低温层积过程中野鸦椿种子生理生化变化的研究
2016-12-19张莉梅张子晗喻方圆
张莉梅 ,张子晗 ,喻方圆
(1.南京林业大学 南方现代林业协同创新中心 林学院,江苏 南京210037;2.北京林业大学 自然保护区学院,北京 100083)
低温层积过程中野鸦椿种子生理生化变化的研究
张莉梅1,2,张子晗1,喻方圆1
(1.南京林业大学 南方现代林业协同创新中心 林学院,江苏 南京210037;2.北京林业大学 自然保护区学院,北京 100083)
通过对低温层积不同时间野鸦椿种子内含物质变化情况,及不同播前预处理种子发芽情况的研究,得出如下结论:(1)野鸦椿种子不通过低温层积处理无法萌发。(2)野鸦椿种子在层积过程中主要通过大分子油脂类物质水解作种子萌发的物质动员。(3)野鸦椿种子通过低温层积的方法解除休眠的时间应控制在90~120 d内,层积时间过长发芽率反而下降。这与同属植物圆齿野鸦椿种子的萌发特性一致。(4)层积90~120 d时种子内可溶性糖含量有较大幅度下降,同时在120 d时种子的发芽率达到峰值,表明可溶性糖是野鸦椿种子萌发的直接利用能源物质。
野鸦椿种子;休眠;低温层积;发芽;储存营养物质
野鸦椿Euscaphis japonica为省沽油科、野鸦椿属落叶小乔木或灌木,是一种兼具工业、药用及观赏价值的树种。由于其果实多被一层形状奇特,色彩鲜艳的外种皮包被,结果丰硕而满足园林绿化与美化功能[1]。同时野鸦椿根微苦,果辛温,为药品血脉通胶囊中的一味主要药材[2]。干果有温中理气,消肿止痛的功效。主治胃痛、泻疾等。根可祛风除湿,健脾调营。花可治头痛眩晕,是一种良好的中草药材兼具优良绿化树种[3],种子油可制皂[4]。
野鸦椿种子近圆形,直径3~5 mm,内种皮结构木质、坚硬,且具一层黑色膜质外种皮,因此可能存在机械障碍和透性障碍及阻止内源抑制物质外渗。同时通过对相关文献的查阅,野鸦椿属种子具有明显休眠特性,种子需催芽处理才能保证其发芽率[5-7]。因此我们在种子低温层积催芽处理一段时间后展开相关试验工作。
近年来在种子休眠与萌发影响方面的研究取得了很大进展,但这些研究很多都局限于一些模型植物,如:拟南芥、谷类植物、烟草等。大量的非模式植物,特别是一些自然萌发率低下的木本植物种子的休眠与萌发调节机制研究方面尚十分薄弱,急待开展深入研究[8]。而对于野鸦椿的研究主要集中于其药理属性方面,并未从野鸦椿种子萌发过程中内含物质变化机制的角度来探索萌发过程中内含物质与种子发芽率的关系。本文旨在探索打破野鸦椿种子休眠的方法及低温层积过程中种子内营养物质变化情况。
1 材料与方法
1.1 材料来源
供试种子为2013年9月20日采自江苏省镇江市南京林业大学下蜀林场当年新鲜野鸦椿种子。补充实验种子于2014年10月采自江苏省镇江市南京林业大学下蜀林场。
1.2 材料处理与测定
采取低温层积加播前80℃温水浸泡的方法对种子进行预处理。于2013年9月20日起对野鸦椿种子进行低温层积处理,基质为洁净河沙,以体积比3份湿河沙与1份种子混合,在4℃条件下冷藏。试验期间适时喷水,保证基质处于湿润状态。当年12月20日第1次取样,之后每30 d取1次样,共取5次,于2014年4月20日取样结束。取样种子一部分立即做发芽试验,另一部分用-70℃超低温冰箱保存,用于生理指标测定。同时,实验组于2014年9月第二次采种,以获取未进行低温层积处理时,种子的发芽情况与生理状态。
1.2.1 野鸦椿种子发芽试验
将不层积或不同层积时间的种子分为处理组和对照组。处理组种子用80 ℃温水浸泡24 h,自然冷却,对照组种子冷水浸泡。处理组与对照组均4次重复,每重复50粒种子。将种子置于发芽盒内,基质为湿河沙,于24 h光照、25 ℃恒温光照培养箱内培养1个月后统计发芽情况。期间适时喷水,使基质保持湿润状态,为种子萌发提供充足的水分供应。
1.2.2 野鸦椿种子层积过程中内含物质含量测定
每次取样后,测定种胚中可溶性糖含量、淀粉含量、粗脂肪含量以及可溶性蛋白质含量。实验中通过敲碎种壳而获取完整种胚的方法,以避免种壳中营养物质对实验结果进行干扰。
可溶性糖含量和淀粉含量测定采用蒽酮比色法[9]、可溶性蛋白质含量测定采用考马斯亮蓝法、粗脂肪含量测定采用索氏提取法[10]。
数据采用Microsoft Excel 2007和SPSS18.0软件进行图表处理和统计分析。
2 结果与分析
2.1 种子发芽率的变化
种子的发芽率是对种子休眠研究的重要参照指标,其结果可较直观的衡量种子休眠的解除程度。因此,本实验用种子的发芽率来描述种子休眠的解除程度,试验结果如表1。
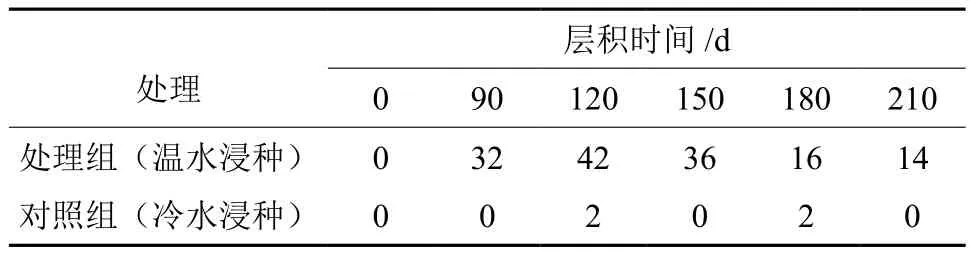
表1 低温层积不同时间+温水浸泡处理对种子发芽率的影响Table 1 Effects of low temperature stritification at different length of time plus soaking with warm water treatment on germination rate of Euscaphis japonica seeds %
表1可见,在种子低温层积解除休眠过程中,随着层积时间的增加,对照组种子的发芽率很低,平均发芽率为0%。同时补充实验种子中,不经过低温层积处理时,处理组即温水浸泡亦不能使种子正常萌发。
通过对实验数据的分析,低温层积120 d时发芽率达峰值42%,而层积150、180和210 d时,发芽率分别下降了14.3%、61.9%和66.7%。
2.2 种子层积过程中生理生化变化
2.2.1 可溶性糖与淀粉含量的变化
由图1可知,层积90~120 d,可溶性糖含量有小幅度下降,而在层积120~150d时,可溶性糖含量有大幅上升,150 d之后,可溶性糖含量小幅下降。低温层积90~210 d这一时间段内,种子内可溶性糖含量的变化不大,此时可溶性糖含量相对稳定。方差分析表明,不同层积时间种子可溶性糖含量差异极显著(见表2)。
由图2可知,在层积90~210 d的过程中,野鸦椿种子中淀粉含量逐渐小幅上升,而层积90~210 d期间,种子中淀粉含量相对稳定。方差分析表明,不同层积时间种子淀粉含量差异不显著(见表2)。
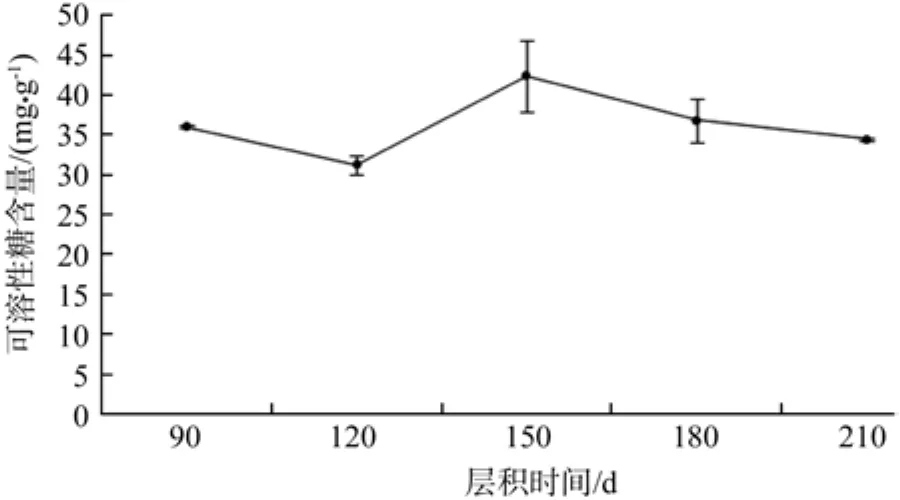
图1 低温层积过程中可溶性糖含量的变化Fig. 1 Change of soluble sugars during the low temperature strati fi cation
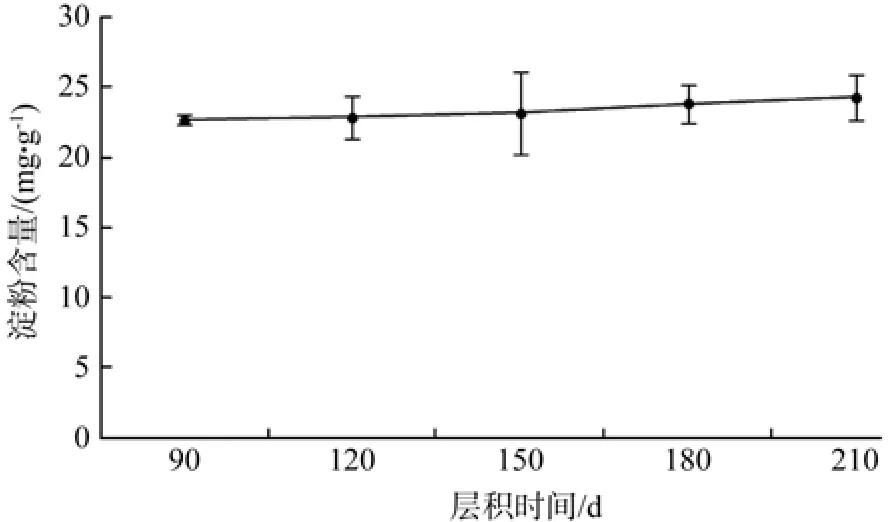
图2 低温层积过程中淀粉含量的变化Fig. 2 Change of starch during the low temperature strati fi cation
2.2.2 可溶性蛋白质与粗脂肪含量的变化
由图3可知,层积过程中,野鸦椿种子中可溶性蛋白质的含量在90~150 d时大幅上升,而在150~210 d时小幅上升趋于平缓。90~150 d内可溶性蛋白质含量上升了32%,而150~210 d内可溶性蛋白质含量上升了2%。方差分析表明,不同层积时间种子可溶性蛋白质含量差异极显著(见表2)。
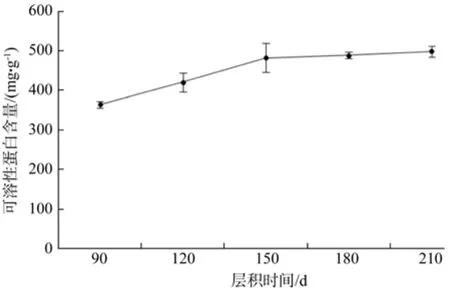
图3 低温层积过程中可溶性蛋白质含量的变化Fig. 3 Change of soluble protein during the low temperature strati fi cation
由图4可以看出,野鸦椿种子低温层积过程中,粗脂肪含量呈下降趋势,层积90 d时,粗脂肪含量占干重的10.89%;层积210 d时,粗脂肪含量占干重的8.43%。粗脂肪含量下降了22.59%。方差分析表明,不同层积时间种子粗脂肪含量差异显著(见表2)。
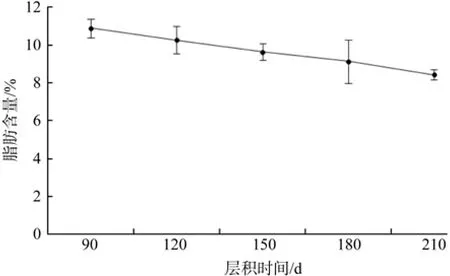
图4 低温层积过程中粗脂肪含量的变化Fig. 4 Change of crude fat during the low temperature strati fi cation
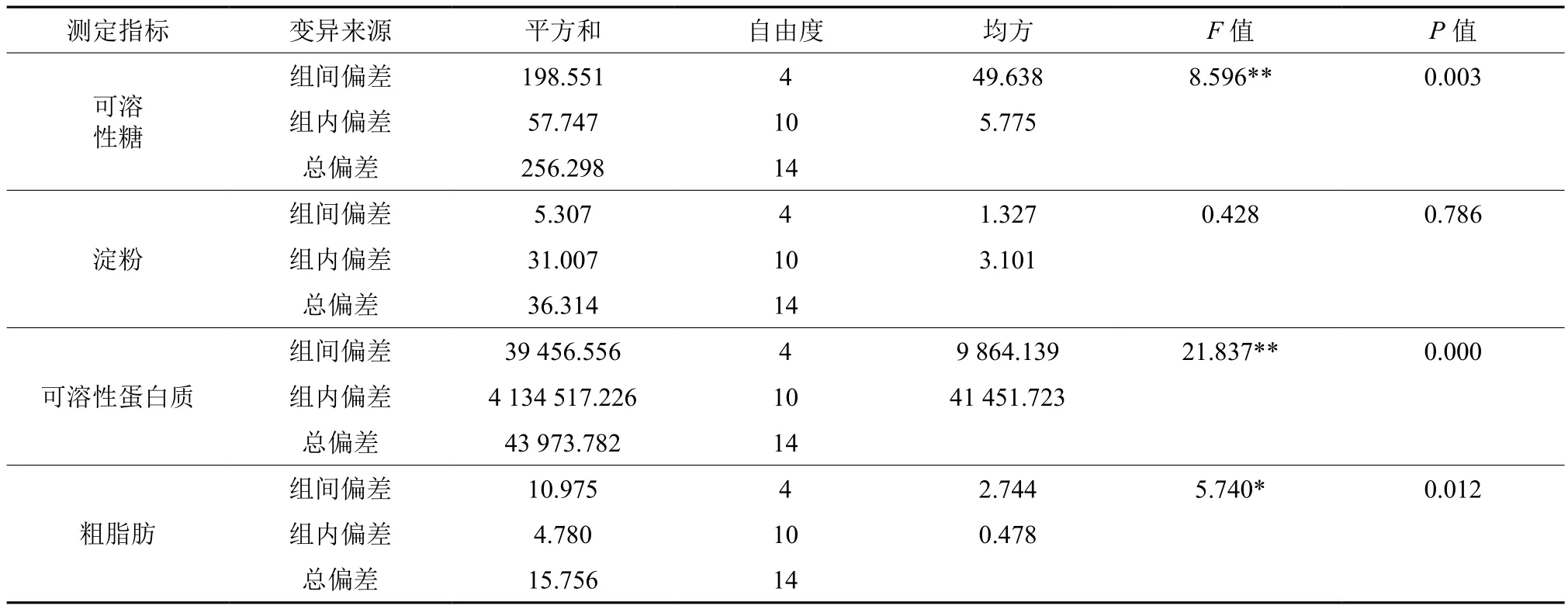
表2 低温层积过程中野鸦椿种子可溶性糖,淀粉,可溶性蛋白质,粗脂肪含量方差分析†Table 2 ANOVA on content of soluble sugars, starch, soluble protein and crude fat during low temperatue stratification of Euscaphis japonica seeds
3 结论与讨论
3.1 野鸭椿种子休眠的原因
休眠是种子植物抵抗外界不良环境的一种适应性,并且是随着种子的发育、成熟而逐渐形成[11]。陈昕等[12]探索黄山花楸种子休眠影响因素的实验中发现,种皮机械障碍是造成黄山花楸种子休眠的重要因素;洑香香等[13]实验发现,山茱萸种子既有种皮的机械束缚作用,还存在胚的形态休眠,属复杂的多因素综合休眠。南方红豆杉种子角质化且坚硬致密,经酸蚀处理能很快吸水膨大,而未处理时,吸水速度较慢[14-15]。在东京野茉莉种子休眠机制的探索中,休眠的原因主要包括种皮障碍、缺少萌发所需激素以及种皮、胚中存在萌发抑制物, 其中种皮障碍和抑制物的存在是限制种子萌发的首要因素[16]。野鸦椿种子种壳硬且浓,密度大,很难吸水膨胀,所以播种催芽难度很大[17]。未经低温层积处理的野鸦椿种子,无论是否通过80℃温水浸种,均未能发芽,说明此时野鸦椿种子还不具备发芽条件,不能正常发芽。低温层积处理210 d后不进行其他预处理情况下种子的发芽率仍极低,说明仅低温层积处理并不足以解除野鸦椿种子的休眠现象。低温层积后用温水浸泡,能显著提高种子发芽率。说明野鸦椿种子的种皮对种子的萌发存在较强的机械阻碍作用,但发芽率仍未达较高水平,因此推断可能还存在其他抑制物质抑制种子萌发。综合来看,野鸦椿种子属于综合休眠类型,一是存在种皮障碍,二是存在抑制物质。这与同属植物圆齿野鸦椿种子的休眠类型一致[18]。
3.2 内含物质含量变化对种子萌发的影响
3.2.1 野鸦椿种子内可溶性糖含量与淀粉含量变化
植物种子种胚中的淀粉、油脂是种子萌发过程中重要的储能和供能物质,而可溶性糖含量水平标志着源端的同化物供应能力,反映出库端对同化物的转化、利用能力[19]。张艳杰[14]实验表明,南方红豆杉种子层积过程中可溶性糖含量大体上呈现出先增加再减少的趋势。本实验数据显示,可溶性糖含量在120~150 d时有较大幅度上升,90~120 d、150~180 d、180~210 d时,可溶性糖含量均下降,这可能是由于在120 d时,种子消耗大量的可溶性糖以萌发,在层积120~150 d期间种子内淀粉、脂类等部分大分子储藏物质分解予以补充种子内可溶性糖,通过代谢作用促进种子萌发,因此此阶段可溶性糖含量有较大幅度的上升,同时层积120 d时,种子发芽率最高。
3.2.2 野鸦椿种子内可溶性蛋白质含量变化
实验中随着低温层积时间的增加,可溶性蛋白质的含量也在上升,说明90~210 d这一阶段大分子蛋白质不断水解为可溶性蛋白质,为种子的萌发提供所需基本物质。同时,90~150 d时,可溶性蛋白质含量上升较快,幅度达32%;150~210 d时,含量上升较慢,幅度为2%。可能是由于90~150 d这一阶段蛋白质的水解更剧烈。
3.2.3 野鸦椿种子内粗脂肪含量变化
野鸦椿种子随着层积时间的增加,脂肪的总含量呈下降的趋势,低温层积90 d~210 d期间,粗脂肪含量下降了23%,结果表明,野鸭椿种子在低温层积过程中,脂肪总含量不断减少,这可能是为种子的萌发提供必要的能量物质,如可溶性糖等小分子可直接利用物质,在种子萌发层积过程中通过大分子油脂类物质水解作萌发的物质动员。张艳杰[14]实验发现,南方红豆杉种子在层积过程中粗脂肪含量呈逐渐下降趋势。黄儒珠等[20]认为脂类物质的分解、转化和利用可能是南方红豆杉种子休萌解除的关键。笔者认为脂类物质的分解与转化为野鸦椿种子解除休眠过程提供能量,是种子萌发过程中的主要供能物质。野鸦椿种子含油量高,种子油可制皂[4], 笔者认为,由于脂肪类物质对于种子萌发起关键作用,种子油的利用应为及时采种及时制油[21]。
3.3 种子最适层积时间的探讨及引起种子发芽率降低的因素
由表1可知,野鸦椿种子低温层积120 d后,处理组种子的发芽率下降了14.3%,说明仅通过温水浸种破除种皮障碍的方法来处理野鸦椿种子时,低温层积时间应控制在90~120 d,层积时间不宜过长。同时,课题组在2013年进行的低温层积打破同属植物圆齿野鸦椿种子休眠的实验中也发现,在低温层积超过120 d时,圆齿野鸦椿种子的发芽率大幅下降[18],因此笔者认为,层积时间的长度与野鸦椿属植物种子萌发特性的关系有一定研究意义。值得一提的是,在低温层积过程中,IAA、ABA、GA3、ZT等种子内源激素含量的变化情况已在部分木本植物中进行了研究讨论[22-24],但野鸦椿属种子的相关研究目前还是空白,而对于发芽率下降的问题,可通过研究内源激素含量的变化来做进一步探究。
[1] 龚敏元.观果树种野鸦椿在泰宁县的绿化应用[J].中国城市林业,2014,12(5):36-38.
[2] 罗海羽,姚 默,申万祥,等.野鸦椿属药学研究概况[J].安徽农业科学,2012,40(12): 8462-8463.
[3] 葛玉珍.野鸦椿资源及其利用[J].中国野生植物资源,2004,23(5): 24-25.
[4] 龙春林,宋洪川.中国柴油植物[M].北京:科学出版社,2012: 158-159 .
[5] Bewley J D, Black M. Physiology of Development and Germination [M]. New York: Plenum Press, 1994.
[6] 欧 斌,李远章.圆齿野鸦椿种子预处理和苗木生长规律及育苗技术研究[J].江西林业科技,2006(3):16-18.
[7] 覃嘉佳,龙云英.圆齿野鸦椿扦插繁殖技术[J].林业科技开发,2007,21(3): 71-73.
[8] 程 鹏,王 平,孙吉康,等.植物种子休眠与萌发调控机制研究进展[J].中南林业科技大学学报,2013,33(5):52-58.
[9] 黄学林,陈润政.种子生理试验手册[M].北京:中国农业出版社,1998.
[10] 刘正祥,张华新,刘 涛.省沽油种子油脂分析与功能特性评价[J].林业科学,2008,44 (2):48-54.
[11] 廖卓毅,钱存梦,马秋月,等.乌桕种子成熟过程中种皮和胚乳超微结构观察[J].南京林业大学学报:自然科学版,2014,38(6): 43-47.
[12] 陈 昕,曹珊珊,张红星.黄山花楸种子休眠影响因素[J].东北林业大学学报,2010,38(7):5-7.
[13] 洑香香,周晓东,刘红娜.山茱萸种子休眠机理与解除方法初探[J].中南林业科技大学学报,2013,33(4):7-12.
[14] 张艳杰. 南方红豆杉种子休眠机理的研究[D].南京林业大学,2007.
[15] 鲁定伟,周要全,李曙波,等.南方红豆杉生物学、生态学特性及培育技术[J].经济林研究,2014,32(4):159-164.
[16] 许晓岚,丁芳芳,李 翔,等.东京野茉莉种子休眠机制及其破除方法初探[J].西北植物学报, 2012,32(11):2270-2278.
[17] 杨燕凌.打破圆齿野鸦椿种子休眠及外植体选择诱导实验研究[D].福建农林大学,2008.
[18] 张莉梅,张子晗,刘 源,等.低温层积过程序中圆齿野鸦椿种子的生理生化变化[J].种子,2015,34(7):37-40.
[19] 郝丽丽,张存旭,杨 阳.栓皮栎种胚发育过程中储藏物质积累变化研究[J].西北林学院学报,2011,26(4):128-131.
[20] 黄儒珠,郭祥泉,方兴添,等.变温层积处理对南方红豆杉种子生理生化特性的影响[J]. 福建师范大学学报:自然科学版,2006, 22(2):95-98.
[21] 王湘南,陈永忠,伍利奇,等.油茶种子含油率和脂肪酸组成研究[J].中南林业科技大学学报,2008,28(3):11-17.
[22] 杨立学,王海南,张 琳,等.低温层积过程中紫椴种子内源激素含量的变化[J].经济林研究,2012,30(2):15-18.
[23] 李冬林,张亚楠,金雅琴,等.毛梾种子低温层积过程中内源激素的变化及其与发芽的关系[J].经济林研究,2014,32(3):36-41.
[24] 金雅琴,李冬林,黄雪方.梾木种子低温层积过程中内源激素含量的动态变化特征[J].西北植物学报,2014,34(11):2255-2261.
Biochemical and physiological changes of Euscaphis japonica seeds during the period of strati fi cation
ZHANG Li-mei1,2, ZHANG Zi-han1, YU Fang-yuan1
(1. Collaborative Innovation Center of Sustainable Forestry in Southern China, College of Forestry, Nanjing Forestry University, Nanjing 210037, Jiangsu, China; 2. School of Nature Reserve,Beijing Forestry University, Beijing 100083, China)
Changes of reserved nutrients in Euscaphis japonica seeds during the process of stratification were studied, and seed germination characteristics after different pretreatments were investigated in this paper. The results were shown as following: (1)Seeds are unable to germinate under no low temperature strati fi cation. (2)Seed germination of Euscaphis japonica were primary mobilized by hydrolysis of macromolecular grease-like substance in the process of strati fi cation. (3) The strati fi cation time should be controlled within the 90~120 d, longer strati fi cation time would make the rate of germination declined. The conclusions was the same as the result of Euscaphis konishii Hayata seeds, which belongs to the same genus as Euscaphis japonica seeds. (4)During the strati fi cation period of 90~120 d, the content of soluble sugar declined signi fi cantly, as the same time, seeds germination rate arrived at peak in the 120 d,showing that soluble sugar was directly used for energy source of seed germination.
Seeds of Euscaphis japonica; Dormancy; Low temperature strati fi cation; Germination; Reserved nutrients
S723.1+31.1
A
1673-923X(2016)11-0036-05
10.14067/j.cnki.1673-923x.2016.11.007
2015-05-09
江苏高校优势学科建设工程资助项目(PAPD);国家林业局林业科技成果推广项目“南方珍贵用材树种苗木高效培育技术示范与推广”
张莉梅,硕士研究生 通讯作者:喻方圆,教授;E-mail:fyyu@njfu.edu.cn
张莉梅,张子晗,喻方圆.低温层积过程中野鸦椿种子生理生化变化的研究[J].中南林业科技大学学报,2016,36(11):36-40.
[本文编校:吴 毅]
