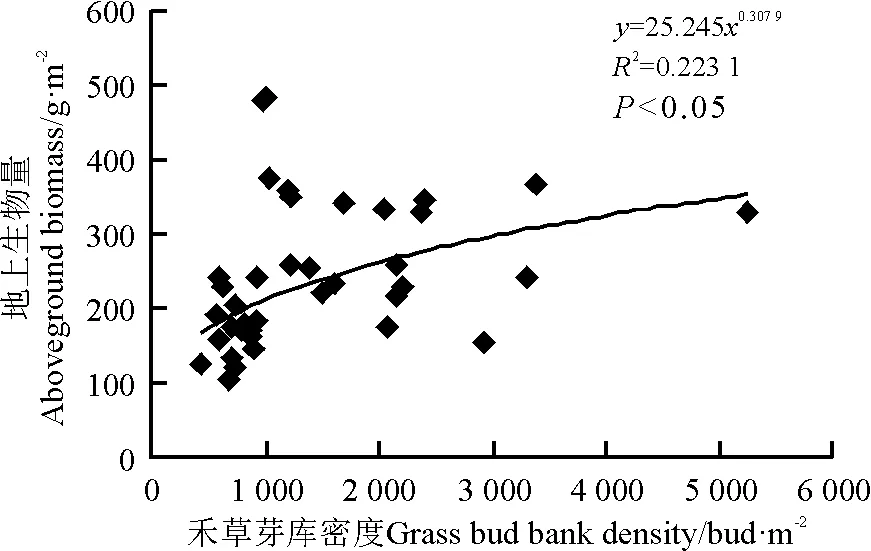
图3 禾草芽库密度与地上生物量的相关性Fig.3 Relationship between grass bud bank density and above-ground biomass
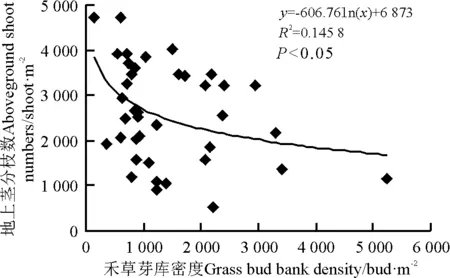
图4 禾草芽库密度与地上茎分枝数的相关性Fig.4 Relationship between grass bud bank density and above-ground shoot numbers
3 讨论与结论
芽库密度是芽库研究的重点问题,在恢复生态学上研究意义深远。研究者指出草地生态系统的芽库密度多为102~103芽·m-2[16]。与湿地生态系统相比[17],草地生态系统的种子密度相对较大。本研究得出黄土高原典型草原芽库密度大小为832~6 848芽·m-2,平均密度为 2 229芽·m-2,与黄土丘陵沟壑区土壤侵蚀环境下的芽库密度(600~1 300芽·m-2)[18]处于同一数量级水平。可见,黄土高原典型草原存在足够大小的芽库用于草地更新。采用变异系数对芽库的水平分布格局进行研究,结果表明短期封育提高了典型草原芽库的变异系数。芽库的水平分布格局主要受地上植被分布格局的影响。在放牧地,群落的物种多样性较高,均匀性较好,地上植被的空间异质性低于封育5年草地。盐池柳杨堡进行研究发现人工封育区的植被空间异质性高于围栏外[19]。与放牧地相比,封育5年样地保护了禾本科牧草免于动物的采食与践踏,已经形成了以本氏针茅和大针茅为优势种的群落,群落结构单一且不稳定。本氏针茅和大针茅为密集型植物,分蘖芽在形态上密集成块,因此封育5年草地芽库水平分布的空间异质性较高。
本研究表明,封育和放牧显著影响典型草原芽库密度。放牧对芽库密度的影响原因有两方面:一方面是由于动物采食和践踏根、茎、叶和芽等部位,直接降低牧草芽的存活率,另一方面是牧草被动物采食后,其再生主要依靠根和根颈贮藏的营养物质,从而进一步影响芽的输入、保存与输出。在该区,封育5年草地的芽库密度明显大于放牧地。可见,放牧增加了芽库的输出量,提高了地上分株数量。适度的动物取食,明显提高了秋龙胆(Gentianellaamarelle)芽的输出和地上分枝个数[20-22]。但放牧干扰对不同类型的芽库影响不同。本研究得出放牧明显提高了非禾草芽库密度,降低了禾草芽库密度。在北美草原的地下芽库中,放牧能显著影响禾草芽的输出,通过减少芽的密度而增加了地上分株密度[23],但放牧对垂穗草(Boutelouacurtipendula)的腋芽数量、地位、寿命和输出影响均不显著[24]。可见,放牧干扰如何影响芽库的密度还需进一步研究。
在本研究中,根据芽的着生位置将芽库分为4种类型:根茎型、分蘖型、根颈型和根蘖型。具有分蘖芽和根颈芽的集团型植物更多表现为侵占行为,称为“利用策略”,而具有根茎芽和根蘖芽的游击型植物则更多的表现为逃避行为,称为“占据策略”[25]。在频繁干扰的生境中,位于地下的根茎芽和根蘖芽更容易由休眠芽转化为活动芽而萌发,进而补充到地上种群更新中。本研究表明,封育和放牧管理措施对典型草原芽库密度的显著影响,主要来自于根茎型、分蘖型和根颈型牧草芽库的贡献。在以本氏针茅和大针茅为优势种的群落,分蘖芽为芽库的主要类型,对草地群落芽库密度影响较大。对分蘖型禾草来说,放牧可降低分蘖芽芽库密度和提高分蘖芽输出量,使牧草分蘖或营养枝条增加。对根茎型禾草来说,放牧可降低根茎芽密度,提高根茎芽的输出,促进根茎上枝条的萌蘖,使其产生更多的枝条。这在一定程度上可阐明放牧增加地上植被物种多样性和密度的机制。研究者指出放牧管理主要通过动物的采食与践踏来抑制牧草的垂直生长,从而增加根茎型禾草和丛生型禾草的分蘖数[26],这与本研究结果一致。
本研究表明,禾草芽库密度能很好预测多年生草地地上植被的动态变化。在该区,禾草芽库密度与地上生物量呈显著正相关,这与研究者[9]在北美高草原得出的结论一致,同时还得出禾草芽库大小是草原地上生物量很好的预测指标,地上生物量变化主要通过芽库数量来调控。另外,在封育5年草地,随着芽库密度的增加,地上茎分枝数显著下降,但地上生物量显著增加,说明短期封育提高地上生物量是通过增加单个枝条的生物量,即通过减少动物采食,提高牧草高度,这在一定程度上阐明了封育和放牧对地上植被动态变化的影响机制。
综上所述,芽库在黄土高原典型草原地上植被的更新与群落动态变化中发挥着重要作用。封育措施是提高芽库密度的有效措施之一,实施封育措施后,能加速退化草地的正向演替。在草地管理中,芽库对地上植被的发展有一定的指示作用,可以依靠芽库的测定来预测未来植被的产量、演替趋势和发展方向。
References:
[1] Clark-Tapia R,Alfonso-Corrado C,Eguiarte L E,Francisco M F.Clonal diversity and distribution inStenocereuseruca(Cactaceae),a narrow endemic cactus of the Sonoran Desert.American Journal of Botany,2005,92(2):272-278.
[2] Abernethy V J,Willby N J.Changes a long a disturbance gradient in the density and composition of propagule banks in flood plain aquatic habitats.Plant Ecology,1999,140(2):177-190.
[3] Liu W,Zhang Q,Liu G.Seed banks of a river-reservoir wetland system and their implications for vegetation development.Aquatic Botany,2009,90(4):7-12.
[4] Benson E J,Hartnett D C,Mann K H.Belowground bud banks and meristem limitation in tallgrass prairie plant populations.American Journal of Botany,2004,91(3):416-421.
[5] Klimešová J,Klimeš L.Bud banks and their role in vegetative regeneration——A literature review and proposal for simple classification and assessment.Perspectives in Plant Ecology,Evolution and Systematics,2007,8(3):115-129.
[6] Ott J P,Hartnett D C.Contrasting bud bank dynamics of two co-occurring grasses in tallgrass prairie:Implications for grassland dynamics.Plant Ecology,2012,213(9):1437-1448.
[7] Hartnett D C,Setshogo M P,Dalgleish H J.Bud banks of perennial savanna grasses in Botswana.African Journal of Ecology,2006,44:56-263.
[8] Dalgleish H J,Kula A R,Hartnett D C,Sandercock B K.Responses of two bunchgrass to nitrogen addition in tallgrass prairie:Tthe role of bud bank demography.American Journal of Botany,2008,95(6):672-680.
[9] Dalgleish H J,Hartnett D C.The effects of fire frequency and grazing on tallgrass prairie productivity and plant composition are mediated through bud bank demography.Plant Ecology,2009,201(2):411-420.
[10] 韩路,潘伯荣,王绍明,王建成.适度放牧对林间草原植物形态特征及生物多样性的影响.草业科学,2015,32(9):1405-1412. Han L,Pan B R,Wang S M,Wang J C.Effects of grazing on the morphological characteristics and biodiversity of the forest steppe in Yili.Pratacultural Science,2015,32(9):1405-1412.(in Chinese)
[11] 卢翔,南志标.放牧对草地植物群落多样性和草地病害的影响.草业科学,2015,32(9):1423-1431. Lu X,Nan Z B.Perspectives in effects of grazing on diversity of grassland plant community and forage diseases.Pratacultural Science,2015,32(9):1423-1431.(in Chinese)
[12] Wang N,Jiao J Y,Jia Y F,Bai W J, Zhang Z G.Germinable soil seed banks and the restoration potential of abandoned cropland on the Chinese hilly-gullied loess plateau.Environmental Management,2010,46(3):367-377.
[13] Zhao L P,Su J S,Wu G L,Gillet F.Long-term effects of grazing exclusion on aboveground and belowground plant species diversity in a steppe of the Loess Plateau,China.Plant Ecology and Evolution,2011,144(3):313-320.
[14] 胡安,陈皓,陈先江,侯扶江.黄土高原农田与草原土壤种子库.草业科学,2015,32(7):1035-1040. Hu A,Chen H,Chen X J,Hou F J.Soil seed banks of cropland and rangeland on the Loess Plateau.Pratacultural Science,2015,32(7):1035-1040.(in Chinese)
[15] 赵凌平,程积民,万惠娥,石绘陆,谭世图.黄土高原草地封育与放牧条件下土壤种子库特征.草业科学,2008,25(10):78-83 Zhao L P,Cheng J M,Wang H E,Shi H L,Tan S T.Characteristics of soil seed banks in grassland under fencing and grazing in Loess Plateau.Pratacultural Science,2008,25(10):78-83.(in Chinese)
[16] 赵凌平,王占彬,程积民.草地生态系统芽库研究进展.草业学报,2015,24(7):172-179. Zhao L P,Wang Z B,Cheng J M.Review of bud banks in grassland ecosystems.Acta Prataculture Sinica,2015,24(7):172-179.(in Chinese)
[17] 邓正苗.洞庭湖湿地典型优势植物地下芽库动态、萌发及其对环境变化的响应.长沙:中国科学院大学博士学位论文,2013. Deng Z M.The demography,sprouting and responses to environmental changes of belowground bud banks of dominant plants in the Dongting Lake werlands,China.PhD Thesis.Changsha:University of Chinese Academy of Sciences,2013.(in Chinese)
[18] 杜华栋,焦菊英,寇萌,王宁.黄土丘陵沟壑区土壤侵蚀环境下芽库的季节动态及垂直分布.应用生态学报,2013,24(5):1269-1276. Du H D,Jiao J Y,Kou M,Wang N.Seasonal dynamics and vertical distribution pattern of bud bank in different erosion environments on hilly-gully Loess Plateau of Northwest China.Chinese Journal of Applied Ecology,2013,24(5):1269-1276.(in Chinese)
[19] 王志述,张克斌,王晓.盐池柳杨堡人工封育区植被空间异质性.中国水土保持科学,2015,13(5):52-57. Wang Z S,Zhang K B,Wang X.Spatial heterogeneity of vegetation in artificially fenced area in Liuyangpu of Yanchi County in Ningxia.Science of Soil and Water Conservation,2015,13(5):52-57.(in Chinese)
[20] Huhta A P,Hellstrom K,Rautio P,Tuomi J.Grazing tolerance ofGentianellaamarellaand other monocarpic herbs:Why is tolerance highest at low damage levels.Plant Ecology,2003,166:49-61.
[21] Tolvanen A,Schroderus J,Henry G H R.Age and stage based bud demography ofSalixarcticaunder contrasting muskox grazing pressure in the High Arctic.Evolutionary Ecology,2001,15(4):443-462.
[22] Enright N J,Miller B P.Livestock grazing impacts on desert vegetation,Khirthar National Park,Pakistan.Rangeland Ecology and Management,2007,60(6):680-684.
[23] Dalgleish H J,Hartnett D C.Belowground bud banks increase along a precipitation gradient of the North American Great Plains:A test of the meristem limitation hypothesis.New Phytologist,2006,171(1):81-89.
[24] Hendrickson J R,Briske D D.Axillary bud banks of two semiarid perennial grasses:Occurrence,longevity,and contribution to population persistence.Oecologia,1997,110(4):584-591.
[25] 陈尚,王刚,李自珍.白三叶分枝格局研究.草业科学,1995,12(2):35-40. Chen S,Wang G,Li Z Z.Branching pattern of white clover.Pratacultural Science,1995,12(2):35-40.(in Chinese)
[26] 杨利民,韩梅,李建东.中国东北样带草地群落放牧干扰植物多样性的变化.植物生态学报,2001,25(1):l10-114. Yang L M,Han M,Li J D.Plant diversity change in grassland communities along a grazing disturbance in the northeast China transect.Acta Phytoecologica Sinica,2001,25(1):110-114.(in Chinese)
(责任编辑 苟燕妮)
Effects of fencing and grazing management on bud bank in a semiarid steppe on the Loess Plateau
Cui Wei1, Zhao Ling-ping1,2, Zhao Fu-rong1,2
(1.Animal Science and Technology School, Henan University of Science and Technology, Luoyang 471003, China;2.Forage Resources Exploration and Animal Health Cultivation Academician Work Station of Henan Province, Luoyang 471003, China)
In perennial grassland ecosystems, the bud bank plays an important role in vegetation maintenance and recruitment. In this study, we used field root-digging to examine the bud bank, analyze above-ground vegetation characteristics, and below-ground bud bank density, under fencing and grazing management measures. Results showed that the bud bank in a semiarid perennial steppe on the Loess Plateau was dominated by tiller buds, and bud bank density ranged from 832 to 6 848 bud·m-2. Short-term fencing significantly increased the bud bank density and the density of grass bud banks, and decreased the density of forb bud banks in relation to grazed grasslands (P<0.05). The change of tiller buds, rhizomatous buds and root sucker buds caused a change in total bud bank density under two management types. The density of tiller buds and rhizomatous buds was significantly higher and the density of root sucker buds was significantly lower in grasslands fenced for five years than in grazed grasslands (P<0.05). Grass bud bank density was significantly and positively correlated with above-ground biomass, but negatively correlated with above-ground shoot numbers (P<0.05). To a certain extent, the effects of grazing and fencing on aboveground vegetation in a semiarid steppe on the Loess Plateau are mediated through grass bud bank demography. The bud bank is an excellent predictor of the above-ground vegetation. We can predict grassland productivity and succession direction using bud bank determination.
fencing; bud bank; semiarid steppe; asexual recruitment
Zhao Fu-rong E-mail:zhaofurong66@163.com
10.11829/j.issn.1001-0629.2016-0151
2016-03-24 接受日期:2016-06-14
国家自然科学青年基金项目(31302013);河南省教育厅科学技术研究重点项目(13A2302812013)
崔伟(1975-),男,河南洛阳人,在读硕士生,主要从事草业科学研究。E-mail:t_s_t@sina.com
赵芙蓉(1964-),女,河南开封人,教授,硕导,博士,主要从事草业科学和动物营养研究。E-mail:zhaofurong66@163.com.
S812.8
A
1001-0629(2017)1-0009-07*
崔伟,赵凌平,赵芙蓉.封育和放牧对黄土高原典型草原芽库的影响.草业科学,2017,34(1):9-15.
Cui W,Zhao L P,Zhao F R.Effects of fencing and grazing management on bud bank in a semiarid steppe on the Loess Plateau.Pratacultural Science,2017,34(1):9-15.
