广东内伶仃岛白桂木群落结构及其演替研究
2017-02-09谭维政徐华林陈艺敏赵万义昝启杰廖文波
谭维政,徐华林,陈艺敏,赵万义,昝启杰,廖文波
(1 中山大学 生命科学学院/有害生物防治与资源利用国家重点实验室/广东省热带亚热带植物资源重点实验室,广东 广州 510275;2 广东内伶仃福田国家级自然保护区管理局,广东 深圳 518040)
广东内伶仃岛白桂木群落结构及其演替研究
谭维政1,徐华林2,陈艺敏1,赵万义1,昝启杰2,廖文波1
(1 中山大学 生命科学学院/有害生物防治与资源利用国家重点实验室/广东省热带亚热带植物资源重点实验室,广东 广州 510275;2 广东内伶仃福田国家级自然保护区管理局,广东 深圳 518040)
【目的】揭示内伶仃岛植被的演替,为中国特有珍稀濒危植物白桂木Artocarpushypargyreus种群的保护提供科学依据。【方法】运用群落生态学方法对内伶仃岛白桂木群落进行样地调查,分析群落的组成和结构、地理成分性质以及物种多样性等,并且与其他3个热带或中亚热带植物群落进行比较。【结果】该群落共有维管植物73种,隶属于40科61属,其中蕨类植物有4科4属4种,种子植物有36科57属69种。种类组成以热带分布属占绝对优势,占总属数达98.18%,具有南亚热带成分向热带成分过渡的性质。群落垂直结构较明显,乔木层优势种主要有翻白叶树Pterospermumheterophyllum、白桂木、假柿木姜子Litseamonopetala和浙江润楠Machiluschekiangensis等,其重要值依次为43.86、31.64、12.75和10.39;灌木层则以破布叶Microcospaniculata和九节Psychotriarubra为主;层间藤本植物亦十分发达。群落中主要优势种群的分布格局皆为聚群分布,年龄结构以II级(DBH<2.5 cm)和III级(2.5 cm≤DBH<7.5 cm)立木占绝大多数,属增长型种群,但白桂木种群为衰退型种群。群落的Simpson多样性指数(E)=0.95,Shannon-Wiener多样性指数(H)=3.27,Pielou均匀度指数(EH)=0.84。【结论】内伶仃岛白桂木群落是一个处于群落演替中期的南亚热带常绿阔叶林类型。
白桂木群落; 珍稀濒危种; 物种多样性; 内伶仃岛; 群落演替
白桂木Artocarpushypargyreus隶属于桑科Moraceae菠罗蜜属Artocarpus,为常绿乔木,中国特有种,其野生种群的数量日渐减少,大多呈单株散生分布,林下幼树、幼苗均少见,属衰退型种群[1]。2013年《中国生物多样性红色名录——高等植物卷》已将白桂木列入濒危等级[2],现主要零星分布于福建南部和西部、江西南部、湖南东南部和西南部、广东北部、海南、广西、云南东南部[3]。白桂木食用和药用价值颇高,果实和种子可生食,亦可做为蜜饯、饮料等的原料,其根入药,主治风湿痹痛、腰膝酸软等症[3]。树干高大,木材可作家具,乳汁可以提取硬性胶;树形优美,是园林绿化的优良树种。白桂木“浑身是宝”,所以国内对其研究报道也较多,主要集中在白桂木的化学成分[4]、药用有效部位的筛选和作用机制[5-6]、育种技术[7]、遗传多样性[8]等方面,另有诸多文献报道从白桂木种子抽提液中分离纯化得到的白桂木凝集素(Artocarpushypargyreuslectin,AHL)应用在免疫学方面的研究[9-12]。但是关于白桂木种群或群落生态学方面的报道却很少,仅在一些地方性科考报告中有所提及。1997—1999年编撰的关于内伶仃岛生态的综合性科考报告[13]中提到白桂木-刺葵群系的存在,但未进行样方调查及进一步的深入研究。
内伶仃岛自上世纪80年代被定为国家级自然保护区后,管理力度大大加强,岛上原来曾遭到严重破坏的植被得到逐步恢复,现以次生性的南亚热带常绿阔叶林为主,代表性的群落有红鳞蒲桃Syzygiumhancei-短序润楠Machilusbreviflora群落、潺槁木姜子Litseaglutinosa-短序润楠-假苹婆Sterculialanceolate群落、白桂木-翻白叶树Pterospermumheterophyllum群落等[14],其中白桂木群落分布在海拔150~250 m的岛中东部。本文运用群落学研究方法对该白桂木群落的结构组成、种类组成的地理成分、物种多样性等进行分析研究,进一步揭示内伶仃岛植被的演替,为白桂木种群的保护提供科学依据。
1 研究区域的自然地理概况
广东内伶仃岛位于珠江口的伶仃洋上,东距香港11.12 km,西距珠海24.08 km,北离深圳蛇口约16.70 km,地理位置为北纬 22°23′49′′~22°25′35′′,东经 113°46′18′′~113°49′49′′,西北至东南长约 4 km ,南北宽约 2 km ,总面积约 4.98 km2,为珠江口现存猕猴的4个岛中面积最小的岛屿,属南亚热带季风气候区[15]。该岛全年高温多雨,年均气温22.0~22.4 ℃,月均温10 ℃以上,有明显的干、湿季之分,降水集中在 4—9月。年均降雨2 000 mm左右,雨日约140 d,日照年总量约 2 000 h[14]。地质地貌性质主要为花岗岩、变质砂岩构成的海岛丘陵,东半部以花岗岩为主,西半部为变质砂岩。地带性土壤为赤红壤,还有海滨砂土和耕作土[16]。研究样地位于内伶仃岛尖峰山附近,海拔175 m,坐标:22°24′22.92″N,113°48′51.75″E;坡度10°左右。样地内乔木和灌丛较稀疏,层间藤本较大且密集,林冠层郁闭度为0.6~0.7;样地外沿为遭破坏的空地,地表大部分覆盖有五爪金龙Ipomoeacairica和薇甘菊Mikaniamicrantha等草质藤本。
2 研究方法
2.1 样地设置
设置样地1 100 m2,划分为11个10 m×10 m方格,采用单株每木记账调查法,起测径阶≥1.5 cm,高度≥1.5 m。记录样方内乔、灌木的种名、胸径、高度、冠幅;另在每个方格内设置1个2 m×2 m的小样方,记录样方中草本和乔、灌木幼苗,包括种名、高度、株数(丛数)和覆盖度等。
2.2 分析方法
根据野外调查所得的相关数据,对白桂木群落的种类组成、地理成分、年龄结构、主要种群分布格局、物种多样性等进行分析。
2.2.1 重要值 按照王伯荪等[17]方法计算群落中胸径≥1.5 cm的各种群的重要值(IV)。IV=相对显著度+相对频度+相对多度。
2.2.2 种群年龄结构 根据株高(h)及胸径(DBH),采用5级立木划分标准: Ⅰ级为苗木,h<33 cm;Ⅱ级为小树,h≥33 cm,DBH<2.5 cm;Ⅲ级为壮树,2.5 cm≤DBH<7.5 cm;Ⅳ级为大树,7.5 cm≤DBH<22.5 cm;Ⅴ级为老树,DBH≥22.5 cm[17]。
2.2.3 种群分布格局 主要种群分布格局分析采用聚集度指标进行测定,度量其分布的聚集程度(随机、均匀或聚集)[18]。具体指标计算公式如下:
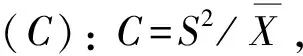
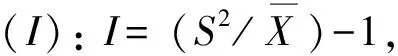
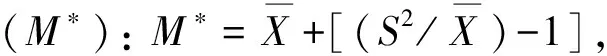
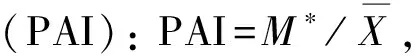
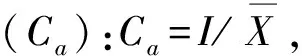
2.2.4 群落物种多样性 根据Simpson多样性指数(D)、Shannon-Wiener多样性指数(H),以及Pielou均匀度指数(EH)进行测度。D= 1-∑Pi2,H= -∑PilnPi, EH =H/ lnS,Pi=Ni/N, 式中,i为随机第i种,Ni为某物种的个体数,N为观察到的个体总数,S为样方内个体总数[19]。
3 结果与分析
3.1 群落组成和结构分析
3.1.1 群落种类组成 根据样方数据统计,该群落有维管植物40科61属73种。其中蕨类植物占4科4属4种,被子植物占36科57属69种(表1)。群落组成中,含4种以上的科有6个,分别为樟科(2属8种)、番荔枝科(2属4种)、大戟科(6属6种)、梧桐科(3属4种)、豆科(3属4种)、茜草科(4属4种),就内伶仃岛的植物群落而言,樟科和番荔枝科虽然其种数占世界区系比例不大,但在群落中占有较大的比例,属于内伶仃岛植物区系的表征科[16];含2种及以下的科有32个,占总科数比例为80.00%。群落中除白桂木外,还有浙江润楠Machiluschekiangensis、常绿臭椿Ailanthusfordii、翻白叶树等中国特有且近危的树种[2]。总体上,该群落组成丰富,多样性较高,属于较典型的南亚热带常绿阔叶林。
表1 内伶仃岛白桂木群落种类组成
Tab.1 Species composition ofArtocarpushypargyreuscommunity in Neilingding Island
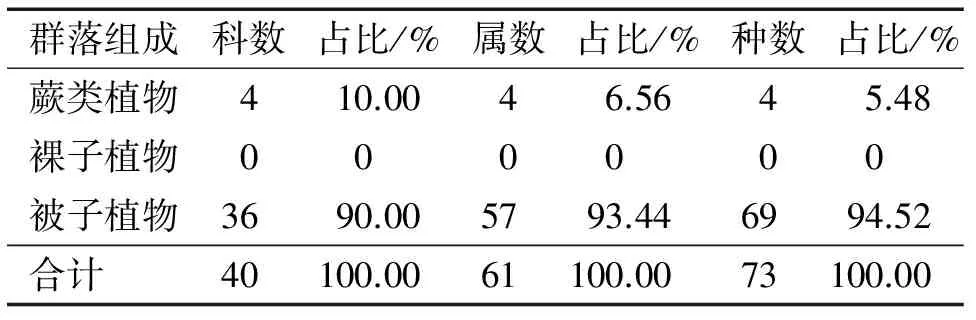
群落组成科数占比/%属数占比/%种数占比/%蕨类植物410.0046.5645.48裸子植物000000被子植物3690.005793.446994.52合计40100.0061100.0073100.00
3.1.2 地理成分分析 根据吴征镒[20-21]关于中国种子植物属的分布区类型划分原则,对内伶仃岛白桂木群落种子植物属的分布区类型进行统计。从表2中可以看出,该群落包含种子植物共57属,除去世界分布属,热带分布属占据绝对优势,共54属,占总属数达98.18%,且覆盖分布区类型的2~7项,其中泛热带分布属和热带亚洲分布属各16属,占比最大,均达29.09%,对群落的种类组成具有重要意义,如菠萝蜜属的白桂木、翅子树属Pterospermum的翻白叶树和润楠属Machilus的刨花润楠M.pauhoi是群落的建群种,破布叶属Microcos的破布叶Mi.paniculata是群落中主要的中层植物,而层间藤本植物刺果藤属Byttneria的刺果藤B.grandifolia更是在群落中密集分布,占据着相当大的空间。除热带分布属外,仅有柳叶菜属Epilobium属于北温带分布,占比1.82%,其余9~15项均无分布。群落中3种以下属共54属,其中单种属有50属,占比达92.60%,可以看出内伶仃岛地理成分的强烈扩散性特征。
表2 内伶仃岛白桂木群落种子植物属的分布区类型
Tab.2 The generic areal-types of seed plants inArtocarpushypargyreuscommunity in Neilingding Island
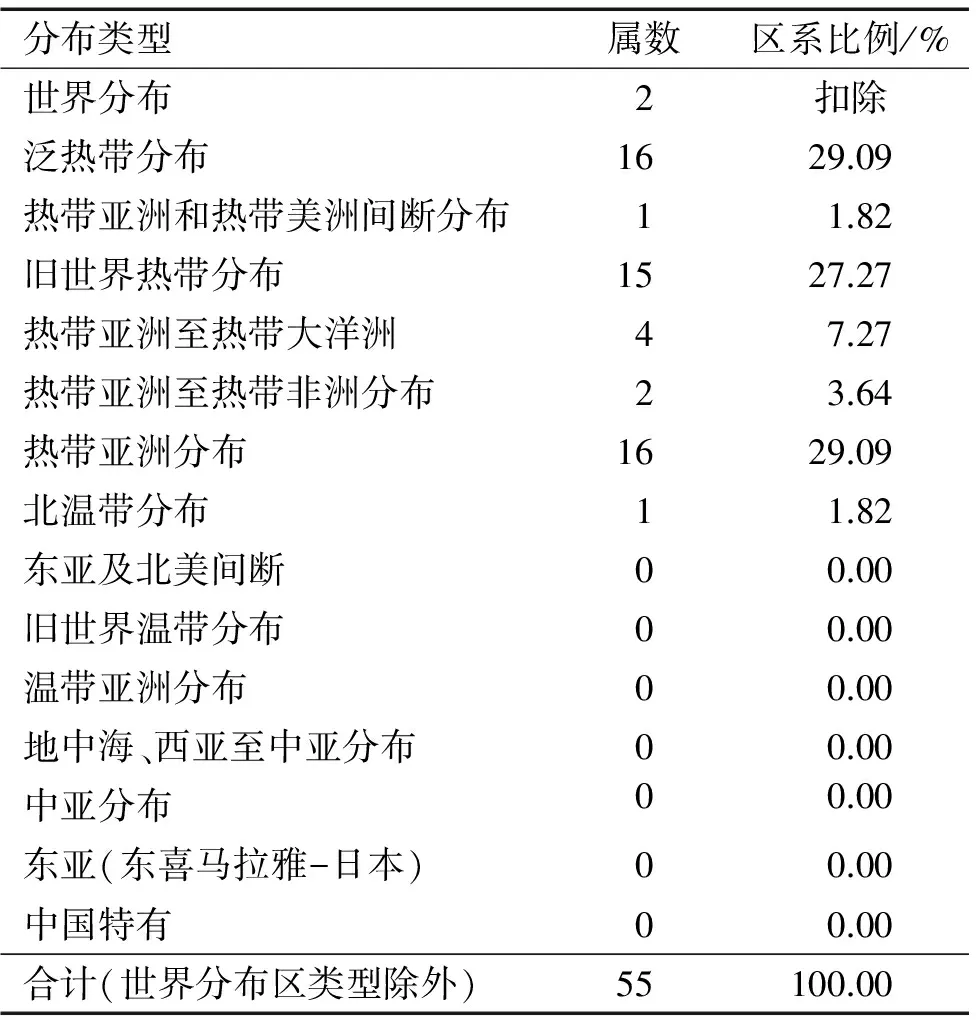
分布类型属数区系比例/%世界分布2扣除泛热带分布1629.09热带亚洲和热带美洲间断分布11.82旧世界热带分布1527.27热带亚洲至热带大洋洲47.27热带亚洲至热带非洲分布23.64热带亚洲分布1629.09北温带分布11.82东亚及北美间断00.00旧世界温带分布00.00温带亚洲分布00.00地中海、西亚至中亚分布00.00中亚分布00.00东亚(东喜马拉雅-日本)00.00中国特有00.00合计(世界分布区类型除外)55100.00
总体来看,该群落中热带成分明显占优势,并且白桂木等优势种具有一定的板根结构以及附生植物,出现一些岛屿和热带林特征,同时也具有南亚热带成分向热带成分过渡的性质。
3.1.3 群落结构分析 森林群落的垂直空间结构以群落的层次结构来反映,而森林群落的层次结构可以通过群落的垂直景观层次,即群落中林木的高度级分布来表征[22]。内伶仃岛白桂木群落内树种高度极差较大,最高达18 m,乔木各亚层区分比较明显,且层间大型藤本植物表现出空间渗透和镶嵌分布的现象。
根据群落的高度组成,按每隔2 m划分成若干个等级,第1高度级为0~2 m,第2高度级为2~4 m,依次类推。乔木层植物共有28种,按照不同高度级乔木树种在群落中的分布,乔木层可以分为2个亚层:第1亚层高度大于10 m(5级),平均高度13.4 m,分布有白桂木、刨花润楠等8种乔木,共33株,其中1株刨花润楠高度达18 m,也是群落中的最高树木,胸围188 cm;第2亚层高5~10 m,平均高度6.42 m,有白桂木、翻白叶树、假柿木姜子Litseamonopetala、浙江润楠等共28种,125株。白桂木的个体数在乔木第1亚层和第2亚层分别占24.24%和4.00%,翻白叶树和刨花润楠在群落中亦有较多分布,均占有一定优势。
灌木层以光叶紫玉盘Uvariaboniana、破布叶、香港大沙叶Pavettahongkongensis、九节Psychotriarubra为主,另有自然繁衍的、数量较少的乔木层幼苗,如白桂木、假苹婆、亮叶猴耳环AbaremaLucida等。该层植物种类丰富(共46种),植株数量较多(410株),占群落总个体数的72.18%;其中,九节数量最丰富,共有61株,占该层总个体数的14.87%,九节属于阴生性植物,常见于遭到破坏处于恢复阶段的热带亚热带次生林中。由此可以看出,该群落一方面优势种较集中,白桂木等明显较高,林分处于成熟林阶段,但另一方面林下阴生植物丰富,说明群落仍处于恢复发展演替期,尚未达到顶级或亚顶极状态。
草本层植物约50种,以三叉蕨Tectariasubtriphylla、半边旗Pterissemipinnata、海芋Alocasiacucullata以及禾本科植物为主,也包括一些乔木、灌木和藤本的幼苗,如浙江润楠、常绿臭椿、龙须藤Bauhiniachampionii等,体现了较高的物种丰富度。层间植物种类相对较多,达15种,且藤体相当发达,如刺果藤、白藤Calamustetradactylus占据了很大的空间。灌草层则以刺果藤、龙须藤为主,另有青江藤Celastrushindsii、柠檬清风藤Sabialimoniacea和香花崖豆藤Millettiadielsiana呈零星分布,体现出群落所在区域良好的水热条件,具有明显的热带型特征。
3.1.4 优势种群和重要值分析 对群落乔木层的重要值进行计算,表3列出了乔木层中重要值大于3.00的物种共22种。其中,翻白叶树和白桂木的重要值最大,分别为43.86和31.64,显然为该群落的建群种;破布叶在内伶仃岛上以较粗大的乔木为主,且分布众多,和常绿臭椿、假柿木姜子、亮叶猴耳环一起为群落乔木层的优势种,分列重要值排名第3、7、8、9位;九节、光叶紫玉盘在群落中个体数很多,刺果藤则藤体粗大,相对显著度高,故重要值排名都相对靠前,分列第4、5、6位。上述结果反映出了群落的结构特征和各种群空间分布的特点。
3.2 主要种群分布格局分析
群落中植物的水平空间分布格局是物种与环境长期相互适应、相互作用的结果,它不仅决定于物种自身的形态结构、生理生态特性,同时也与自然生境条件(包括土壤、地形、地貌等)及其所处群落中其他种群的竞争排斥等生态效应密切相关[23]。本文主要选取群落中占据优势地位(重要值大于10)的7种乔木,分析其空间分布格局。从表4可以看出,7种乔木种群的扩散系数C均大于1,为聚集分布;聚集度指数I均大于0,为聚集分布;平均拥挤度M*均大于各自的种群多度平均值,为聚集分布;聚块性指数PAI均大于1,为聚集分布;聚集指数Ca均大于0,为聚集分布;负二项分布指数K均大于0且小于8,为聚集分布。6个聚集度指标皆显示这7个种群的分布格局都服从集群分布,繁殖体散播的局限性和种群内竞争是内因,而岛上生存环境的一度破坏,如曾经的水土流失,大量的人类活动等是重要的外部因素。白桂木属于典型的聚集分布的物种,对生境要求较为严格,一般只生长在较为阴湿、富含有机质的下坡或山谷溪边,异质性的自然生境限制了白桂木的分布,使其只能在适宜的生境片断聚集生长[3],加之上述内、外部因素,导致了白桂木的濒危现状。
表3 内伶仃岛白桂木群落主要物种的重要值1)
Tab.3 Important values (IV) of dominant populations inArtocarpushypargyreuscommunity in Neilingding Island
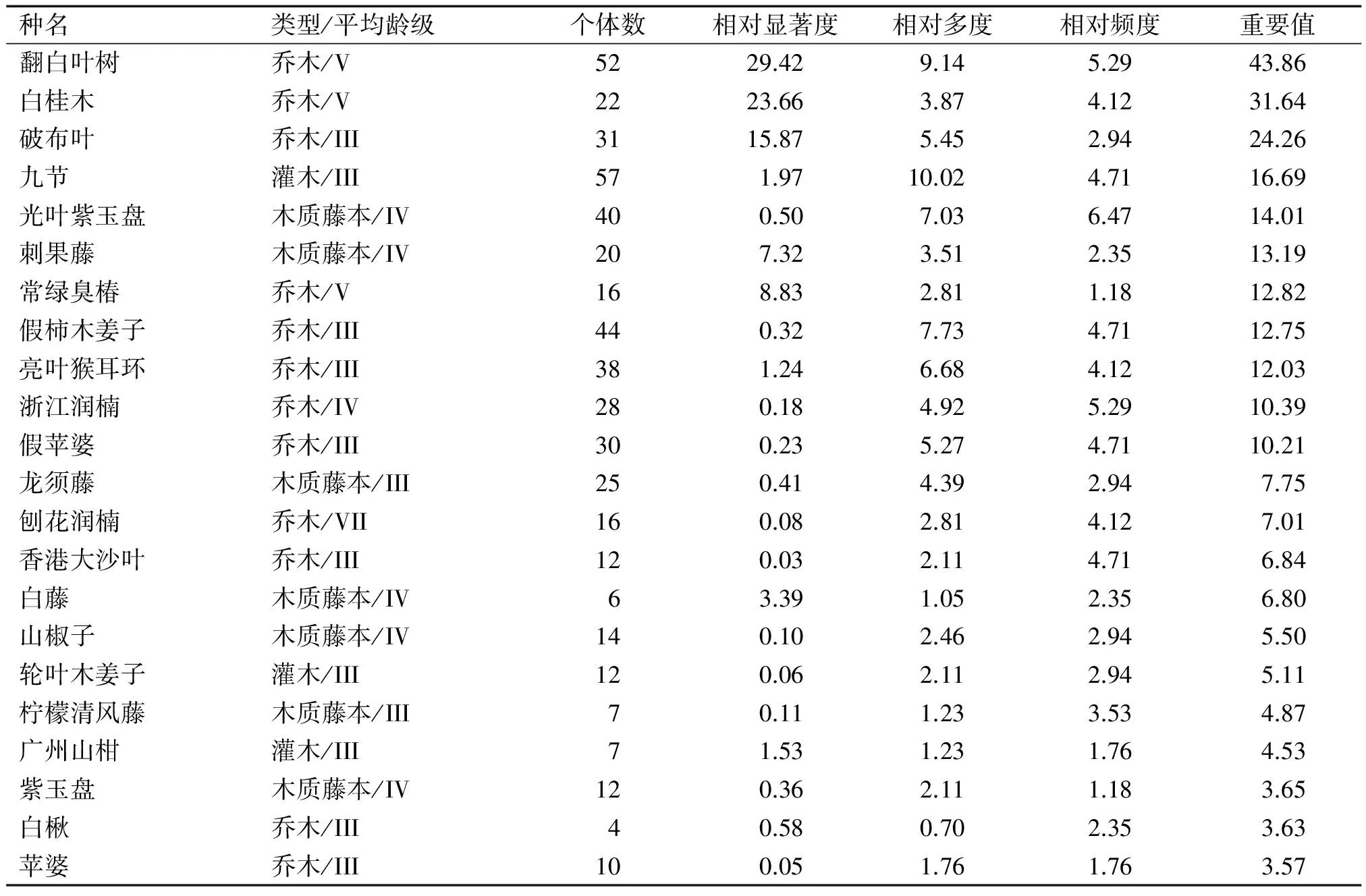
种名类型/平均龄级个体数相对显著度相对多度相对频度重要值翻白叶树乔木/V5229.429.145.2943.86白桂木乔木/V2223.663.874.1231.64破布叶乔木/III3115.875.452.9424.26九节灌木/III571.9710.024.7116.69光叶紫玉盘木质藤本/IV400.507.036.4714.01刺果藤木质藤本/IV207.323.512.3513.19常绿臭椿乔木/V168.832.811.1812.82假柿木姜子乔木/III440.327.734.7112.75亮叶猴耳环乔木/III381.246.684.1212.03浙江润楠乔木/IV280.184.925.2910.39假苹婆乔木/III300.235.274.7110.21龙须藤木质藤本/III250.414.392.947.75刨花润楠乔木/VII160.082.814.127.01香港大沙叶乔木/III120.032.114.716.84白藤木质藤本/IV63.391.052.356.80山椒子木质藤本/IV140.102.462.945.50轮叶木姜子灌木/III120.062.112.945.11柠檬清风藤木质藤本/III70.111.233.534.87广州山柑灌木/III71.531.231.764.53紫玉盘木质藤本/IV120.362.111.183.65白楸乔木/III40.580.702.353.63苹婆乔木/III100.051.761.763.57
1)表中为重要值大于3.00的种群,共计22种。
表4 白桂木群落7个主要种群的分布格局
Tab.4 Distribution patterns of seven dominant populations inArtocarpushypargyreuscommunity
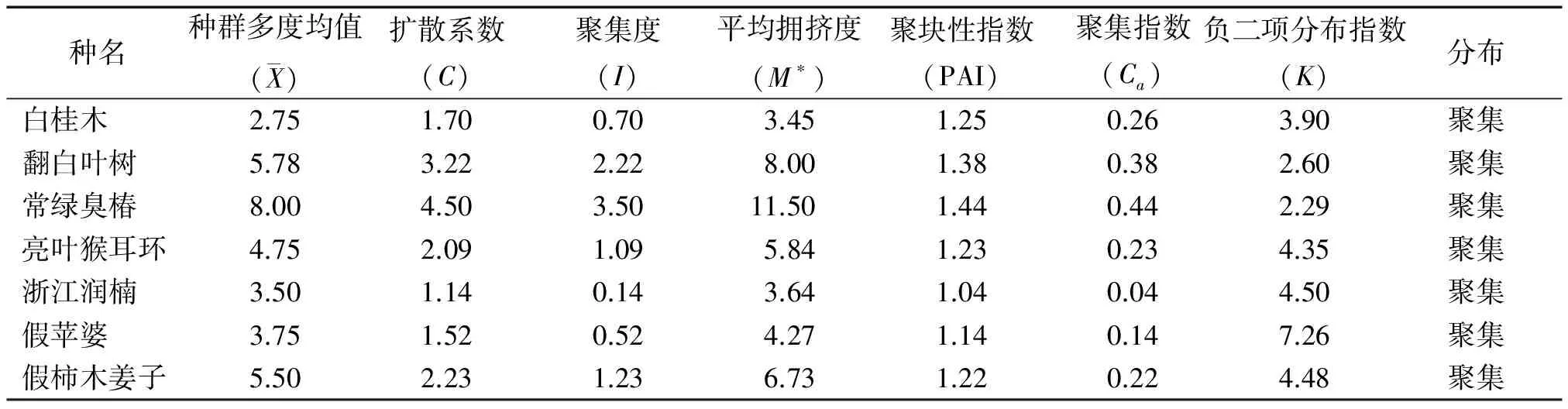
种名种群多度均值( X)扩散系数(C)聚集度(I)平均拥挤度(M∗)聚块性指数(PAI)聚集指数(Ca)负二项分布指数(K)分布白桂木2.751.700.703.451.250.263.90聚集翻白叶树5.783.222.228.001.380.382.60聚集常绿臭椿8.004.503.5011.501.440.442.29聚集亮叶猴耳环4.752.091.095.841.230.234.35聚集浙江润楠3.501.140.143.641.040.044.50聚集假苹婆3.751.520.524.271.140.147.26聚集假柿木姜子5.502.231.236.731.220.224.48聚集
3.3 乔木层优势种群的年龄结构分析
在内伶仃岛白桂木群落中选取乔木层中重要值较大的浙江润楠、亮叶猴耳环、假柿木姜子、假苹婆、常绿臭椿、翻白叶树和白桂木7个优势种群进行年龄结构分析(图1)。由于森林树种幼苗生存率极低,仅靠幼苗多寡难以对种群的未来做出预测,因此分析年龄结构中不包括幼苗[24]。
群落中浙江润楠、假柿木姜子和假苹婆3个种群的II级立木的占比均极高,其中假柿木姜子甚至达到84%,III级立木亦占有一定比例,由此说明这3个种群的幼态性和发展性具有很大潜力,有可能最终成为建群种。亮叶猴耳环种群III级立木最多,其次为II级小树,属于增长种群。常绿臭椿和翻白叶树的年龄结构相似,各级立木均占有一定比例,分布较均匀,且III级为最高,整体表现出一种平衡状态,具有稳定性,预测在群落中将持续保持其优势地位。而白桂木种群,V级老树最多,另有一定比例的III级和IV级立木,虽然现在在群落中优势地位明显,但是缺乏小树,意味着缺少新的补充,预测在以后群落的自然演替过程中将逐渐衰退,被其他增长型种群取代。
总体来看,该群落乔木层处于增长状态,并逐步向稳定状态过渡,说明岛上植被恢复状况良好,但是白桂木等珍稀濒危物种的生存和发展状况不太乐观。
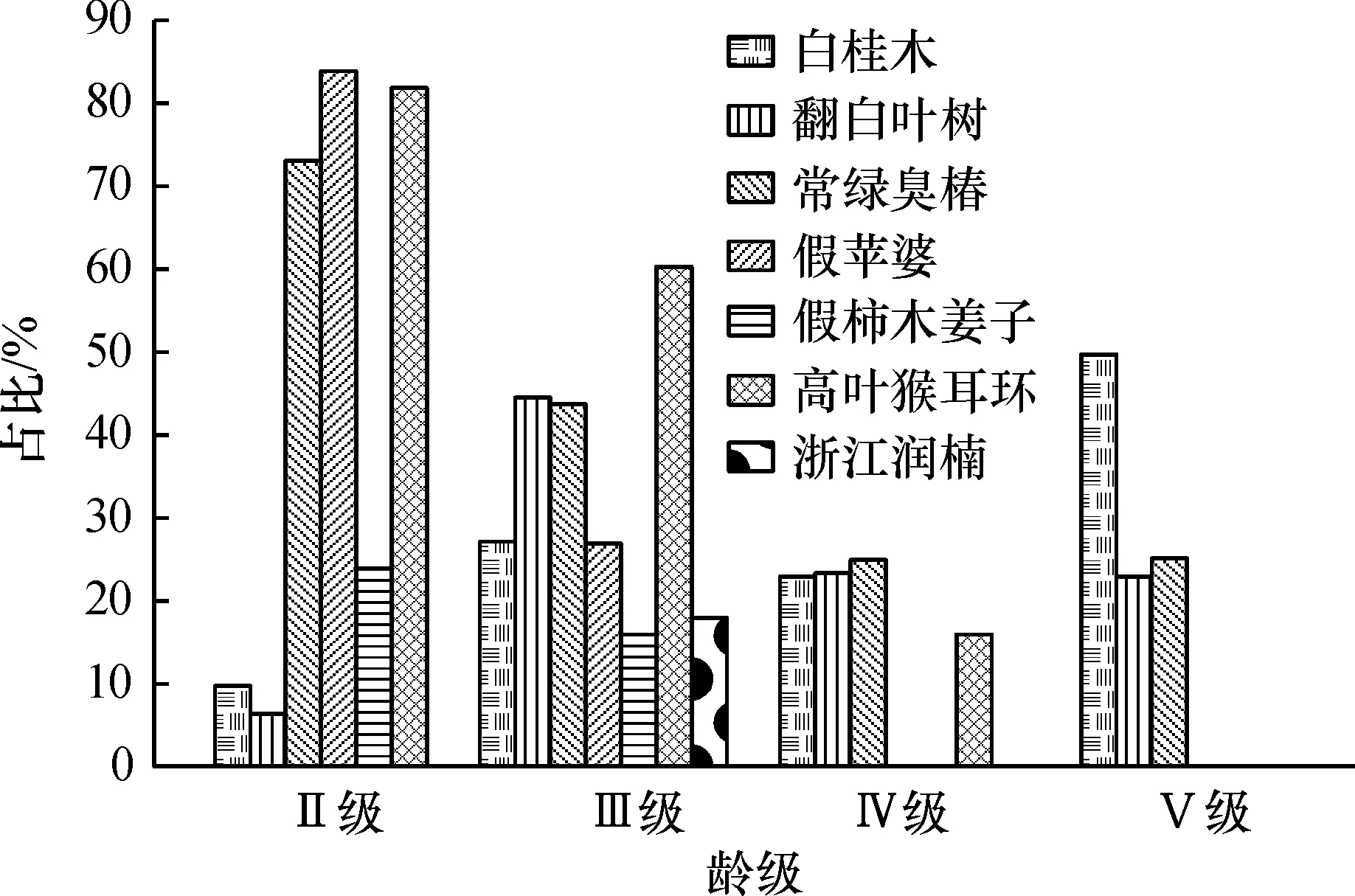
图1 内伶仃岛白桂木群落乔木层主要优势种年龄结构
Fig.1 Age structure of dominant populations in tree layer ofArtocarpushypargyreuscommunity in Neilingding Island
3.4 白桂木群落物种多样性分析
物种多样性能够表征生物群落和生态系统的结构复杂性,体现群落的结构类型、组织水平、发展阶段、稳定程度和生境差异,具有重要的生态学意义[25]。本文通过计算应用最为广泛的2个多样性指数(Simpson指数和Shannon-Wiener指数)对白桂木群落的物种多样性进行描述,但研究表明:Simpson指数更适于表征草本群落多样性[26],故此处重点讨论Shannon-Wiener指数并计算Pielou均匀度,并与典型的南亚热带常绿阔叶林:香港岛黄桐Endospermumchinense森林群落[27]和珠海淇澳岛肉实树Sarcospermalaurinum群落[28]以及1998年内伶仃岛上的布渣叶+小叶榕Ticusmicrocarpa-黄牛木Cratoxylumcochinchinense群落[29]进行比较(表5)。从表5中可以看出,4个群落均匀度指数接近,说明分布较均匀;白桂木群落的Shannon-Wiener指数低于黄桐森林群落和肉实树群落,但是高于1998年内伶仃岛黄牛木群落。从资料中可知,该黄桐森林群落是香港地区植被的顶极群落,肉实树群落处在演替阶段的中期到顶极群落之间,而黄牛木群落则处于演替初期到中期之间。相比较之下,白桂木群落物种多样性稍低,可见尚未达到与前面2个群落一致的发展阶段,但高于1998年内伶仃岛黄牛木群落,说明该群落处于演替中期,总体比较稳定。
表5 内伶仃岛及邻近地区几个南亚热带常绿阔叶林群落指数比较
Tab.5 Comparison of indexes of south subtropical evergreen broad-leaved forest communities in Neilingding Island and neighboring regions
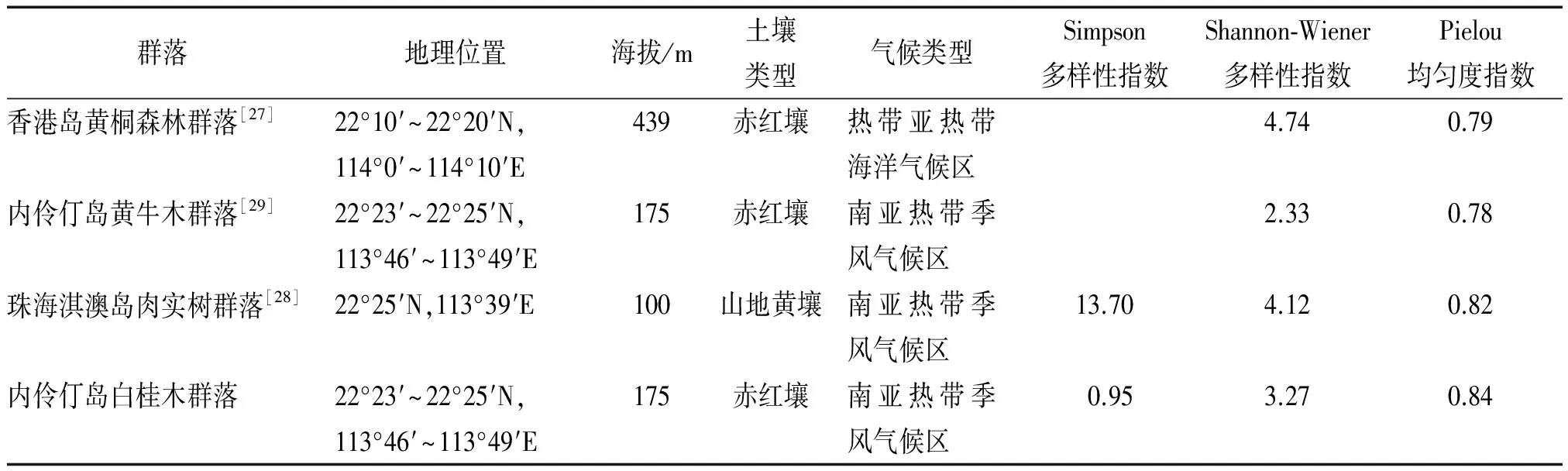
群落地理位置海拔/m土壤类型气候类型Simpson多样性指数Shannon⁃Wiener多样性指数Pielou均匀度指数香港岛黄桐森林群落[27]22°10′~22°20′N,114°0′~114°10′E439赤红壤热带亚热带海洋气候区4.740.79内伶仃岛黄牛木群落[29]22°23′~22°25′N,113°46′~113°49′E175赤红壤南亚热带季风气候区2.330.78珠海淇澳岛肉实树群落[28]22°25′N,113°39′E100山地黄壤南亚热带季风气候区13.704.120.82内伶仃岛白桂木群落22°23′~22°25′N,113°46′~113°49′E175赤红壤南亚热带季风气候区0.953.270.84
4 讨论与结论
从组成结构和地理成分看,内伶仃岛白桂木群落属于南亚热带常绿阔叶林类型,它既有亚热带常绿阔叶林和热带雨林的特征,又能与二者明显区分。首先,该群落热带区系成分明显,大戟科Euphorbiaceae和樟科Lauraceae等热带性较强科属的植物在群落中占据很大优势,林中木质藤本植物发达,空间占有率相当高;其次,主要种群的分布格局近似于其他典型的南亚热带常绿阔叶林类型,与热带雨林亦有相似性[27],区别于亚热带常绿阔叶林。然而,该群落在物种多样性上远低于热带雨林,优势种数量较中亚热带丰富,但在群落中未发现有气生根、茎花、附生的巢蕨、星蕨、兰花等特征性热带雨林现象,这些又与典型的热带雨林有较大不同。
从群落演替的阶段来划分,内伶仃岛白桂木群落处于演替发展的中期。优势种群整体以III级(2.5 cm≤DBH<7.5 cm)立木数量占优,II级(DBH≤2.5 cm)亦占有相当比例,说明群落处在快速发展时期。预测未来该群落的物种多样性将进一步提高,组成成分趋于复杂,群落内优势种或者建群种可能出现更替,直至过渡到稳定状态。
作为珍稀濒危物种的白桂木,在群落中发展的前景不容乐观。从种群内部分析,老树占绝大多数,幼树很少,种子散播范围有限,种子萌发率较低[3],因对生境要求较高从而形成的种群地理分布片段化、群落内部聚集化的分布方式,使得不同白桂木种群间的基因交流少,小种群分化严重,种群内杂合度低,导致适应性弱[8];从外部因素分析,其他优势种群的种间竞争,以及大量的人类活动等因素都是导致白桂木种群在该群落演替中处于发展劣势的原因。为了对白桂木种群进行有效的保护,可以从保护遗传学的角度出发,开发高效的分子标记进行遗传多样性方面的研究,然后在遗传多样性较高的种群内采种,并采用扦插和组织培养等快繁途径来扩大和更新现有种群[8]。同时施加以适当的辅助措施,如在内伶仃岛上进行人工播种抚育,扩大种群规模;将一些尚不耐荫蔽的幼树迁出林外建立适当的苗圃基地加以抚育保护;减少人类活动,严禁砍伐,同时适当加强宣传教育,提高人们的保护意识。
致谢:中山大学2012级本科生罗梦诗,硕士研究生袁天天、迟盛南、刘宇等参加了野外调查,特此鸣谢!
[1] 黎国运, 徐佩玲, 陈光群. 濒危植物白桂木组培育苗技术研究[J]. 热带林业, 2011, 39(3): 24-29.
[2] 中华人民共和国环境保护部,中国科学院. 中国生物多样性红色名录:高等植物卷[EB/OL].[2016-02-08].http://www.zhb.gov.cn/gkml/hbb/bgg/201309/W020130917614244055331.pdf.
[3] 范繁荣. 濒危植物白桂木的濒危机制与迁地保育研究[D]. 福州: 福建农林大学, 2008.
[4] 陈黎明, 谢平, 肖庆青, 等. 白桂木化学成分研究[J]. 中草药, 2007, 38(6): 815-818.
[5] 欧阳胜, 申作洁, 潘琳娜. 白桂木抗炎镇痛作用有效部位筛选[J]. 中草药, 2010, 41(11): 1850-1853.
[6] 申作洁, 乔玉丹, 欧阳胜, 等. 白桂木抗类风湿性关节炎的有效部位筛选及其作用机制研究[J]. 中草药, 2011, 42(9): 1792-1795.
[7] 黎国运, 徐佩玲, 陈光群. 濒危植物白桂木种子育苗技术研究[J]. 热带林业, 2010, 38(3): 23-24.
[8] 范繁荣. 濒危植物白桂木的遗传多样性研究[J]. 浙江农林大学学报, 2010, 27(2): 266-271.
[9] 周德义, 杨尔滨, 邓勇, 等. 白桂木凝集素的纯化与性质的研究[J]. 生物化学与生物物理学报, 1995, 27(1): 61-66.
[10]周素芳, 周德义. 两种亲和层析法纯化白桂木凝集素的比较[J]. 广西医科大学学报, 1997, 14(3):17-19.
[11]邓家珍, 何志义, 梁国容, 等. 白桂木凝集素-HPR夹心法在过敏性哮喘患者唾液SIgA含量检测中的应用[J]. 广西医科大学学报, 2001, 18(2): 226-227.
[12]WU Y S, ZHANG H, ZHOU S F,et al. Characterization ofArtocarpushypargyreuslectin interacting with glycoproteins [J]. Chin J Biochem Mol Biol, 2000, 16(2): 210-214.
[13]蓝崇钰, 王勇军. 广东内伶仃岛自然资源与生态研究[M]. 北京: 中国林业出版社, 2001: 37.
[14]蓝崇钰, 廖文波, 王勇军. 广东内伶仃岛的生物资源及自然保护规划[J]. 植物资源与环境学报, 2002, 11(1): 47-52.
[15]崔大方, 廖文波, 昝启杰, 等. 广东内伶仃岛国家级自然保护区的植物资源[J]. 华南农业大学学报, 2000, 21(3): 48-52.
[16]昝启杰, 廖文波, 陈继敏, 等. 广东内伶仃岛植物区系的研究[J]. 西北植物学报, 2001, 21(3): 507-519.
[17]王伯荪, 余世孝, 彭少麟, 等. 植物群落学实验手册[M]. 广州: 广东高等教育出版社, 1996: 1-56.
[18]洪伟. 闽江流域森林生态研究[M]. 厦门: 厦门大学出版社, 2000: 176-183.
[19]孙儒泳, 李博, 诸葛阳, 等. 普通生态学[M]. 北京: 高等教育出版社, 1996: 136-137.
[20]吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊): 1-179.
[21]吴征镒. 中国种子植物属的分布区类型的增订和勘误[J]. 云南植物研究, 1993 (增刊Ⅳ): 141-178.
[22]彭少麟, 方炜. 广州白云山次生常绿阔叶林的群落结构动态[J]. 应用与环境生物学报, 1996, 2(1):22-28.
[23]谢宗强, 陈伟烈, 路鹏, 等. 濒危植物银杉的种群统计与年龄结构[J]. 生态学报, 1999, 19(4): 523-528.
[24]王伯荪, 余世孝, 彭少麟, 等. 植物种群学[M]. 广州: 广东高等教育出版社, 1995: 8-15.
[25]许晴, 张放, 许中旗, 等. Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”[J]. 草业科学, 2011, 28(4): 527-531.
[26]吴昊. 不同类型群落物种多样性指数的比较研究[J]. 中南林业科技大学学报, 2015, 35(5): 84-89.
[27]王伯荪, 陆阳, 张宏达, 等. 香港岛黄桐森林群落分析[J]. 植物生态学与地植物学学报, 1987, 11(4): 241-251.
[28]田广红, 丁明艳, 杨雄邦, 等. 珠海市淇澳岛肉实树群落及其物种多样性特征[J]. 植物科学学报, 2013, 31(5): 461-466.
[29]袁天天, 赵万义, 徐华林, 等. 广东内伶仃岛马尾松群落和布渣叶群落的演替动态[J]. 广东林业科技, 2015, 31(1): 49-55.
【责任编辑 李晓卉】
Structure and succession of Artocarpus hypargyreus community in Neilingding Island, Guangdong
TAN Weizheng1, XU Hualin2, CHEN Yimin1, ZHAO Wanyi1, ZAN Qijie2, LIAO Wenbo1
(1 School of Life Sciences, Sun Yat-Sen University/State Key Laboratory of Biocontrol/Guangdong Provincial Key Laboratory of Plant
Resources, Guangzhou 510275, China; 2 Futian-Neilingding National Nature Reserves of Guangdong, Shenzhen 518040, China)
【Objective】 To reveal the succession of vegetation in Neilingding Island, and provide a scientific basis for protection ofArtocarpushypargyreus, a rare and endangered species endemic to China. 【Method】Method in community ecology was used to survey the sample plots ofA.hypargyreuscommunity in Neilingding Island, Guangdong. The community composition and structure, characteristics of geographical components and species diversity were analyzed, and were compared to those of other three plant communities in tropic or mid-subtropical regions.【Result】A total of 73 species of vascular plants belonging to 61 genera of 40 families were discovered in theA.hypargyreuscommunity.Among them, 4 species of pteridophyta belong to 4 genera of 6 families, and 69 species of seed plants belong to 57 genera of 36 families. Tropical genera (98.18%) occupied a foremost position in this community which shows a transitional property that range from south subtropical to tropical regions. The stratification of the community was obvious: the dominant populations in the arborous layer mainly werePterospermumheterophyllum,A.hypargyreus,LitseamonopetalaandMachiluschekiangensis, with the important values of 43.86, 31.64, 12.75 and 10.39 respectively, the dominant population in the shrub layer mainly wereMicrocospaniculataandPsychotriarubra, and the lianas were abundant among the layers. The dominant populations in this community had clustering distribution pattern, and were characterized by stumpage of level II (DBH<2.5 cm) and level III (2.5 cm≤DBH<7.5 cm), indicating that they belonged to the increasing population except theA.hypargyreuspopulation was declining. In this community, Simpson diversity index (E) was 0.95, Shannon-Wiener diversity index (H) was 3.27, and Pielou evenness index (EH) was 0.84. 【Conclusion】TheA.hypargyreuscommunity in Neilingding Island is a south subtropical evergreen broad-leaved forest at the middle stage of succession.
Artocarpushypargyreuscommunity; rare and endangered species; species diversity; Neilingding Island; community succession
2016- 03- 22优先出版时间:2017-01-10
谭维政(1992—),男,硕士研究生,E-mail: tan593326892@163.com;通信作者:廖文波(1963—),男,教授,博士,E-mail: lsslwb@mail.sysu.edu.cn; 昝启杰(1968—),男,教授级高工,博士,E-mail:zqjmangrove@126.com
深圳市财政局《深圳市珍稀重点野生植物调查》项目(71020106);深圳市级自然保护区生态环境监测项目(71021028);广东省野生动植物保护管理项目(42090404);2014—2015年度广东省本科教学质量和教学改革项目(18822504,18822506)
S718
A
1001- 411X(2017)02- 0099- 07
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20170110.1424.034.html
谭维政,徐华林,陈艺敏, 等.广东内伶仃岛白桂木群落结构及其演替研究[J].华南农业大学学报,2017,38(2):99- 105.
