叶艺兰辐射突变体的气孔特征与染色体核型分析*
2017-01-04苏畅李枝林付永恒李夏媛王玉英
苏畅,李枝林,付永恒,李夏媛,王玉英
(云南农业大学 园林园艺学院花卉研究所,云南 昆明650201)
叶艺兰辐射突变体的气孔特征与染色体核型分析*
苏畅,李枝林,付永恒,李夏媛,王玉英
(云南农业大学 园林园艺学院花卉研究所,云南 昆明650201)
为阐明辐射诱发的叶艺兰植株与未经诱变植株的差异,以经辐射诱发的叶艺兰植株(TRIR-2)与未经诱变的全绿植株(TRIR-1)为试材,运用显微镜观察植物叶片的气孔及根尖细胞的染色体并使用Photoshop软件对染色体进行分析。结果表明,(1)TRIR-2的气孔数量、面积、密度值均小于TRIR-1;(2)TRIR-1植株的染色体核型公式为2n=2x=10m+24sm+6st,属较为对称的“2B”型;TRIR-2植株的染色体核型公式为2n=2x=10m+28sm+2st,属不对称的“2C”型。说明辐射诱变产生的叶艺兰突变体气孔特征和染色体结构发生了变化。
辐射诱变;叶艺兰;气孔;染色体;核型
兰花是中国传统名贵花卉,叶艺兰(线艺兰)是指兰属(Cymbidium)植株叶片上镶嵌着或金黄、或银白、或浓绿、或朱红、或墨黑的嘴、边、点、线、斑的中国兰统称[1]。叶(线)艺品种以墨兰〔Cymbidiumsinense(Jackson ex Andr.)Willd.〕最多,建兰(C.ensifolium)次之,春兰(C.goerigii)、蕙兰(C.faberi)与寒兰(C.kanran)较少[2]。艺象主要表现为:金棱边类、绿艺类、先明后暗类(全斑类)、钳帽子类、爪艺类、水晶类、斑缟类(先明,后明)、棒缟类(先明,后明)、图斑艺类、虎(豹)斑类等[3]。叶艺兰栽培已有千年,因其叶片艳丽,形态优美,被誉为“花中君子”,是珍贵的观赏花卉[4];因其奇花、奇叶和矮种等各种变异类型层出不穷,市场前景十分广阔。
辐射诱变育种是一种新型育种方法,具有创造多种突变体、丰富种质资源库,打破基因连锁、提高重组率,保持优良性状、改良品种的某些不良性状,以及突变性状稳定快、缩短育种年限等特点,γ射线是植物诱变育种中最常用的辐射诱变因素[5]。目前,辐射诱变技术已在月季(RosachinensisJacq.)、菊花〔Dendranthemamorifolium(Ramat.) Tzvel〕、美人蕉(CannaindicaL.)、大丽花(DahliapinnataCav.)、叶子花(BougainvilleaspectabilisWilld.)、荷花(Nelumbonucifera)植物上获得60多种突变体[6]。应用60Coγ射线处理碧玉兰〔Cymbidiumlowianum(Rchb.f.)Rchb.f.〕、西藏虎头兰(C.tracyanumL.)、冬凤兰(C.dayanumRchb.f.)、竹叶兰〔Arundinagraminifolia(D.Don)Hochr.〕的组培苗以及墨兰[7]和兰花春剑隆昌素(C.longibracteatumcv.‘longchangsu’)根状茎[8]已获得了植株矮化、变粗、叶变宽、叶上有淡绿斑、绿芽分化和叶色失绿变淡等表型变化。已有研究表明,兰花经辐射诱变后引起一系列基因突变,进而使兰花的气孔性状发生变化[9]。此外,辐射可诱发染色体数量、结构和行为畸变,重组体的产生、染色体断裂、结构重排,以及引起染色体行为变异,如染色体桥的出现、染色体落后等[10]。
为指导兰属其他种和变种的人工叶艺育成,本研究以经辐射诱发的叶艺兰与未经诱变的全绿植株为试材,通过对植物叶片气孔和根尖染色体的观察,从细胞学水平分析两种植株的差异,以期为阐释叶艺形成的机理奠定基础,为其他科、属观叶植物人工诱发叶艺的研究提供参考。
1 材料与方法
1.1 材料
2004年,本课题组获得素花虎头兰(C.hooke-rianum)与黄蝉兰(C.iridioides)F1杂交苗,并筛选出性状稳定、一致的株系;2005年7月,将性状稳定、一致的F1杂交种苗在云南华源辐射有限公司进行60Coγ射线急性照射。培育2年,发现数株叶艺品系;经过6年的继代培养,获得特异性状稳定的叶艺新品系2种。一种类型的叶尖和叶边缘为绿色、其他部分为白色的叶艺株系,是叶艺部分观赏价值较高、叶形较为优美的新品系,命名为TRIR-2;另一种类型为全绿叶片,即未出现叶艺的新品系,命名为TRIR-1。本研究供试材料即为TRIR-2和TRIR-1的组培苗(图1)。

TRIR-1植株TRIR-2植株TRIR-1植株与TRIR-2植株
图1 TRIR-1和TRIR-2植株
Fig.1 TRIR-1 and TRIR-2
1.2 研究方法
1.2.1 气孔观察
取TRIR-1和TRIR-2各3株观察植株的叶片,选取每一株植株中最长的叶片观察,每一片叶片平均分成上、中、下 3部分,撕取叶片下表皮,在显微镜下观察,每个样观测3个视野。用Photoshop处理图片并记下每张图片中的气孔数,用Photoshop计算每个气孔的面积和整个视野面积,并计算其平均值。气孔的面积(μm2)=长径×短径。
1.2.2 染色体分析
中国兰花通常是自养型的地生草本植物,具有粗大的根系,且被海绵状白色的根被所覆盖,中央有1个维管组织的髓部。因此,对兰花根部进行染色体制片较为困难。在对染色体制片各个环节进行比较后,选择出效果相对较好的制片方案[11~12]。
取TRIR-1和TRIR-2各3株观察植株的根尖,每1株植株中选取3个根长0.5cm左右的根尖,取根尖分生组织,在显微镜下观察。
(1)预处理 将取出的大小为0.5cm的根尖组织加入0.1%的秋水仙素预处理液[13~14],于常温下预处理约4h,洗涤后转入Carony’s液(乙醇︰乙酸=3︰1)中固定30min,取出后放入60℃浓度为1mol/ L的HCl中水解10min。
(2)染色 用蒸馏水清洗3~5次,加入1~2滴的卡宝品红染色30min。
(3)压片 将完成染色的根尖放于载玻片上,盖上盖玻片,橡皮头轻敲团状物,使细胞均匀分散。
(4)镜检 将制作完成的装片置于显微镜下观察,选择染色体形态、分散较好的根尖装片进行拍照。
(5)染色体核型分析 用Photoshop从原始图中解析出染色体,根据目测进行同源染色体配对、排序;然后按从大到小的顺序进行同源染色体排序,使用变形工具对染色体的位置进行调整,使其短臂朝上,长臂朝下,使所有染色体主缢痕尽量在同一水平线上[15~16]。
核型分析采用李懋学等的核型标准进行[17],染色体的相对长度、臂比及类型按Levan等的命名系统,核型分类按照Stebbins的标准划分[18]。利用Photoshop版本辅助分析。
1.3 数据分析
方差分析采用DPS软件,染色体核型分析采用Photoshop软件。
2 结果与分析
2.1 气孔特征分析
由表1和图2可知,TRIR-1、TRIR-2白色区和TRIR-2绿色区的气孔数量、气孔面积和气孔密度均存在差异,但三者之间的气孔数量差异不显著(P>0.05),而TRIR-1的气孔面积显著高于TRIR-2白色区和TRIR-2绿色区(P<0.05)。说明辐射诱发了叶艺兰的叶片气孔数量、面积和密度的变化。
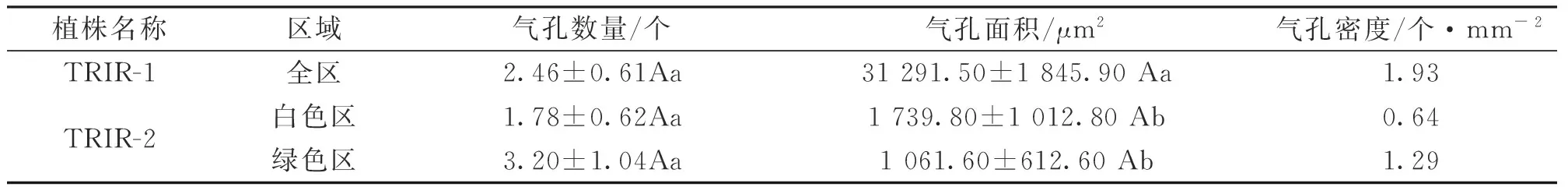
表1 TRIR-1和TRIR-2植株的气孔特征
注:小写字母不同表示差异显著(P<0.05),大写字母不同表示差异极显著(P<0.01),下同。

TRIR-1全区TRIR-2白色区TRIR-2绿色区
图2 TRIR-1和TRIR-2植株的植株气孔(×10)
Fig.2 Stomata of TRIR-1 and TRIR-2 (×10)
2.2 染色体核型分析
由表2、表3、图3和图4可知,TRIR-1的m(中着丝粒染色体,下同)染色体有10个,sm(近中着丝粒染色体,下同)染色体有24个,st(近端着丝粒染色体,下同)染色体有6个;TRIR-2的m染色体有10个,sm染色体有28个,st染色体有2个;TRIR-1的近中部染色体数比TRIR-2少4个,TRIR-1的近端部染色体数比TRIR-2多4个;TRIR-1的染色体长度变化范围和最长染色体与最短染色体值比均大于TRIR-2;TRIR-1的核型公式为2n=2x=10m+24sm+6st,属“2B”型;TRIR-2的核型公式为2n=2x=10m+28sm+2st,属“2C”型。说明与TRIR-2相比,TRIR-1具有较对称的核型,辐射诱发叶艺兰的染色体核型发生了变化,进而导致叶艺兰突变体的形成。

表2 TRIR-1和TRIR-2植株的染色体参数

TRIR-1的根尖染色体TRIR-2的根尖染色体

图3 TRIR-1和TRIR-2的染色体(×100)
图4 TRIR-1和TRIR-2植株核型模式
Fig.4 Karyotypic of TRIR-1 and TRIR-2
3 结论与讨论
3.1 结论
辐射诱变导致叶艺植株气孔的数量、面积和密度的变化;同时染色体核型由较为对称的“2B”型转为不对称的“2C”型。表明辐射诱发了叶艺兰植株(TRIR-2)变异体的产生。
3.2 讨论
目前研究发现影响气孔发育的因素有基因[19]、激素[20]、微管和环境因子4方面[21]。其中基因控制着气孔的发生;脱落酸、赤霉素和乙烯等激素影响气孔的产生。本试验结果表明辐射诱发叶艺兰植株的气孔数量、面积和密度的变化,说明辐射能够影响植物气孔的发育。相关研究表明突变株系的叶片气孔密度增大,气孔面积减小,气孔分布更密,气孔更小,有利于水分的蒸腾扩散[22]。本研究中气孔面积缩小与前人研究结果一致,但辐射导致叶艺兰植株叶片中白色区域气孔数量降低,绿色区域气孔数量增加,这可能是由于叶艺兰植株为了能够生存,当白色区域气孔数量较少时,绿色部分的气孔数量就会增多,使叶艺植株的气孔在光合、呼吸、蒸腾等作用中成为空气和水蒸汽的通路,以此来满足叶艺兰植株的生长需求。
相关研究表明菊花花色的辐射诱变导致菊花染色体结构的变化,使基因发生缺失、重复、倒位、易位等现象[23]。菊花的花色变化是染色体的核型经过辐射之后由对称变为不对称所致,同时染色体进化与生物进化有着极其密切的关系[24]。本研究结果也表明了辐射诱变导致叶艺兰植株染色体核型结构的变化。研究结果对整个植物核型进化方式的研究提供了新的证据。高等植物核型进化的基本趋势是由对称向不对称方向发展,系统演化上处于比较古老或原始的植物,往往具有较对称的核型,不对称的核型通常出现在较进化或特化的植物中[25~26]。李玉阁等[27]报道的7种兰亚属植物的核型,结果也表明兰属的染色体数目保守,除了兔耳兰(C.lancifolium)和线艺春兰外,其余染色体数目均为2n=40,且染色体的结构主要由m和sm染色体组成,核型具有较高的对称性,为“2A”或“2B”型。金兰(Cephalantheraerecta)、银兰(Cephalantherafalcata)的染色体主要由sm和st染色体组成,核型均属较不对称的“3C”型,且为二型式核型,即1~3对为大型染色体,而4~17却为小型染色体[27]。本研究表明TRIR-1植株的核型公式是2n=2x=10m+24sm+6st,属较为对称的“2B”型;TRIR-2植株的核型公式是2n=2x=10m+28sm+2st,属不对称的“2C”型。说明通过辐射诱变的叶艺兰植株的遗传物质发生了变化,导致核型由对称向不对称方向发展,这与高等植物核型进化的基本趋势一致。
[1]李丽,罗君琴,聂振鹏,等.线艺兰的组织培养及植株再生研究[J].浙江农业科学,2008,6(1):679-681.
[2]林建院.线艺兰及其栽培技术[J].广东园林,1999,3(1):35-38.
[3]陈少宏.试论国兰的遗传变异与选育[J].广东园林,1999,4(1):34-36.
[4]卢思聪.中国兰与洋兰[M].北京:金盾出版社,1994:34-39.
[5]王元东,赵久然,郭景伦,等.诱变育种在创造玉米新种质中的应用[J].北京农业科学,1999,17(2):2-16.
[6]陈子元.从辐射育种的发展来展望航天育种的前景[J].核农学报,2002,16(5):261-265.
[7]傅雪琳,张志胜,何平.60Coγ射线辐射对墨兰根状茎生长和分化效应研究[J].核农学报,2000,14(6):333-336.
[8]蒋彧,何俊蓉,刘菲,等.60Coγ辐射兰花春剑隆昌素根状茎分化苗的ISSR分析[J].核农学报,2013,27(9):1247-1252.
[9]赵燕,洪亚辉,刘清波.菊花辐射后代部分器官维管束与气孔的解剖观察[J].湖南农业大学学报(自然科学版),1999,29(6):468-470.
[10]潘宏.兰花辐射诱变与组织培养技术初步研究[D].福州:福建农林大学,2008.
[11]杨宁,谈永霞,李巧峡,等.百里香染色体制片优化及核型分析[J].草业学报,2012,21(1):184-189.
[12]申建双,叶远俊,潘会堂,等.12份连翘种质资源的核型参数分析[J].植物遗传资源学报,2015,16(1):178-184.
[13]杨玲,罗梦婷.不同预处理对长寿沙田柚根尖染色体制片的影响[J].南方农业,2012,6(6):73-76.
[14]周翼虎,霍秀文,刘向宇,等.山药染色体制片技术的优化[J].中国园艺文摘,2014,8(1):7-10.
[15]蒋姗姗,梁英民,王作军.利用个人电脑系统及photoshop软件进行核型分析[J].第四军医大学学报,2000,21(7):860-865
[16]杨大翔.用A Dobe Photoshop进行核型分析[J].农业网络信息,2005,3(1):44-45.
[17]李懋学,陈瑞阳.关于植物核型分析标准化问题[J].武汉植物学研究,1985,3(4):298-301.
[18]Stebbins G L.Chromosomal evolution in higher plants[M].London:Edward Arnold Ltd,1971:10-88.
[19]赵益超,公华林,宋开侠.气孔发育的研究进展[J].现代农业科技,2008,11(1):361,366.
[20]周丽娟,陈尔娟,韩笑,等.激素与气孔发育研究进展[J].西北植物学报,2015,35(4):845-851.
[21]高素娟,张丽丽,王小菁.微管动态不稳定性及其微管排向的光调控[J].天水师范学院学报,2009,29(5):16-19.
[22]韩蕾.太空环境对草地早熟禾的诱变效应及其诱成突变体的生物学变化[D].北京:中国林业科学研究院,2005.
[23]洪亚辉,朱兆海,黄璜,等.菊花组织培养与辐射诱变的研究[J].湖南农业大学学报,2003,29(6):459-460.
[24]姚世鸿.染色体进化与生物进化关系研究的历史与现状[J].贵州师范大学学报,1993,4(1):2-5.
[25]Stebbins G L.Adaptive radiation of reproductive characteristics in angiosperms,II:seeds and seedlings[J].Annual Review of Ecology and Systematics,1971,35(4):237-260.
[26]李桂芬,梁国鲁,林顺权.枇杷属植物核型分析及其在系统分类中的应用[J].园艺学报,2013,40(8):1465-1474.
[27]李玉阁,郭卫红,吴伯骥.4种国产兰属植物的核型比较研究[J].西北植物学报,2002,22(6):1438-1444.
Leaf Stomatal and Chromosome Karyotype of Ye-yi Orchid Mutant Induced by Radiation
SU Chang,LI Zhi-lin,FU Yong-heng,LI Xia-yuan,WANG Yu-ying
(Horticulture and Landscape College of Yunnan Agricultural University,Kunming Yunnan 650201,P.R.China)
In order to expound the differences of Ye-yi orchid that was induced by irradiation and that was not induced,the leaf stomatal and the chromosome of root tip cell were observed by microscope,and the chromosome were further analyzed by Photoshop software.The results showed that,(1) quantity,stomatic size and density of stomata of induced plant (TRIR-2)were less than plant that was not induced (TRIR-1);(2) karyotype formula of TRIR-1 is 2n=2x=10m+24sm+6st,a relatively symmetrical 2B type, and karyotype formula of TRIR-2 is 2n=2x=10m+28sm+2st,an asymmetric 2C type.
radiation induced;orchid with verge line pattern leaves (Ye-yi orchid); stomatal ;chromosome;karyotype
10.16473/j.cnki.xblykx1972.2016.06.016
2015-10-29
国家自然科学基金项目(30160074),云南省自然科学基金重点项目(2002C0003P),云南省重点新产品开发项目(2012BB008),
苏畅(1989-),女,研究生,主要从事园艺植物资源的利用及创新研究。E-mail:515718100@qq.com
简介:王玉英(1980-),女,讲师,主要从事植物资源的利用和创新研究。E-mail:wyysxp@126.com
S 682.31
A
1672-8246(2016)06-0089-06
云南农业大学园林园艺学院院级基金(2014001),云南省昆明市科学技术局重点项目(2015-1-N-00984)。
