IL-6诱导小鼠肌原细胞系C2C12 UⅡ/UT系统的高表达
2017-01-10鄢雯刘立新夏山王红霞
鄢雯 刘立新 夏山 王红霞
(1.首都医科大学燕京医学院,北京 101300;2.中国人民解放军66222部队医院,北京 102202; 3.首都医科大学基础医学院,北京 100084)
IL-6诱导小鼠肌原细胞系C2C12 UⅡ/UT系统的高表达
鄢雯1刘立新1夏山2王红霞3△
(1.首都医科大学燕京医学院,北京 101300;2.中国人民解放军66222部队医院,北京 102202; 3.首都医科大学基础医学院,北京 100084)
目的 观察外源给予IL-6对C2C12细胞中UⅡ/UT系统表达的影响。方法 培养小鼠肌原细胞系C2C12细胞株,应用胎盘蓝染色和测定细胞培养液中LDH的含量检测IL-6对细胞损伤的影响,应用放射免疫的方法检测细胞培养液和裂解液中UⅡ的含量,应用RT-PCR法测定C2C12中UT mRNA的表达。结果 外源给予不同浓度的IL-6,C2C12细胞裂解液中UⅡ的含量各组间无明显差异,但增加了细胞孵育液中UⅡ的含量,10 ng/mL、50 ng/mL和100 ng/mL时较对照组分别增加了104.9%(P<0.01)、86.4%(P<0.01)和94.3%(P<0.01)。低浓度IL-6(0.1 ng/mL、1 ng/mL、10 ng/mL和50 ng/mL)刺激明显增加了UT mRNA的表达,而高浓度的IL-6(100 ng/mL)刺激反而使UT mRNA的表达降低。 结论 IL-6作为T2DM发病因素之一的炎症因子,可能是T2DM时骨骼肌组织中UⅡ/UT系统表达增加的因素之一。
尾加压素Ⅱ; G蛋白偶联受体; 白介素-6
尾加压素Ⅱ(UⅡ)是从硬骨鱼的尾部下垂体中提纯的神经肽[1]。UⅡ与其受体(UT)结合后,可参与多种生物学效应[2-3]。2型糖尿病(T2DM)的重要特征之一是胰岛素抵抗(IR),肌肉组织是胰岛素抵抗的主要靶点,研究发现UⅡ在骨骼肌含量最高[4]。目前UⅡ和T2DM的关系亦引起了人们的重视。炎症因子的过度激活可能是胰岛素抵抗发展的一个基本步骤[5-6],骨骼肌UⅡ/UT系统改变是否与炎症因子有关?尚未见报道。本研究采用小鼠的肌原细胞系C2C12,外源给予不同浓度的白介素-6(IL-6),观察UⅡ/UT系统表达的变化,探讨影响T2DM时骨骼肌中UⅡ/UT系统高表达的因素。
1 材料和方法
1.1 细胞和试剂 小鼠肌原细胞系C2C12细胞株购自中国医学科学院基础所。胎牛血清和马血清购自Gibco公司;总RNA提取及反转录试剂盒购自Promega公司;UT-S(5’-GCATCTTCACCCTGACCATAA-3’)、UT-A引物(5’-CCCAGAAGAGAAGGACGATACC-3’)[7]、β-actin-S引物(5’-ATCTGGCACCACACCTTC-3’)及β-actin-A引物(5’-AGCCAGGTCCAGACGCA-3’)[8]均由上海生工生物工程技术服务有限公司合成。UⅡ放射免疫测定法试剂盒购自美国Phoenix PharmacINC公司,按照说明书操作。
1.2 实验方法 待细胞长到80%融合时,用2%马血清分化2~3 d,可见到细胞内有明显的肌管出现可用于实验。采用胎盘蓝染色和测定LDH含量观察IL-6对细胞的损伤;采用RT-PCR方法测定C2C12细胞中UT mRNA表达,重复3次独立试验;采用放射免疫测定法测定细胞培养液及细胞裂解液中U Ⅱ 的含量。
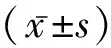
2 结 果
2.1 台盼蓝染色结果 台盼蓝染色是对细胞存活率比较精确的定量分析,在实验中给予不同浓度的IL-6后,测定台盼蓝染色阳性的细胞,以评价IL-6刺激对细胞死亡的影响。结果发现外源给予不同浓度的IL-6刺激24 h,各组未见染色阳性的细胞或仅见个别染色阳性的细胞,各组间无明显差异。
2.2 细胞培养液中LDH的含量 乳酸脱氢酶是代表组织或细胞受到损伤的一个指标,在实验中给予不同浓度的IL-6后,测定细胞培养液中LDH的含量,以评价IL-6刺激对细胞损伤的影响。结果发现给予不同浓度的IL-6刺激24 h,细胞培养液中LDH含量增高,0.1 ng/mL IL-6刺激时,培养液中LDH的含量为50 U/L,较对照组增高2.75倍,1 ng/mL和10 ng/mL IL-6刺激时,LDH的含量略有下降,但仍较对照组增高2倍和1.75倍,当IL-6刺激浓度升至50 ng/mL和100 ng/mL时,培养液中LDH浓度出现明显升高,分别为100 U/L 和159.1 U/L,较对照组分别增高5倍和8倍,差异有统计学意义(P<0.01)。
2.3 IL-6增加了C2C12细胞孵育液中UⅡ的含量 与对照组相比较,外源给予不同浓度的IL-6(0.1、1、10、50、100 ng/mL)对C2C12细胞裂解液中UⅡ的含量无明显影响,但增加了C2C12细胞孵育液中UⅡ的含量,0.1 ng/mL和1 ng/mL较对照组分别增加了13.1%和9.5%,差异无统计学意义(均P>0.05),而10 ng/mL、50 ng/mL和100 ng/mL时较对照组分别增加了104.9%、86.4%和94.3%,差异有统计学意义(均P<0.01)。见表1。

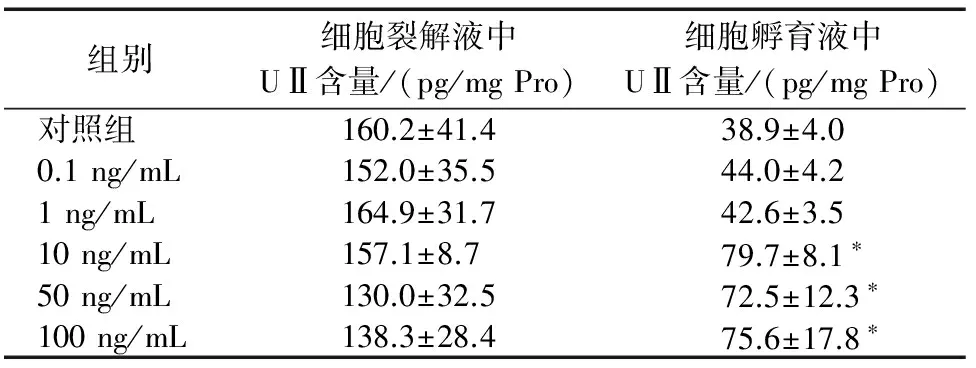
组别细胞裂解液中UⅡ含量/(pg/mgPro)细胞孵育液中UⅡ含量/(pg/mgPro)对照组160.2±41.438.9±4.00.1ng/mL152.0±35.544.0±4.21ng/mL164.9±31.742.6±3.510ng/mL157.1±8.779.7±8.1∗50ng/mL130.0±32.572.5±12.3∗100ng/mL138.3±28.475.6±17.8∗
注:与对照组相比,*P<0.01。
2.4 不同浓度的IL-6刺激对肌原细胞系C2C12中UT mRNA表达的影响 RT-PCR分析表明UT mRNA在C2C12细胞中表达,较低浓度的IL-6增加了UT mRNA的表达,不加模板的引物对照组的UT/β-actin光密度比值为(0.33±0.06);0.1 ng/mL IL-6刺激组为(0.4±0.05),较对照组增加了25.9%,差异无统计学意义;当IL-6的刺激浓度升高至1 ng/mL、10 ng/mL及50 ng/mL时,细胞中UT mRNA的表达水平出现明显升高,UT/β-actin光密度比值分别为(0.56±0.05)、(0.62±0.06)和(0.78±0.06),较对照组分别增加了71.8%、90.1%和138.9%(均P<0.05);然而,100 ng/mL IL-6刺激时UT mRNA的表达水平较对照组反而降低了34.4%(P<0.05)。UT基因RT-PCR扩增片段的电泳结果见图2。

注:1.对照组(不加模板的引物);2~6.分别为刺激浓度是0.1 ng/mL、1 ng/mL、10 ng/mL、50 ng/mL、100 ng/mL IL-6的处理组。图2 外源给予不同浓度的IL-6对C2C12细胞系中UT含量的影响
3 讨 论
IR是指胰岛素作用的靶器官对胰岛素作用的敏感性下降。胰岛素抵抗是一个慢性亚临床炎症过程,在这过程中胰岛素抵抗和炎症反应相互促动,引起一系列机体内环境的改变[9]。涉及胰岛素抵抗的主要靶器官包括肝脏、脂肪、肌肉等。其中肌肉组织在IR中的作用尤为重要。骨骼肌组成体质量的40%~50%,负责静息氧耗的20%~30%,负责生理状态下胰岛素调节的葡萄糖的处理的75%,因此也成为糖尿病及其胰岛素抵抗的主要靶点[4,10]。2009年,我室首先报道[11]了遗传性2型糖尿病小鼠(KKAy/Upj)骨骼肌U Ⅱ/UT系统上调;并在体外孵育的骨骼肌组织薄片上,观察到U Ⅱ 呈浓度(10-9~10-7mol/L)依赖地抑制胰岛素刺激的骨骼肌2-DG摄入。但什么因素影响了伴有胰岛素抵抗T2DM时骨骼肌中U Ⅱ/UT系统的高表达并不清楚。
IL-6是调节免疫反应的一个重要细胞因子,它具有促炎和抗炎作用。多种细胞可产生IL-6,包括骨骼肌。锻炼能够使肌肉释放大量的IL-6,表明IL-6在锻炼期间和锻炼后维持葡萄糖稳定中具有重要的作用。但有证据[12]表明IL-6血清水平随着年龄的增加而增加;IL-6的升高是T2DM发展的一个危险因素;在各种各样的胰岛素抵抗状态时循环中IL-6水平的升高,包括糖尿病和肥胖;大鼠在体情况下,急性IL-6处理可减少胰岛素刺激的骨骼肌葡萄糖摄取;由于发现IL-6常常在代谢性疾病中升高,普遍认为IL-6是慢性疾病的一个病因,是促使胰岛素抵抗和高脂血症发生的促炎因子[13]。U Ⅱ 在急慢性炎症中具有作用,而且M.Birker[14]等培养人的横纹肌肉瘤细胞系,发现IFN-γ增加UT受体表达上调。那么IL-6是否作为一个炎症因子刺激了骨骼肌组织中U Ⅱ/UT系统的上调,从而进一步引起骨骼肌胰岛素抵抗的发生目前尚无文献报道。
本研究在肌原细胞上观察外源给予IL-6对U Ⅱ/UT系统表达的影响,发现外源给予不同浓度的IL-6,用放射免疫方法测定到C2C12细胞裂解液中UⅡ的含量各组间无明显差异,但增加了细胞孵育液中UⅡ的含量,0.1 ng/mL和1 ng/mL较对照组分别增加了13.1%和9.5%,差异无统计学意义(均P>0.05),而10 ng/mL、50 ng/mL和100 ng/mL时较对照组分别增加了104.9%、86.4%和94.3%,差异有显著性(均P<0.01)。用RT-PCR的方法测定了C2C12细胞中UT mRNA的表达,结果发现低浓度IL-6(1 ng/mL、10 ng/mL和50 ng/mL)刺激明显增加了UT mRNA的表达,而高浓度的IL-6(100 ng/mL)刺激反而使UT mRNA的表达降低。以上结果表明IL-6作为T2DM发病因素之一的炎症因子,可能是T2DM时骨骼肌组织中UⅡ/UT系统表达增加的一个因素;至于高浓度时IL-6抑制UT mRNA表达的原因,可能是自身的代偿去拮抗UⅡ的作用,但由于高浓度IL-6增加了细胞孵育液中LDH的含量,因此不能排除是由于细胞损伤本身导致其UT mRNA表达的下降。
本研究首次报道了炎症因子IL-6可能是2型糖尿病小鼠骨骼肌UⅡ/UT系统上调的一个因素。鉴于T2DM时引起胰岛素抵抗的因素众多,是否还有其他因素参与了其表达的升高还值得进一步探讨。
[1] 卢宁,于海云,王睿,等.中枢活性氧介导尾加压素Ⅱ在自发性高血压大鼠的心血管效应[J].生理学报,2012,64(2):142-148.
[2] Vaudry H,Leprince J,Chatenet D,et al.International Union of Basic and Clinical Pharmacology.XCⅡ.Urotensin Ⅱ,urotensin Ⅱ-related peptide,and their receptor:from structure to function[J].Pharmacol Rev,2015,67(1):214-58.
[3] 杨冬花,付蓉,罗建华,等.心力衰竭大鼠不同时期尾加压素Ⅱ、内皮素-1和肾上腺髓质素水平变化及益气温阳活血方对其影响的研究[J].贵州医药,2012,36(1):4-7.
[4] Suzan T,Hakan K,Mehmet A E,Elif O,et al.Urotensin-Ⅱ level and its association with oxidative stress in early diabetic nephropathy[J].J Diabetes Complications,2015,29(1):115-119.
[5] Julia Beilfuss,Vivian Berg,Monica Sneve,Rolf Jorde,Elena Kamycheva.Effects of a 1-year supplementation with cholecalciferol on interleukin-6,tumor necrosis factor-alpha and insulin resistance in overweight and obese subjects[J].Cytokine,2012,60(3):870-874.
[6] Pedersen BK,Steensberg A,Schjerling P.Muscle-derived interleukin-6:possible biological effects[J].J Physiol,2001,15:329-337.
[7] Wang ZJ,Shi LB,Ding WH et al.Alteration of vascular urotensin Ⅱ receptor in mice with apolipoprotein E gene knockout[J].Peptides,2006,27(4):858-863.
[8] 王红霞,曾翔俊,王珏,等.尾加压素Ⅱ及其受体在2型糖尿病小鼠肝脏中表达的变化[J].首都医科大学学报,2008,29(6):741-744.
[9] 周厚荣,张谦,杨秀林.肿瘤坏死因子-α、肾素-血管紧张素系统与机械通气患者胰岛素抵抗相关关系的研究[J].贵州医药,2011,35(12):1074-1076.
[10] Barrette PO,Schwertani AG.A closer look at the role of urotensin Ⅱ in the metabolic syndrome[J].Frontiers in Endocrinology(Lausanne),2012,3:165-174.
[11] Hong Xia Wang,Li Ke Zhang,Chao Shu Tang,et al.Elevated expression of urotensin Ⅱ and its receptor in skeletal muscle of diabetic mouse Regulatory[J].Regul Pept,2009,10:85-90.
[12] Alicia HC,Allison MG,Teresa AZ,et al.Deletion of interleukin-6 improves pyruvate tolerance without altering hepatic insulin signaling in the leptin receptor-deficient mouse[J].Metabolism,2011,60(11):1610-1619.
[13] Janina W,Stefan RJ,Christoph G.Interleukin-6 and its receptors:A highly regulated and dynamic system[J].Cytokine,2014,70(1):11-20.
[14] Birker-Robaczewska M,Binkert C,Nayler O,et al.The expression of urotensin Ⅱ receptor(U2R)is up-regulated by interferon-gamma[J].J Recept Signal Transduct Res,2003,23(4):289-305.
High expression of IL-6 induced the up-regulation of UII/UT system in mouse myogenic cell line C2C12
YanWen1,LiuLixin1,XiaShan2,WangHongxia2△.
1.YanjingMedicalCollege,CapitalMedicalUniversity,Beijing101300,China. 2.ChinesePeople'sLiberationArmy66222ArmyHospital,Beijing102202,China. 3.BasicMedicalCollege,CapitalMedicalUniversity,Beijing100084,China.
Objective To observe the effect of IL-6 on the expression of UⅡ/UT system in C2C12. Methods Cell injury was determined by Placenta blue staining and LDH kit. The content of UⅡ in cell lysate and cell incubation fluid were assayed by RIA, mRNA levels were assayed by RT-PCR. Results UⅡ content in cell incubation fluid was increased after IL-6 of different concentration was administrated, the UⅡ content was increased by 104.9 %(P<0.01), 86.4 %(P<0.01)and 94.3 %(P<0.01) when the concentration of IL-6 was elevated to 10 ng/mL, 50 ng/mL and 100 ng/mL. The simulation of different concentration of IL-6 (0.1 ng/mL, 1 ng/mL, 10 ng/mL and 50 ng/mL) significantly increased the expression of UT mRNA. However, the expression of UT mRNA was decreased significantly when the concentration of IL-6 was elevated to100 ng/mL. Conclusion IL-6 might be a factor involved in up-regulating the expression of UⅡ/UT system for the skeletal muscle of Type 2 diabetes mellitus.
Urotensin Ⅱ G protein coupled receptor; Interleukin-6
北京高等学校青年英才项目(YETP1679);首都医科大学校自然科学基金(2014QD03)
R363
A
1000-744X(2016)03-0233-03
2015-09-21)
△通信作者,E-mail:whxdy112@ccmu.edu.cn
