淠河流域河源溪流鱼类空间分布格局及主要影响因素
2017-01-20张晓可王慧丽方云祥刘志刚郑爱芳陈敏敏于道平
张晓可,王慧丽,万 安,方云祥,刘志刚,郑爱芳,陈敏敏,于道平**
(1:安庆师范大学水生生物保护与水生态修复安徽省高校工程技术研究中心,安庆 246011)(2:安徽省环境科学研究院,合肥 230061)
淠河流域河源溪流鱼类空间分布格局及主要影响因素
张晓可1,王慧丽1,万 安1,方云祥2,刘志刚1,郑爱芳1,陈敏敏1,于道平1**
(1:安庆师范大学水生生物保护与水生态修复安徽省高校工程技术研究中心,安庆 246011)(2:安徽省环境科学研究院,合肥 230061)
淠河是安徽省内淮河右岸最大的支流,也是淮河中游重要的水源地. 为了解淠河流域河源溪流鱼类的空间分布格局及其主要影响因素,本研究于2015年4-5月对6条河源溪流鱼类及其环境因子进行了调查. 研究结果表明,6条溪流共采集鱼类19种,其中杂食性种类占57.9%,肉食性和植食性种类分别占26.3%和15.8%. 所有种类中,宽鳍鱲(Zaccoplatypus)是主要优势种,绿太阳鱼(Lepomiscyanellus)为研究区域首次报道的外来入侵种. 就6条溪流各样点的平均值而言,淠河西部3条溪流鱼类种类数及个体数均明显高于东部3条溪流,但重量却并没有类似趋势. Sorensen相似性分析表明,6条溪流鱼类组成具有较高相似性,且相对较小值主要位于高、低海拔溪流之间. 除趋势对应分析二维空间排序与相似性分析结果一致. Pearson相关分析表明,影响鱼类种类数的主要是局域栖息地参数(包括海拔、流速、底质、水深、河宽和电导率)和溪流的空间位置参数(溪流级别、流量量级和下游量级);影响鱼类个体数和重量的均仅有局域栖息地参数中的底质因素. 本研究结果可为淠河流域鱼类的保护和管理提供重要的基础资料.
淠河流域;鱼类群落;空间分布;影响因素
河流是具有水、陆间物质与能量交换的开放生态系统[1],与湖泊、水库等静水生态系统相比,其水文环境具有极高的空间异质性和周期性[2]. 河源溪流(headwater stream)是大型河流的源头支流,常位于海拔较高的山区地带. 与大型河流干流相比,其栖息地结构较为简单、营养物质贫乏、水文动荡更为明显、物种多样性较低但特有性高[3-4]. 因此,河源溪流生态系统更为脆弱,对外界干扰的抵抗力和恢复力都较低,一旦受到人为破坏将更难恢复. 鱼类作为河源溪流的高级消费者,对溪流生态系统结构的稳定和功能的维持至关重要. 在长期进化过程中,溪流鱼类已经逐步形成了相应的形态特征、物候节律和生活史对策,使其能够耐受甚至受益于河源溪流这种独特的自然环境[5]. 然而,随着人类干扰的增加(如非法捕捞、采砂、低头坝建设等),我国多数溪流鱼类资源已受到严重威胁,导致其物种多样性和群落结构均发生较大变化. 而我国目前对鱼类研究主要集中于湖泊和大江大河的现状[6],必然不利于山区溪流鱼类资源的恢复和发展.
淮河是我国七大河流之一,自西向东流经河南、湖北、安徽和江苏四省,沿途支流众多,水系复杂. 较特殊的是,安徽省内淮河两岸支流除淠河和史河发源于大别山区,属于山溪性常年型河流外,其他支流均为平原型河流(极易断流,属间歇性河流)[7]. 目前,国内对淮河干流鱼类基本没有研究,而关于淮河支流鱼类的报道也较为少见. 淠河作为淮河流域少有的山溪性河流,同时也是淮河中游重要的水源地,历史上有丰富的鱼类资源. 近几十年来,由于流域内大规模的采砂和筑坝,鱼类资源已经严重衰退. 因此,本研究于2015年4-5 月对淠河流域6条河源溪流进行了鱼类调查,其主要目的是:1)了解淠河河源溪流鱼类的物种多样性和空间分布格局;2)确定影响河源溪流鱼类空间分布的主要影响因素. 其研究结果不仅对淠河流域鱼类资源的保护和管理具有重要意义,还可以为淮河的鱼类学研究积累基础资料.
1 材料与方法
1.1 研究区域概况
淠河位于安徽省西南部,是安徽省内淮河右岸最大的支流,全长260 km,年均径流量4.96×108m3,占安徽省境内淮河水量的20.1%. 淠河发源于大别山区,流经霍山县、岳西县、六安市,于正阳关汇入淮河. 淠河由于源于山区,植被覆盖度高且降水充沛,因此属于山溪性常年型河流[7].
淠河流域属于亚热带季风气候区,流域面积6000多平方公里,其中山区占70.4%,丘陵占23.2%,平原仅占6.4%[7]. 年均气温14~15℃、年均降水量1250~1400 mm,雨量充沛,但季节差异较大. 淠河流域河源溪流根据其发源地的不同,可分为东、西两个区域. 东部源头溪流发源于岳西县大别山北麓,主要有漫水河、板河、上东河和头陀河等;西部源头溪流发源于金寨县大别山北麓,主要有西淠河、青龙河和毛坦河;东、西两个区域源头溪流在六安市两河口汇合后始称淠河(图1). 由于淠河流域拥有丰富的砂石资源,从1990s开始大规模采砂现象随处可见,区域内生态环境已受到严重破坏,被国家治淮体系列为重点治理河流.
1.2 采样方法
2015年4-5月对淠河流域河源溪流鱼类进行了调查. 淠河西部所选河源溪流为西淠河、青龙河和毛坦河,淠河东部所选河源溪流为漫水河、上东河和板河. 根据不同溪流的河网复杂性及野外可到达性,分别设置4~9个样点,全流域共设置36个样点(图1). 每个样点鱼类采集时采取1人以背负式捕鱼器电捕,2人手持捞网跟随,以“之”字形路线在可涉水区域进行捕捞. 采集时间为30 min,采样长度为100 m,捕捞区域包括可涉水的深潭、浅滩、急流等不同生境. 采集后的标本在新鲜状态下进行鉴定,并统计不同物种的个体数和重量(精确到0.1 g). 对于疑难种类,用10%甲醛固定后带回实验室鉴定.
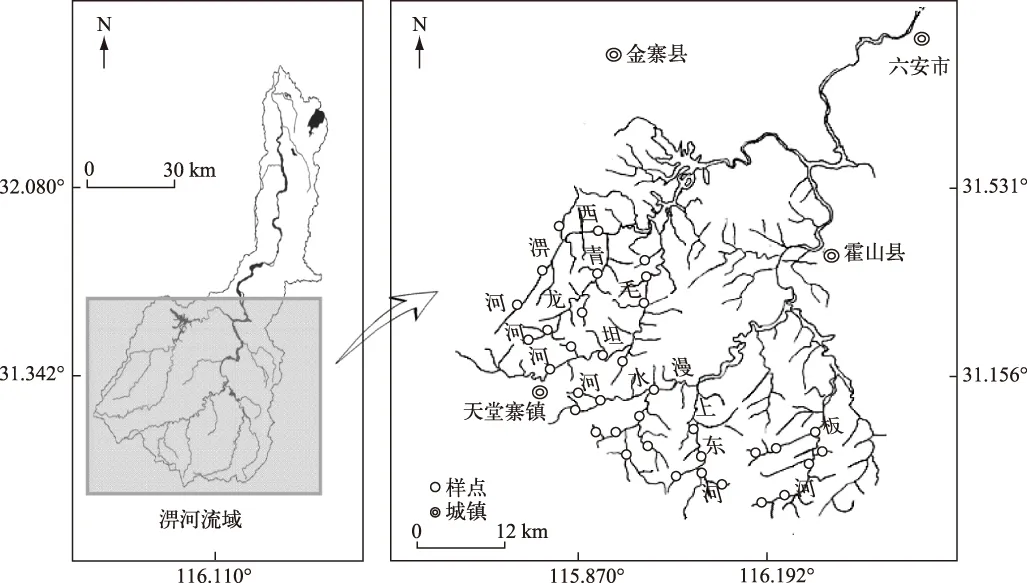
图1 淠河流域河源溪流鱼类采样点位置Fig.1 Location of the fish sampling sites in headwater streams of the Pihe River Basin
每个采样点测定的环境因子分为4大类,即局域栖息地参数、化学参数、生物参数和溪流空间位置参数. 其中,局域栖息地参数包括海拔、水宽、水深、流速、底质类型、水温、溶解氧和电导率等指标. 海拔采用etrex型手持式GPS测定,水宽采用Trupulse 200型激光测距仪测定,水深和流速采用FP211型流速仪测定,底质类型采用打分法判定,水温、溶解氧和电导率采用便携式水质分析仪测定. 化学参数包括pH、总氮(TN)、总磷(TP)和氨氮(NH3-N)等参数,每个样点取水样后采用标准方法进行室内分析测定[8]. 生物参数主要包括大型底栖动物的种类数、密度和生物量. 使用索伯网(30 cm×30 cm)在调查河段的左、中、右侧分别进行一次定量采集[9],带回实验室后鉴定. 溪流空间位置参数包括溪流级别、流量量级和下游量级3个参数. 溪流级别的划分参照Strahler的方法,即最小的溪流为一级溪流,两个同级别溪流汇合后引起级别的上升[10];流量量级代表某样点上游所有不分支源头溪流的数量[11],下游量级代表某样点下游连接溪流的流量量级[12].
1.3 数据分析
所有鱼类根据其食性不同分为肉食性、植食性和杂食性3类;根据其来源不同分为本地种、本地入侵种(native invasion)和外来入侵种3类[13]. 分别计算每种鱼类的出现频率(出现频率=某物种出现的样点个数/全部样点数)和相对丰度(相对丰度=某物种的个体数/全部物种的个体数). 如果某物种的出现频率≥40%,则视为常见种;如果处于10%~40%之间,则视为偶见种;如果<10%,则视为稀有种[14]. 根据相对重要性指数(Index of Relative Importance,IRI)来判断不同鱼类的优势度,如果其值>100则视为优势种[15],计算公式为:相对重要性指数=出现频率×相对丰度×104[16].
单因素方差分析(One-way ANOVA)用于检验6条溪流环境因子之间的差异显著性,如果有显著差异,采用Tukey多重比较来检验组间的差异. Sorensen相似性分析用于检验6条溪流之间的物种相似性.S=2c/(a+b),式中S为相似性系数,c为两个溪流中共有的物种数,a、b分别为两个溪流各自的物种数. 运用CANOCO 软件,采用除趋势对应分析(Detrended Correspondence Analysis, DCA)对6条河流的样点进行二维排序. 由于2个样点没采集到鱼类,所以实际只有34个样点用于排序. 运用SPSS 13.0 软件,采用Pearson相关分析检验鱼类种类数、个体数、重量和4大类环境因子的相关性,如果P<0.05,则认为有显著相关关系.
2 研究结果
2.1 环境参数
6条溪流基本环境参数见表1. 方差分析表明,6条溪流生物参数(底栖动物种类数、密度和生物量)无显著差异. 就局域栖息地和水化学参数而言,底质类型、水深、河宽、水温、溶解氧和pH这6个指标无显著差异,而海拔、流速、电导率、TN、TP和NH3-N存在显著差异. 与淠河西部3条河源溪流相比,淠河东部河源溪流中的上东河和板河海拔相对更高、流速更大,而电导率和营养物质浓度却更低.
表1 淠河流域6条河源溪流相关环境参数*
Tab.1 Environmental parameters in six headwater streams of the Pihe River Basin
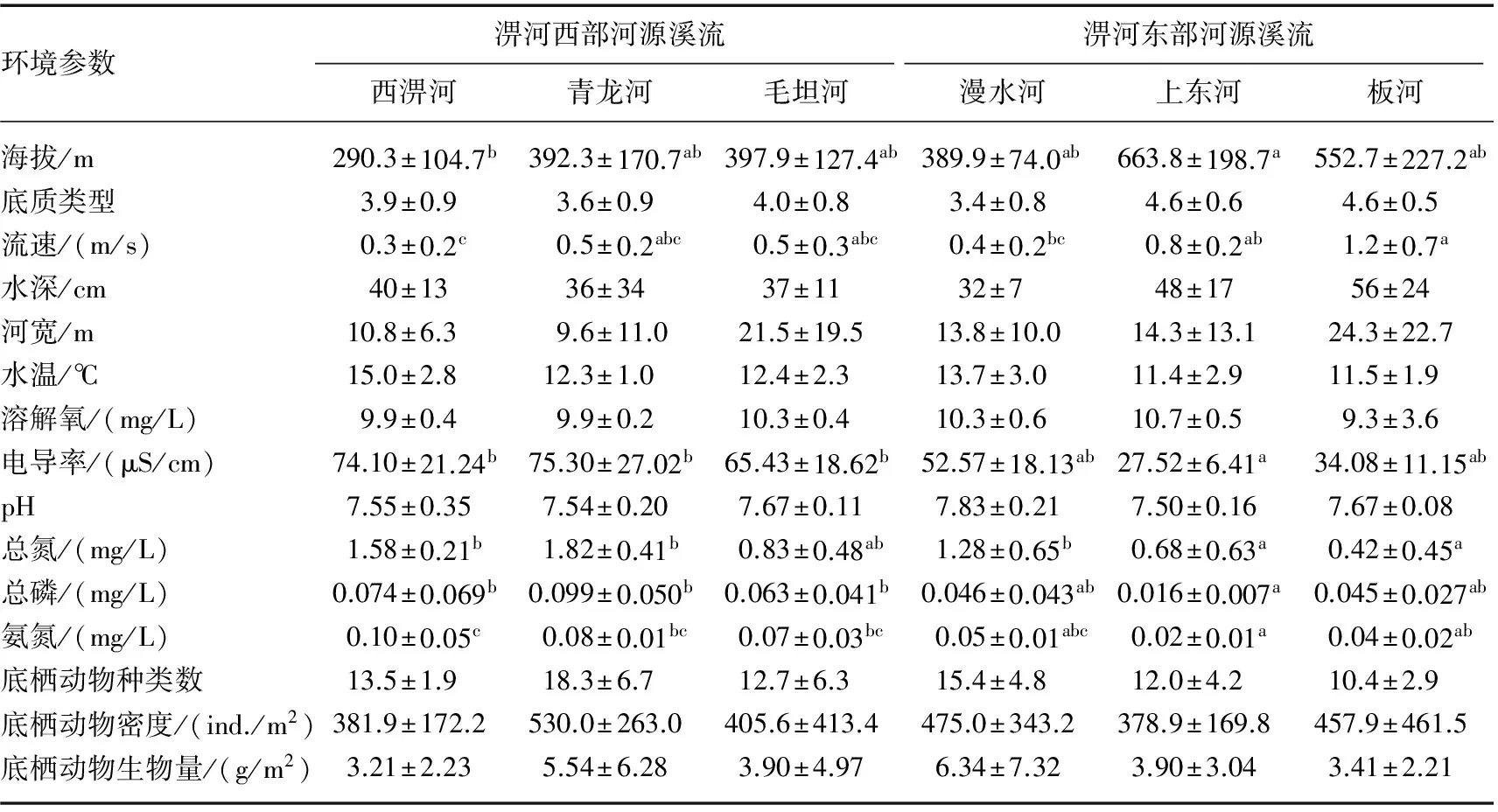
环境参数淠河西部河源溪流淠河东部河源溪流西淠河青龙河毛坦河漫水河上东河板河海拔/m290.3±104.7b392.3±170.7ab397.9±127.4ab389.9±74.0ab663.8±198.7a552.7±227.2ab底质类型3.9±0.93.6±0.94.0±0.83.4±0.84.6±0.64.6±0.5流速/(m/s)0.3±0.2c0.5±0.2abc0.5±0.3abc0.4±0.2bc0.8±0.2ab1.2±0.7a水深/cm40±1336±3437±1132±748±1756±24河宽/m10.8±6.39.6±11.021.5±19.513.8±10.014.3±13.124.3±22.7水温/℃15.0±2.812.3±1.012.4±2.313.7±3.011.4±2.911.5±1.9溶解氧/(mg/L)9.9±0.49.9±0.210.3±0.410.3±0.610.7±0.59.3±3.6电导率/(μS/cm)74.10±21.24b75.30±27.02b65.43±18.62b52.57±18.13ab27.52±6.41a34.08±11.15abpH7.55±0.357.54±0.207.67±0.117.83±0.217.50±0.167.67±0.08总氮/(mg/L)1.58±0.21b1.82±0.41b0.83±0.48ab1.28±0.65b0.68±0.63a0.42±0.45a总磷/(mg/L)0.074±0.069b0.099±0.050b0.063±0.041b0.046±0.043ab0.016±0.007a0.045±0.027ab氨氮/(mg/L)0.10±0.05c0.08±0.01bc0.07±0.03bc0.05±0.01abc0.02±0.01a0.04±0.02ab底栖动物种类数13.5±1.918.3±6.712.7±6.315.4±4.812.0±4.210.4±2.9底栖动物密度/(ind./m2)381.9±172.2530.0±263.0405.6±413.4475.0±343.2378.9±169.8457.9±461.5底栖动物生物量/(g/m2)3.21±2.235.54±6.283.90±4.976.34±7.323.90±3.043.41±2.21
*数值为平均值±标准差;每行上标不同字母表示各河流间存在显著差异(P< 0.05).
2.2 种类组成
6条溪流共采集鱼类1762尾,隶属于3目7科19种. 其中鲤科鱼类种类最多,为11种;其次是鳅科和虾虎鱼科,均为2种;其他各科均只有1种. 宽鳍鱲、泥鳅和原缨口鳅出现频率均大于40%,为研究区域的常见种;马口鱼、鲫、麦穗鱼、高体鰟鲏、银鮈、细纹颌须鮈、子陵吻虾虎、波氏吻虾虎和盎堂拟鲿的出现频率介于10%~40%之间,为偶见种;其余7种鱼类为稀有种(表2).
从食性来看,杂食性种类数最多,占总种类数的57.9%;肉食性和植食性鱼类分别占总种类数的26.3%和15.8%. 从物种来源看,本地入侵种共4种,分别是鲫、麦穗鱼、高体鳑鲏和泥鳅;外来入侵种1种,为绿太阳鱼;其他14种均为本地物种. 从重要值来看,宽鳍鱲是主要优势种,其他优势种还包括细纹颌须鮈、马口鱼、原缨口鳅、泥鳅、麦穗鱼和波氏吻虾虎(表2).
2.3 渔获物种类数、个体数和重量
淠河西部3条河源溪流渔获物种类数差异不大,西淠河、青龙河和毛坦河分别为13、10和12种;淠河东部3条河源溪流渔获物种类数差异较大,漫水河、上东河和板河分别为16、4和8种(表2). 就每个样点的平均值而言,淠河西部3条溪流渔获物种类数和个体数均明显高于东部3条溪流,但重量却并没有类似趋势(图2).
2.4 相似性分析和排序
Sorensen相似性分析结果表明,6条溪流之间具有较高的物种相似性,最大值为西淠河和青龙河之间的78.26%,最小值为青龙河和板河之间的44.44%(表3). 6条溪流34个样点的DCA二维排序图中,前3轴特征值分别为0.90、0.36和0.24,解释了42.6%的物种变异. 第1轴和第2轴解释变量最多,所以选用前两轴来做二维排序图. 由于彼此之间较高的物种相似性,6条溪流不同样点分布区间和界限十分模糊,相互之间有较多重叠(图3).
2.5 影响因素分析
Pearson相关分析表明,影响鱼类种类数的主要是局域栖息地参数和溪流的空间位置参数,局域栖息地中主要的影响因子是海拔,其次是流速、底质类型、水深、河宽和电导率;空间位置中主要的影响因子是流量量级和下游量级,其次是河流级别. 影响鱼类个体数和重量的均仅有局域栖息地中的底质因素. 水化学相关参数和生物参数对鱼类种类数、个体数和重量均无显著影响(表4).
表2 淠河流域6条河源溪流鱼类种类组成
Tab.2 Catch composition in six headwater streams of the Pihe River Basin
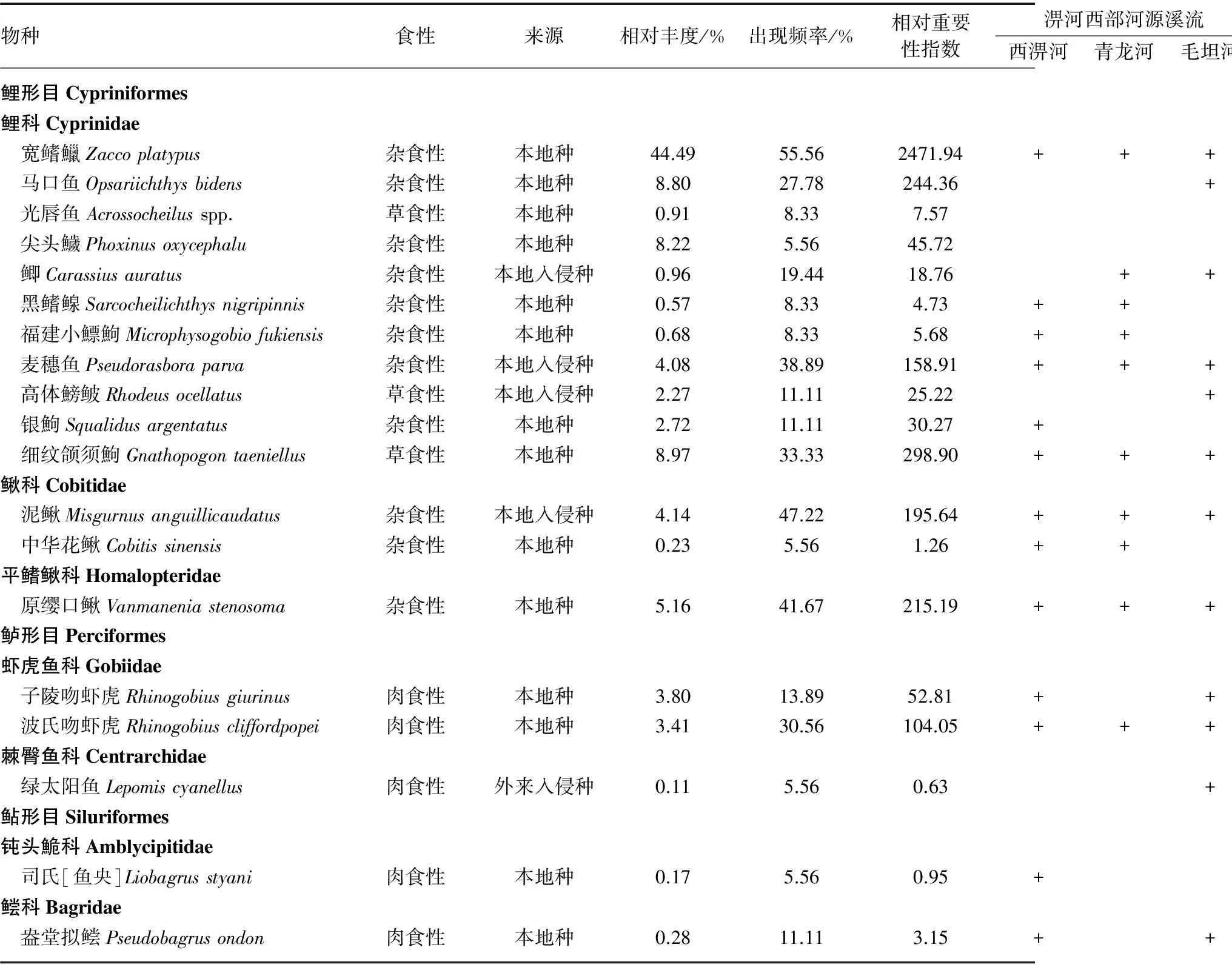
物种食性来源相对丰度/%出现频率/%相对重要性指数淠河西部河源溪流淠河东部河源溪流西淠河青龙河毛坦河漫水河上东河板河鲤形目Cypriniformes鲤科Cyprinidae 宽鳍鱲Zaccoplatypus杂食性本地种44.4955.562471.94++++++ 马口鱼Opsariichthysbidens杂食性本地种8.8027.78244.36+++ 光唇鱼Acrossocheilusspp.草食性本地种0.918.337.57++ 尖头鱥Phoxinusoxycephalu杂食性本地种8.225.5645.72+ 鲫Carassiusauratus杂食性本地入侵种0.9619.4418.76+++ 黑鳍鳈Sarcocheilichthysnigripinnis杂食性本地种0.578.334.73+++ 福建小鰾鮈Microphysogobiofukiensis杂食性本地种0.688.335.68+++ 麦穗鱼Pseudorasboraparva杂食性本地入侵种4.0838.89158.91++++ 高体鰟鲏Rhodeusocellatus草食性本地入侵种2.2711.1125.22++ 银鮈Squalidusargentatus杂食性本地种2.7211.1130.27+++ 细纹颌须鮈Gnathopogontaeniellus草食性本地种8.9733.33298.90++++++鳅科Cobitidae 泥鳅Misgurnusanguillicaudatus杂食性本地入侵种4.1447.22195.64++++++ 中华花鳅Cobitissinensis杂食性本地种0.235.561.26++平鳍鳅科Homalopteridae 原缨口鳅Vanmaneniastenosoma杂食性本地种5.1641.67215.19++++++鲈形目Perciformes虾虎鱼科Gobiidae 子陵吻虾虎Rhinogobiusgiurinus肉食性本地种3.8013.8952.81++ 波氏吻虾虎Rhinogobiuscliffordpopei肉食性本地种3.4130.56104.05++++棘臀鱼科Centrarchidae 绿太阳鱼Lepomiscyanellus肉食性外来入侵种0.115.560.63++鲇形目Siluriformes钝头鮠科Amblycipitidae 司氏[鱼央]Liobagrusstyani肉食性本地种0.175.560.95++鲿科Bagridae 盎堂拟鲿Pseudobagrusondon肉食性本地种0.2811.113.15+++
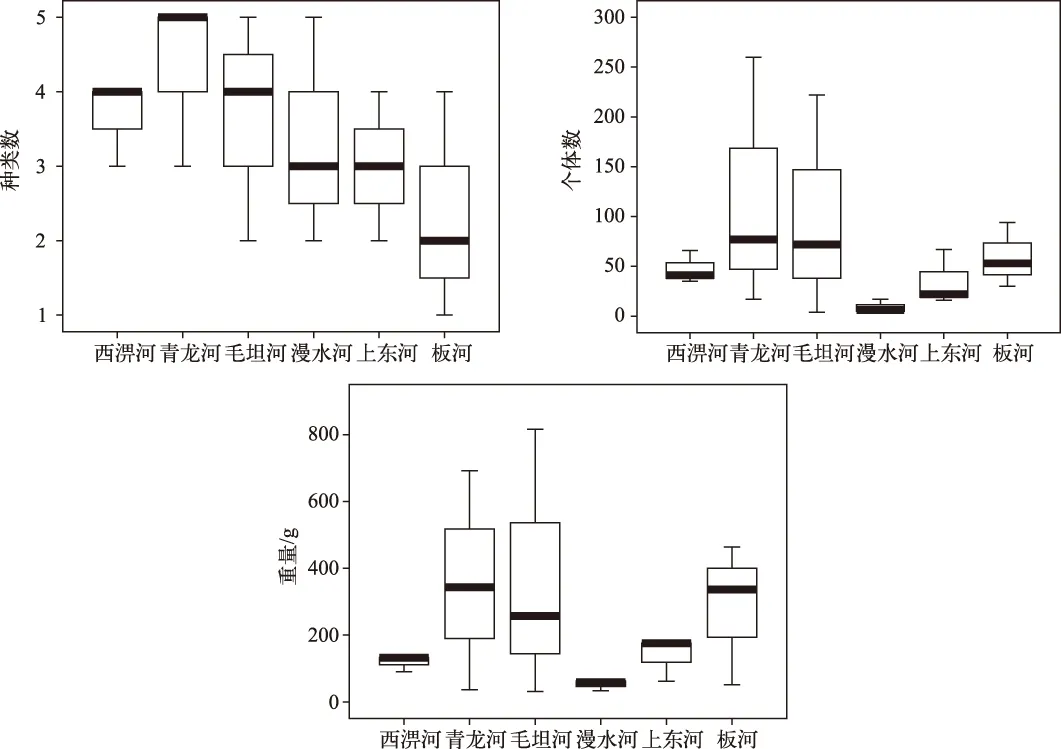
图2 淠河流域6条河源溪流各样点渔获物种类数、个体数和重量对比Fig.2 Comparison of the species number, individual number,and weight of catch in sites of six headwater streams of the Pihe River Basin
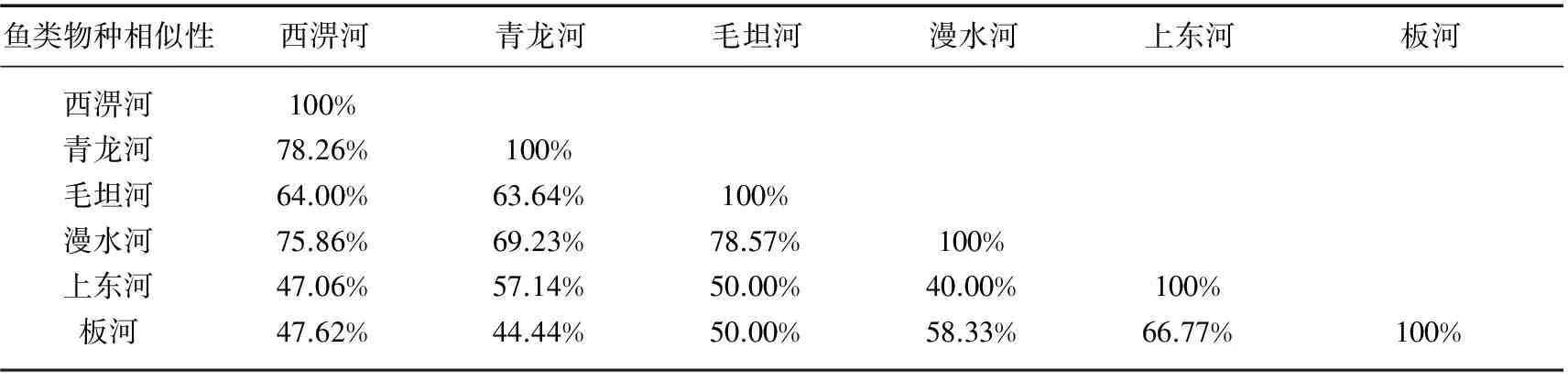
鱼类物种相似性西淠河青龙河毛坦河漫水河上东河板河西淠河100%青龙河78.26%100%毛坦河64.00%63.64%100%漫水河75.86%69.23%78.57%100%上东河47.06%57.14%50.00%40.00%100%板河47.62%44.44%50.00%58.33%66.77%100%
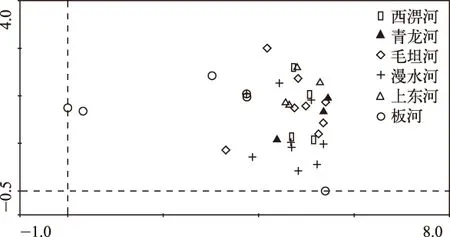
图3 淠河流域6条河源溪流34个样点的DCA二维排序Fig.3 Two dimensional ordination diagram of 34 sites in six headwater streams of the Pihe River Basin
表4 鱼类种类数、个体数、重量和各环境因子的Pearson相关关系
Tab.4 Pearson correlation between environmental parameters and species number, individual number and weight of the catch
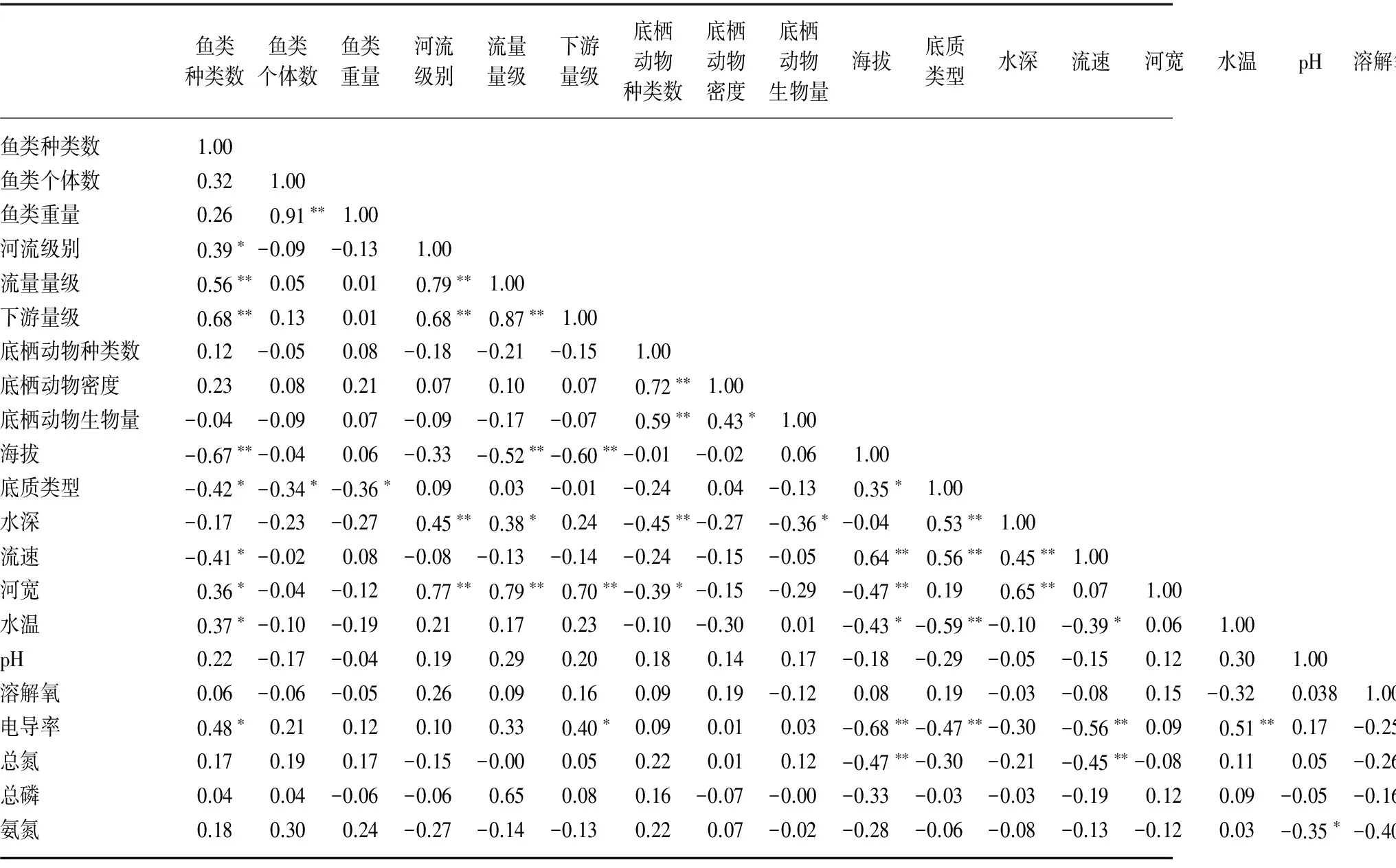
鱼类种类数鱼类个体数鱼类重量河流级别流量量级下游量级底栖动物种类数底栖动物密度底栖动物生物量海拔底质类型水深流速河宽水温pH溶解氧电导率总氮总磷氨氮鱼类种类数1.00鱼类个体数0.321.00鱼类重量0.260.91∗∗1.00河流级别0.39∗-0.09-0.131.00流量量级0.56∗∗0.050.010.79∗∗1.00下游量级0.68∗∗0.130.010.68∗∗0.87∗∗1.00底栖动物种类数0.12-0.050.08-0.18-0.21-0.151.00底栖动物密度0.230.080.210.070.100.070.72∗∗1.00底栖动物生物量-0.04-0.090.07-0.09-0.17-0.070.59∗∗0.43∗1.00海拔-0.67∗∗-0.040.06-0.33-0.52∗∗-0.60∗∗-0.01-0.020.061.00底质类型-0.42∗-0.34∗-0.36∗0.090.03-0.01-0.240.04-0.130.35∗1.00水深-0.17-0.23-0.270.45∗∗0.38∗0.24-0.45∗∗-0.27-0.36∗-0.040.53∗∗1.00流速-0.41∗-0.020.08-0.08-0.13-0.14-0.24-0.15-0.050.64∗∗0.56∗∗0.45∗∗1.00河宽0.36∗-0.04-0.120.77∗∗0.79∗∗0.70∗∗-0.39∗-0.15-0.29-0.47∗∗0.190.65∗∗0.071.00水温0.37∗-0.10-0.190.210.170.23-0.10-0.300.01-0.43∗-0.59∗∗-0.10-0.39∗0.061.00pH0.22-0.17-0.040.190.290.200.180.140.17-0.18-0.29-0.05-0.150.120.301.00溶解氧0.06-0.06-0.050.260.090.160.090.19-0.120.080.19-0.03-0.080.15-0.320.0381.00电导率0.48∗0.210.120.100.330.40∗0.090.010.03-0.68∗∗-0.47∗∗-0.30-0.56∗∗0.090.51∗∗0.17-0.251.00总氮0.170.190.17-0.15-0.000.050.220.010.12-0.47∗∗-0.30-0.21-0.45∗∗-0.080.110.05-0.260.39∗1.00总磷0.040.04-0.06-0.060.650.080.16-0.07-0.00-0.33-0.03-0.03-0.190.120.09-0.05-0.160.42∗0.41∗1.00氨氮0.180.300.24-0.27-0.14-0.130.220.07-0.02-0.28-0.06-0.08-0.13-0.120.03-0.35∗-0.40∗0.38∗0.35∗0.48∗∗1.00
*表示P<0.05; **表示P< 0.01; ***表示P<0.001.
3 讨论
3.1 种类组成和空间分布
一般情况下,人为改变溪流的物理栖息地条件会降低地方性敏感性鱼类的适合度,同时提高耐受性鱼类的适合度,使原来栖息于某流域中下游河段的鱼类成功入侵其上游,导致“本土入侵”现象发生[17-18]. 本研究在淠河河源溪流中共调查到19种鱼类,但本地入侵种和外来入侵种却分别为4种和1种,这表明淠河河源溪流的生态环境已受到较大改变. 为了灌溉和生活用水,调查中发现小型水坝(主要是低头坝,河水可自然漫过)在淠河流域随处可见. 虽然小型水坝对溪流的影响相对较小且更加局域化[19],但其在坝上区域会抬高水位、降低流速,产生相对的静水区. 本研究中4种本地入侵种(鲫、麦穗鱼、高体鳑鲏和泥鳅)均喜栖息于静水或缓流水环境,因此,小型水坝建设可能是其成功入侵并定殖的主要原因. 此外,本研究还首次在大别山区发现外来入侵种绿太阳鱼[20],该物种原产北美,1999年作为观赏和游钓用鱼引入我国,主要在广东和广西地区人工养殖[21]. 由于其种群数量在一定程度上随环境的恶化而增加,因此美国鱼类学家Karr早在1981年便把绿太阳鱼单独列出,把其个体数量百分比作为河流健康评价的一个重要指标[22]. 对研究区域2015年7月的后续调查表明,绿太阳鱼目前已经形成了稳定的种群. 由于河源溪流鱼类个体较小、生长缓慢,且缺乏凶猛性鱼类,因此肉食性的绿太阳鱼对本地种威胁极大,亟需采取有效防控措施.
在自然情况下沿河源至下游的纵向梯度,河流生态系统中众多生物因子和非生物因子都表现出规律性和连续性的变化[4],比如流速逐渐减小,底质逐渐由基岩、砾石变为细砂、淤泥,物种多样性逐渐增加等. 本研究对6条溪流环境因子的对比表明(表1),随着平均海拔的显著下降,流速显著降低,而电导率却显著增加,与上述规律一致. 此外,低海拔区域一般更适合人类居住,因此低海拔溪流受人为干扰程度也要远大于高海拔溪流. 本研究中6条溪流营养物质浓度随着平均海拔的降低表现出显著增加的趋势也与此一致. 结果表明,东部3条源头溪流每个样点的平均种类数和个体数都明显低于西部3条源头溪流,除溪流自身的空间位置外,东部源头溪流相对较高的海拔可能是造成这一结果的主要原因. 就鱼类的物种相似性而言,6条溪流表现出两个特点:1)总体相似性很高,DCA空间排序也证明了这一点;2)高海拔溪流(板河和上东河)与相对低海拔溪流之间的相似性低于高海拔或低海拔溪流之间. 由于6条溪流同处淠河流域大别山区,从大流域尺度上其降雨、温度等自然条件差异不大,因此总体物种相似性较高. 但是,从局域栖息地(小流域尺度)环境来看,又各具其自身的特殊性. 比如尖头鱥常栖息于高海拔、冷水性溪流当中,其种群分布受高海拔与低水温等的限制[23],仅在板河的部分高海拔样点出现.
3.2 主要影响因素
一般而言,局域鱼类群落是生物因素、非生物因素和历史因素(如物种形成与灭绝)联合作用的产物[24]. 这些因素对局域鱼类群落有重要的过滤作用,在不同的空间尺度上影响鱼类的种类数、个体数和重量等. 就山区源头溪流而言,国内部分研究结果已经表明,影响鱼类群落结构的主要因素是局域栖息地环境条件和溪流的空间位置;局域栖息地因素中又主要包括海拔、流速、底质、电导率、水温和水深等参数,而溪流的空间位置则主要包括溪流级别、河流量级和下游量级等参数[23, 25-27]. 本研究中,影响6条溪流鱼类种类数的主要是局域栖息地条件和溪流的空间位置参数,与上述结论一致. 在局域栖息地参数中,主要是海拔因素,其次是流速、底质、水深、河宽和电导率,这是因为海拔与其他相关参数之间都存在显著的相关关系(P<0.05),国内对皖河流域鱼类的研究也有类似的结果[25]. 就溪流的空间位置而言,本研究中溪流级别、流量量级和下游量级3个参数均与鱼类种类数呈显著正相关. 溪流级别概念于1962年首次应用到溪流生态学领域,一直是生态学家分析鱼类空间分布重要的环境因子[28]. 一般情况下,随着溪流级别的增大鱼类种类数会逐渐增加,这是因为栖息地的多样性和复杂性也逐渐增加[29]. 流量量级为采样点上游全部一级河流的总和,所以该指标直接反映溪流流量的大小,进而间接反映溪流级别的大小. 溪流下游量级对鱼类多样性影响也有大量研究,野外调查常发现直接汇入大河的一级支流鱼类多样性要高于源头性一级支流的多样性[30]. 对这一现象,生态学家普遍认为是由于干流为前者鱼类的迁入提供了源泉[31]. 对鱼类数量和重量的相关分析结果表明,其影响因素仅有底质. 底质常被认为是反映栖息地状况重要的指标之一,因为底质可以为很多溪流鱼类提供摄食、避敌与繁殖的场所[32]. 以山区溪流常见的优势种为例,马口鱼和宽鳍鱲对底质的选择较宽,属于溪流生态系统中的泛化种;而光唇鱼、尖头鱥和原缨口鳅多栖息于大石块的夹缝中;中华花鳅和泥鳅则分别栖息于砂质和淤泥底质中.
在山区溪流环境因子调查中,受野外实验条件限制,影响鱼类生存的化学因素和生物因素一般较少测定. 淠河流域6条溪流营养水平随平均海拔降低而显著上升,但对鱼类的种类数、个体数和重量均没有显著影响,这可能主要是山区溪流总体营养水平较低造成的. 山区溪流主要的营养来源是植物碎屑和生活污水排放,但山区较高的植被覆盖度可以起到很好的拦截和过滤作用,从而缓解了山区溪流自净能力和抵抗力较弱的缺点. 此外,6条河源溪流调查到的19种鱼类中,肉食性和杂食性鱼类占了16种,因此底栖动物是山区溪流鱼类重要的食物来源. 但ANOVA和相关分析结果表明,6条河流之间底栖动物的种类数、密度和生物量均无显著差异,且与鱼类的种类数、个体数和重量均无显著相关关系,这可能主要是由于底栖动物的野外采集中没有考虑到局域栖息地斑块类型. 根据水文形态特征可将溪流河道划分为深潭、急滩和湍流等不同的栖息地斑块类型[33],并分别栖息有不同生活习性的鱼类. 而本研究中底栖动物的采集是在调查河段的左、中、右侧分别进行,未涵盖全部栖息地斑块类型. 关于底栖动物与山区溪流鱼类之间的关系,还有待进一步研究.
[1] Ward JV. The four-dimensional nature of lotic ecosystems.JournaloftheNorthAmericanBenthologicalSociety, 1989, 8: 2-8.
[2] Allan JD. Stream ecology: Structure and function of running waters. London: Chapman and Hall, 1995.
[3] Grossman GD, Dowd JF, Crawford M. Assemblage stability in stream fishes: A review.EnvironmentalManagement, 1990, 14(5): 661-671.
[4] Vannote RL, Minshall GW, Cumins KWetal. The river continuum concept.CanadianJournalofFisheriesandAquaticSciences, 1980, 37(1): 130-137.
[5] Lytle DA, Poff NL. Adaptation to natural flow regimes.TrendsinEcologyandEvolution, 2004, 19: 94-100.
[6] Liu Jiankang, Tang Tao. Freshwater ecology. In: Li Wenhua, Zhao Jingzhu eds. Review and prospect on the study of ecology. Beijing: Meteorological Press, 2004. [刘建康, 唐涛. 淡水生态学. 见: 李文华, 赵景柱编. 生态学研究回顾与展望. 北京: 气象出版社, 2004.]
[7] Compilation commission of local record of Anhui Province ed. Record of Anhui Province-natural environment. Beijing: Fangzhi Press, 1999. [安徽省地方志编纂委员会. 安徽省志-自然环境志. 北京: 方志出版社, 1999.]
[8] "Monitoring and analysis method of water and wastewater" editorial board of State Environmental Protection Administration of China ed. Monitoring and analysis method of water and wastewater: Fourth Edition. Beijing: China Environmental Science Press, 2002. [国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法: 第4版. 北京: 中国环境科学出版社, 2002.]
[9] Zhang Xiaoke, Xuan Hao, Wang Huilietal. Macrozoobenthic community structure in different order streams of the Qingyi River basin.ResourcesandEnvironmentintheYangtzeBasin, 2014, 23(12): 1659-1664. [张晓可, 宣昊, 王慧丽等. 青弋江流域不同级别河流底栖动物群落结构研究. 长江流域资源与环境, 2014, 23(12): 1659-1664.]
[10] Strahler AN. Quantitative analysis of watershed geomorphology.TransactionsofAmericanGeophysicalUnion, 1957, 38(6): 913-920.
[11] Shreve RL. Statistical law of stream numbers.TheJournalofGeology, 1996, 74(1): 17-37.
[12] Osborne LL, Wiley MJ. Influence of tributary spatial position on the structure of warmwater fish communities.CanadianJournalofFisheriesandAquaticScience, 1992, 49(4): 671-681.
[13] Chu L, Wang WJ, Zhu Retal. Variation in fish assemblages across impoundments of low-head dams in headwater streams of the Qingyi River, China: Effects of abiotic factors and native invaders.EnvironmentalBiologyofFishes, 2015, 98(1): 101-112.
[14] Dai Yinggui, Li Min. Fish resources around Fanjing Mountain, Guizhou.BiodiversityScience, 2006, 14(1): 55-64. [代应贵, 李敏. 梵净山及邻近地区鱼类资源的现状. 生物多样性, 2006, 14(1): 55-64.]
[15] Yan Yunzhi, Zhan Yaojun, Chu Lingetal. Effects of stream size and spatial position on stream-dwelling fish assemblages,ActaHydrobiologicaSinica, 2010, 34(5): 1022-1030. [严云志, 占姚军, 储玲等. 溪流大小及其空间位置对鱼类群落结构的影响. 水生生物学报, 2010, 34(5): 1022-1030.]
[16] Krebs CJ. Ecological methodology. New York: Harper Collins, 1989.
[17] Rahel FJ. Homogenization of freshwater faunas.AnnualReviewofEcologyandSystematics, 2002, 33: 291-315.
[18] Scott MC,Helfman GS. Native invasions, homogenization, and the mismeasure of integrity of fish assemblages.Fisheries, 2001, 26(11): 6-15.
[19] Gillette DP, Tiemann JS, Edds DRetal. Spatiotemporal patterns of fish assemblage structure in a river impoundment by low-head dams.JournalofGreatLakesResearch, 2009, 2005(3): 539-549.
[20] Wan An, Zhang Xiaoke, Fang Yunxiangetal. Being vigilant to spreading and breeding of the green sunfish in Tapieh mountain area.ActaHydrobiologicaSinica, 2015, 39(4): 685. [万安, 张晓可, 方云祥等.警惕外来种绿太阳鱼在大别山区的扩散与繁殖. 水生生物学报, 2015, 39(4): 685.]
[21] Zheng Huifang, Lin Gang, Qin Zhibiao. Biology habit ofLepomiscyanellus.ChineseJournalofZoology, 2006, 41(2): 15-23. [郑惠芳, 林岗, 覃志彪. 绿色太阳鱼的生物学习性. 动物学杂志, 2006, 41(2): 15-23.]
[22] Karr JR. Assessment of biotic integrity using fish communities.Fisheries, 1981, 6(6): 21-27.
[23] Yan YZ, Xiang XY, Chu Letal. Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed, the Qingyi Stream, China.EcologyofFreshwaterFish, 2011, 20(2): 199-208.
[24] Dauwalter DC, Splinter DK, Fisher WLetal. Biogeography, ecoregions, and geomorphology affect fish species composition in streams of eastern Oklahoma, USA.EnvironmentalBiologyofFishes, 2008, 82(3): 237-249.
[25] Li Yanhui, Yan Yunzhi, Zhu Renetal. Spatial variations in fish assemblages within the headwater streams of the Wanhe watershed: A review network-based approach,JournalofFisherySciencesofChina, 2015, 21(5): 988-999. [李艳慧, 严云志, 朱仁等. 基于河流网络体尺度的皖河河源溪流鱼类群落的空间格局. 中国水产科学, 2015, 21(5): 988-999.]
[26] Wang Wenjian, Chu Ling, Si Chunetal. Spatial and temporal patterns of stream fish assemblages in the Qiupu Headwaters National Wetland Park.ZoologicalResearch, 2013, 34(4): 417-428. [王文剑, 储玲, 司春等. 秋浦河源国家湿地公园鱼类群落的时空格局. 动物学研究, 2013, 34(4): 417-428.]
[27] Chu Ling, Wang Wenjian, Yan Lilietal. Fish assemblages and longitudinal patterns in the headwater streams of the Chencun Reservoir in the Huangshan Area.ActaEcologicaSinica, 2015, 35(3): 900-910. [储玲, 王文剑, 闫莉莉等. 黄山陈村水库上游河源溪流的鱼类群落及其纵向梯度格局. 生态学报, 2015, 35(3): 900-910.]
[28] Kuehne RA. A classification of streams, illustrated by fish distribution in an eastern Kentucky creek.Ecology, 1962, 74(4): 1659-1673.
[29] Matthews WJ. Fish faunal “breaks” and stream order in the eastern and central United States.EnvironmentalBiologyofFishes, 1986, 17(2): 81-92.
[30] Groman OT. Assemblage organization of stream fishes: The effects of rivers on adventitious streams.AmericanNaturalist, 1986, 128(4): 611-616.
[31] Taylor CM, Warren ML. Dynamics in species composition of stream fish assemblages: Environmental variability and nested subsets.Ecology, 2001, 82(8): 2320-2330.
[32] Bain MB, Stevenson NJ. Aquatic habitat assessment: Common methods. Bethesda: American Fisheries Society, 1999.
[33] Eros T, Grossman GD. Effects of within-patch habitat structure and variation on fish assemblage characteristic in Bernrcei stream, Hungary.EcologyofFreshwaterFish, 2005, 14: 256-266.
Fishes in headwater streams of the Pihe River Basin: Spatial distribution pattern and its main influencing factors
ZHANG Xiaoke1, WANG Huili1, WAN An1**, FANG Yunxiang2, LIU Zhigang1, ZHENG Aifang1, CHEN Minmin1& YU Daoping1**
(1:ResearchCenterofAquaticOrganismConservationandWaterEcosystemRestorationinAnhuiProvince,AnqingNormalUniversity,Anqing246011,P.R.China)(2:AnhuiAcademyofEnvironmentalSciencesResearch,Hefei230061,P.R.China)
Pihe River is not only the largest tributary on north bank of the Huaihe River in Anhui Province, but also the important water source area in middle reaches of the Huaihe River. To determine the spatial distribution pattern of fish community and its influenced factors, a field investigation was conducted in six headwater streams of the Pihe River Basin from April to May, 2015. The results showed that (1) a total of 1762 individuals were collected and identified as 19 species representing 7 families in the six headwater streams. Among them, the omnivore species is accounted for 57.9% of the total species, while the carnivore and herbivore only comprised 26.3% and 15.8%, respectively.Zaccoplatypuswas the main dominant species with an index of relative importance of 2471.94.Lepomiscyanelluswas the first reported exotic invasion species in this basin. (2) As for the average value per site of the six streams, the species number and individual number of the three west headwater streams were obviously higher than those of the three east headwater streams, but such a tendency was not found as for the weight of fish. (3) Sorensen similarity analyses showed that great similarity of the fish species was detected among the six streams, and there was the relative smaller similarity between high and low altitudes of streams. The detrended correspondence analysis ordination of the six streams was further examined for the accuracy of similarity analyses. (4) Pearson correlation indicated that the fish species number had significantly correlated with local habitat parameters (including altitude, velocity, substrate type, water depth, water wide, and conductivity) and with spatial location parameters (including stream order, link, and D-link). However, the individual number and fish weight were only significantly correlated with the river substrate type of the local habitat parameters. The results of this study can provide important basic data for the protection and management of fish resources in Pihe River Basin.
Pihe River Basin; fish community; spatial distribution; influencing factor
*安徽省教育厅自然科学研究重点项目(KJ2015A222)、安庆师范学院人才引进基金项目(K05000130032)和农业部热带亚热带水产资源利用与养殖重点实验室开放基金项目(041-160002001)联合资助. 2016-01-22收稿;2016-04-30收修改稿. 张晓可(1986~),男,博士,副教授;E-mail: zxksgsg@163.com.
*通信作者; E-mail: wananahnu@126.com, ahyudp@163.com.
J.LakeSci.(湖泊科学), 2017, 29(1): 176-185
DOI 10.18307/2017.0119
©2017 byJournalofLakeSciences
