GA3对‘石硖’龙眼假种皮糖积累及蔗糖代谢相关酶的影响
2016-12-17刘丽琴李建光王一承武红霞决登伟石胜友
刘丽琴,李建光,舒 波,王一承,武红霞,决登伟,石胜友*
(1中国热带农业科学院南亚热带作物研究所,农业部热带果树生物学重点实验室,广东 湛江 524091;2广东省农业科学院果树研究所,广东 广州 510640)
GA3对‘石硖’龙眼假种皮糖积累及蔗糖代谢相关酶的影响
刘丽琴1,李建光2,舒 波1,王一承1,武红霞1,决登伟1,石胜友1*
(1中国热带农业科学院南亚热带作物研究所,农业部热带果树生物学重点实验室,广东 湛江 524091;2广东省农业科学院果树研究所,广东 广州 510640)
为明晰外源赤霉素对龙眼果实发育与糖分形成的影响。本实验以‘石硖’龙眼为材料,测定外源GA3处理与对照龙眼果实生长发育指标、假种皮糖分含量及糖代谢相关酶活性水平,分析外源GA3在龙眼果实发育及糖分累积中的调控作用。果实生长发育指标表明,GA3处理增加了果实纵横径、重量及体积并显著增加了假种皮的重量。假种皮糖分水平及酶活性数据则显示,石硖龙眼假种皮以积累蔗糖为主且蔗糖代谢酶AI和SS(分解方向)在蔗糖累积中起主要作用。GA3处理并不影响糖的积累类型,但调控了蔗糖代谢相关酶活性的大小和变化趋势。
GA3;果实发育;糖积累;蔗糖代谢相关酶
龙眼属于无患子科龙眼属植物,是我国南亚热带特产水果。龙眼果实的糖含量和种类是决定果实品质和风味的重要因子,而糖分的积累和代谢离不开蔗糖代谢相关酶的调控。外源GA3是一种高效的植物生长促进物质,可有效调控植物的生长发育。生产上,已在调控果实无核化[1]、果实发育[2-3]、单性结实[4]、花发育[5]、光合作用[6]等方面广泛应用,且有研究表明GA3对葡萄[3]、梨[11]、骏枣[12]、百合[13]等植物糖的代谢具有调控作用。与其它植物类似,外源GA3可调控龙眼花芽分化[7-8]、促进花粉萌发[9],诱导龙眼焦核[10]但其对龙眼假种皮糖分积累的影响尚未明晰。课题组前期以不同浓度的GA3处理龙眼花穗,发现10 mg/L GA3显著提高了龙眼可溶性糖水平。本研究以‘石硖’龙眼为试材,采用10 mg/L GA3处理果实,测定GA3处理对龙眼果实生长发育、假种皮糖分积累以及蔗糖代谢相关酶活性的影响,为探讨龙眼果实糖积累机理及激素调控龙眼果实糖代谢提供理论依据。
1 材料与方法
1.1 材料
试验于2014年4月在广东湛江中国热带农业科学院南亚热带作物研究所进行。试材为龙眼品种“石硖”,树龄10年,株距3 m×3 m。选取生长健壮,肥水长势一致的龙眼树3棵,单株小区,重复3次。实验设2个处理,每棵树从不同方向分别选取无病虫害的花序20枝,其中10枝于花后5 d喷施10 mg/L GA3,10枝喷施清水作对照,挂牌标记。自花后一周开始采样,每周采样1次,至果实成熟结束。每棵树每个处理随机采样,采样后立即装入冰盒带回实验室,一部分测定其形态指标,一部分液氮冷冻后置于-80 ℃超低温冰箱保存,用于测定糖的含量和蔗糖代谢相关酶的活性。因花后45 d龙眼假种皮开始发育,因此假种皮从花后45 d开始剥取并保存备用。
1.2 方法
1.2.1 果实发育动态研究 每次采样从每个处理中随机选取10个果实,用电子天平测定单果重量和假种皮重量,用游标卡尺测定果实纵横径,用排水法测定果实体积。
1.2.2 糖的提取和测定 总糖的测定方法参照马志良[14]的蒽酮法。单糖的提取参照赵智中[15]的方法,糖含量采用高效液相色谱(HPLC)测定,色谱条件为:Series200 氨基酸柱,柱温35 ℃,流动相80 %乙腈,流速1 mL/min,PE高效液相色谱仪及其相应的数据处理系统。
1.2.3 酶的提取 蔗糖代谢相关酶的提取参照赵智中[15]的方法,测定参照张秀梅[16]的方法。所有的操作均在低温进行。重复3次。
2 结果与分析
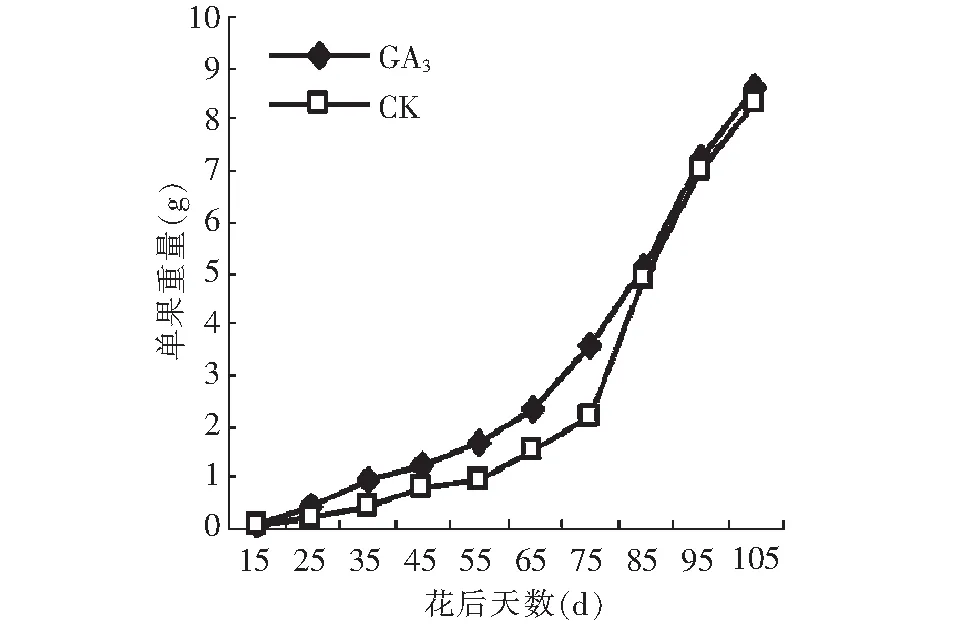
图1 GA3处理对单果重量的影响Fig.1 Effect of GA3 on weight of fruit
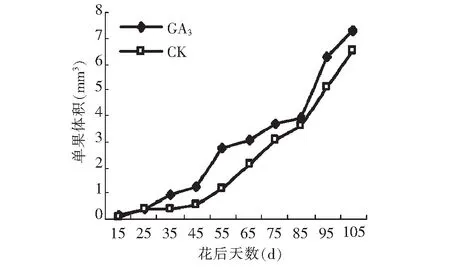
图2 GA3处理对果实体积的影响Fig.2 Effect of GA3 on volume of fruit
2.1 GA3处理对龙眼果实生长发育的影响
GA3处理后单果重量的变化趋势和对照基本一致,均呈先慢后快的上升趋势(图1)。除花后75~85 d对照的单果重变化幅度大于GA3处理之外,其余各阶段均为GA3处理后单果重变化幅度较大,说明GA3处理对单果重的增加有明显的促进作用。在成熟果中,GA3处理后单果重仅稍高于对照,二者差异不显著。
GA3处理后龙眼果实体积的膨大呈慢-快-慢-快逐渐增加的变化趋势,这与对照(慢-快)的变化趋势有较大差异。由图2可以看出,除花后15~25 d外,GA3处理对果实体积膨大有明显的促进作用,尤其是在45~55,85~95 d体积增加的幅度较大,说明这2个阶段,GA3处理的促进作用更为明显。
由图3~4可见,GA3处理对果实纵横径有显著的促进作用,其由快到慢逐渐增大的变化趋势和对照基本一致,且同步变化。数据分析显示,15~25,55~75 d,以及95~105 d GA3处理后纵径生长量大于对照(图5),在15~35,65~75 d,以及85~105 d GA3处理后横径生长量大于对照(图6),而15~35,55~75 d以及85~105 d这3个阶段分别处于果实发育初期,假种皮缓慢生长期和假种皮快速生长期,说明GA3处理对这3个发育时期果实纵横径的促进作用较大。果实成熟期,GA3处理后纵横径相比对照分别增加了0.57和1.89 mm。在花后50 d前,石硖龙眼果实纵径生长的速度大于横径,在花后50 d后,横径略大于纵径,GA3处理和对照都遵循这一规律。
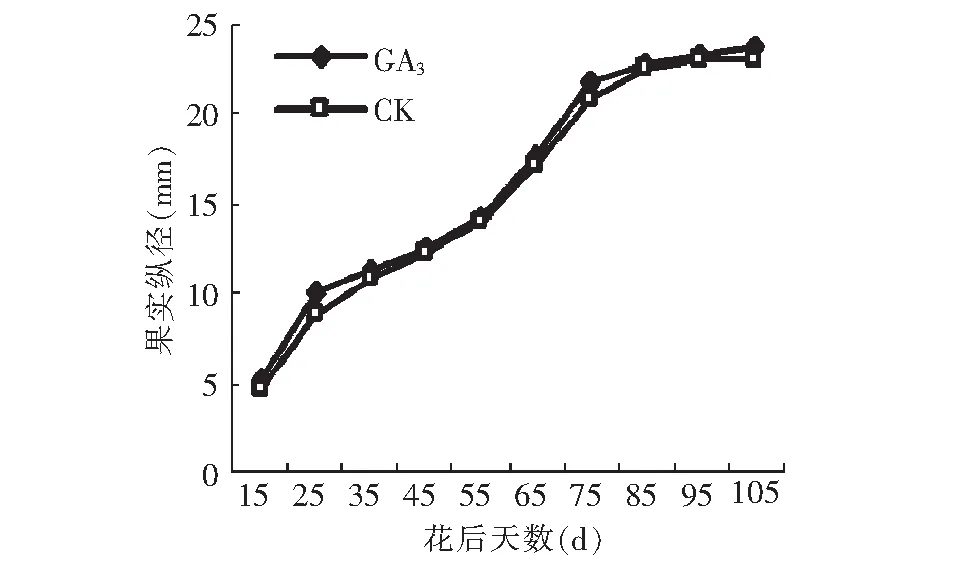
图3 GA3处理对果实纵径的影响Fig.3 Effect of GA3 on diameter of fruit
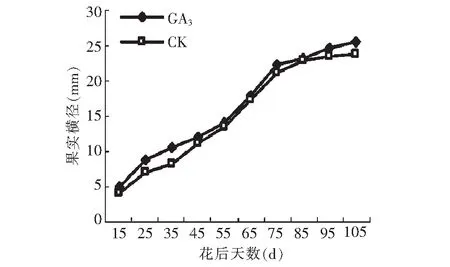
图4 GA3处理对果实横径的影响 Fig.4 Effect of GA3 on transverse diameter of fruit
由图7可以看出,GA3处理对假种皮重量的影响非常明显,GA3处理没有改变假种皮重量的变化趋势,二者均表现出由慢到快的上升趋势。在花后45~55, 65~75 d GA3处理的假种皮重量变化幅度高于对照,说明GA3处理在这2个时间段对假种皮重量的促进作用较为明显。到果实成熟期,GA3处理后的假种皮重量比对照高0.358 g。
2.2 GA3处理对龙眼果实生长发育过程中糖组分的影响
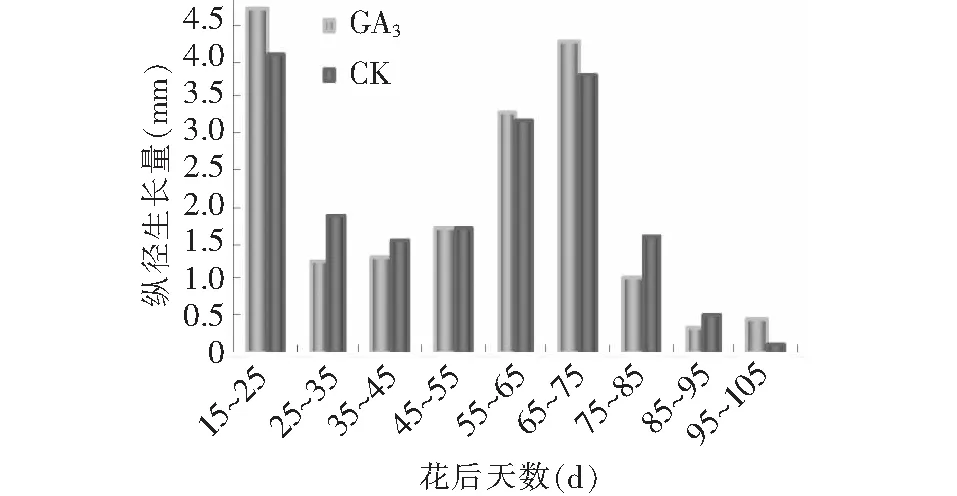
图5 GA3处理对果实纵径生长量的影响Fig.5 Effect of GA3 on growth of diameter of fruit
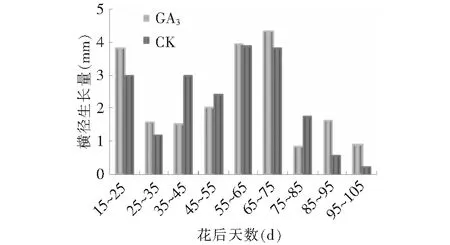
图6 GA3处理对果实横径生长量的影响Fig.6 Effect of GA3 on growth of transverse diameter of fruit
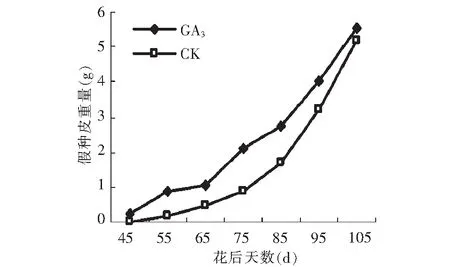
图7 GA3处理对假种皮重量的影响Fig.7 Effect of GA3 on weight of sarcocarp
由图8可以看出,在假种皮生长发育过程中,GA3处理后可溶性总糖的含量基本呈慢-快-慢的增长趋势,在花后45~65 d总糖含量缓慢上升,之后呈迅速上升趋势,到花后85 d,总糖积累趋于缓和。而对照总糖含量以65 d为节点呈先慢后快的积累趋势。花后60 d之后,GA3处理后总糖含量高于对照,到成熟期,二者总糖积累量相当,说明GA3处理可以提高总糖积累效率,但不能改变其积累量。
图9显示,GA3处理对果糖积累的促进效果明显,其变化趋势跟对照均呈“慢-快-慢”的曲线变化。但GA3处理后果糖的积累速度和积累量明显高于对照,成熟期二者果糖的含量基本一致。
GA3处理后葡萄糖的积累和对照均呈先慢后快逐渐增加的变化趋势(图10)。花后45~95 d,GA3处理后葡萄糖的积累量和积累速度均高于对照,因此,GA3对葡萄糖的促进作用主要体现在果实成熟前。成熟期葡萄糖的含量跟对照差异不显著,因此GA3处理只提高葡萄糖积累速率,并不能改变葡萄糖积累的变化规律和成熟期葡萄糖的含量。
GA3处理后蔗糖的积累变化趋势跟对照均呈先升高后降低的变化趋势,从45 d到花后80 d,蔗糖含量呈上升趋势,从花后80 d以后,蔗糖含量迅速降低。到成熟期,蔗糖含量与果糖和葡萄糖总含量相当,蔗糖成为假种皮中的主要糖分。假种皮缓慢生长期(45~55 d)和假种皮快速生长期(80~10 5 d)的蔗糖含量均低于对照,而中间时期(60~80 d)的蔗糖含量高于对照(图11)。
2.3 GA3处理对龙眼果实发育过程中的糖代谢相关酶的影响
由图12可以看出, GA3处理显著降低了AI活性,并改变其变化趋势。GA3处理后AI活性呈慢-快-慢的下降趋势,在花后45~55 d,AI活性迅速降低,之后降低速率减缓,到花后95 d,又开始急剧下降至最小值4.29 μmol/h·g·FW。而对照呈先升高后下降的趋势,果实成熟期达到最低峰值,此时的AI活性是GA3处理的3倍。
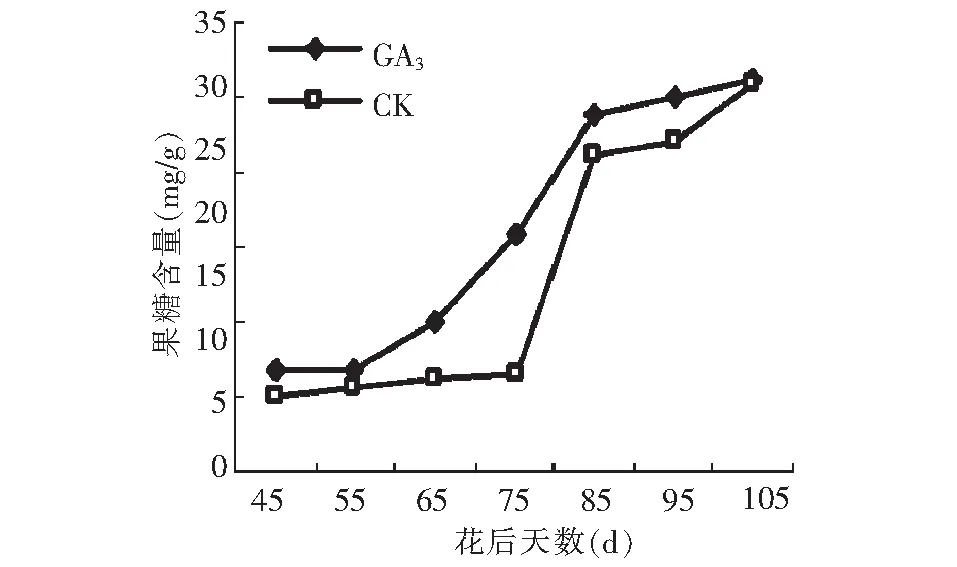
图9 GA3处理对假种皮果糖含量的影响Fig.9 Effect of GA3 on fructose of fruit
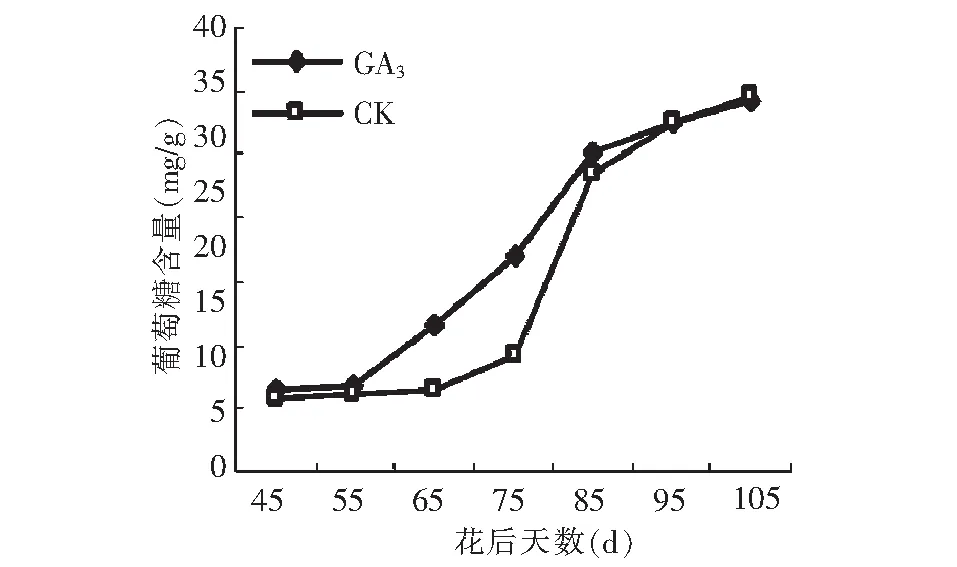
图10 GA3处理对假种皮葡萄糖含量的影响Fig.10 Effect of GA3 on glucose of sarcocarp
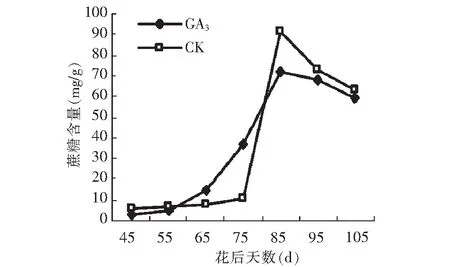
图11 GA3处理对假种皮蔗糖含量的影响Fig.11 Effect of GA3 on sucrose of sarcocarp
NI活性在假种皮发育过程中呈波浪状动态变化。假种皮缓慢生长期,GA3处理和对照NI活性均为先升高后降低的趋势,假种皮快速生长期,GA3处理后NI活性以升高-降低-升高的趋势变化,而对照则为升高-降低的变化趋势。到果实成熟时,GA3处理后NI活性达到最高峰值6.89 μmol/h·g·FW,比对照高0.80 μmol/h·g·FW。
由图14可以看出,GA3处理与对照SPS活性均呈升高~降低~升高的变化趋势,但GA3处理后SPS活性最高峰值和最低峰值出现的时间较对照延迟。GA3处理后SPS活性最大值比对照小2.0 μmol/h·g·FW,峰值出现的时间比对照延迟10 d;GA3处理后SPS活性的最低峰值跟对照差异不显著,但比对照的最低值出现的时间晚20 d。到果实成熟期,二者的活性差异不大。除花后60~80 d,GA3处理后SPS活性高于对照外,其他阶段均低于对照。
对SS活性测定结果表明:在SS合成蔗糖方向,GA3处理显著提高了SS活性,且改变了SS活性变化趋势。GA3处理后SS活性呈升高-下降-升高的趋势,与对照(升高~降低)有显著差异。GA3处理后SS活性在花后65 d出现最高峰值8.53 μmol/h·g·FW,较对照最高峰值出现的时间提前30 d,且最高值较对照高1.15 μmol/h·g·FW。SS分解蔗糖方向的活性变化较为复杂。GA3处理的SS活性在花后45 d处于较低水平,而后迅速升高至峰值16.26 μmol/h·g·FW,随后又开始降低,到果实成熟期又开始升高。而对照在花后45~55 d出现小幅度下降,然后迅速升高至峰值16.85 μmol/h·g·FW,而后又迅速降低,到果实成熟期又开始上升,成熟期两者SS活性差异不显著。GA3处理使峰值出现的时间延迟。
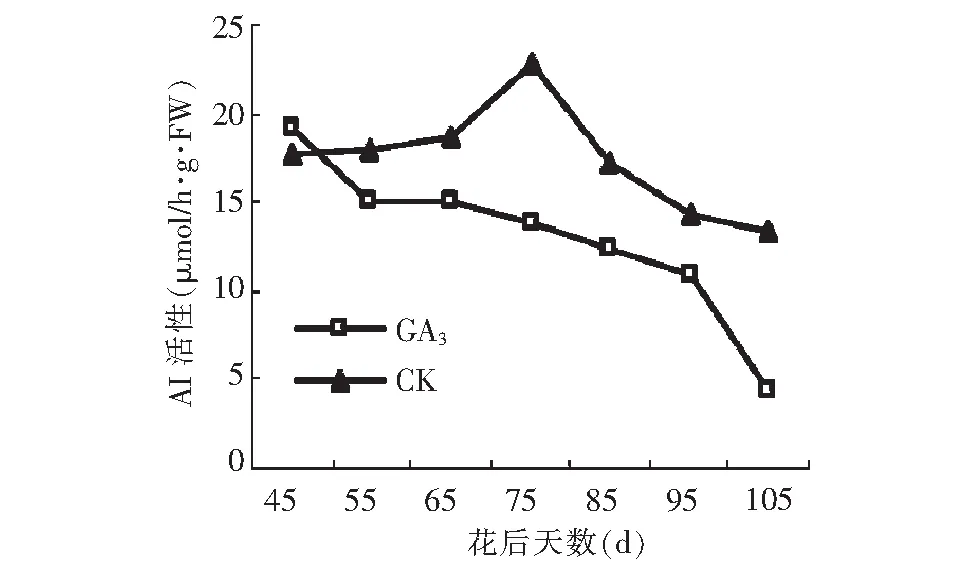
图12 GA3处理对假种皮酸性转化酶活性的影响Fig.12 Effect of GA3 on activities of AI of sarcocarp
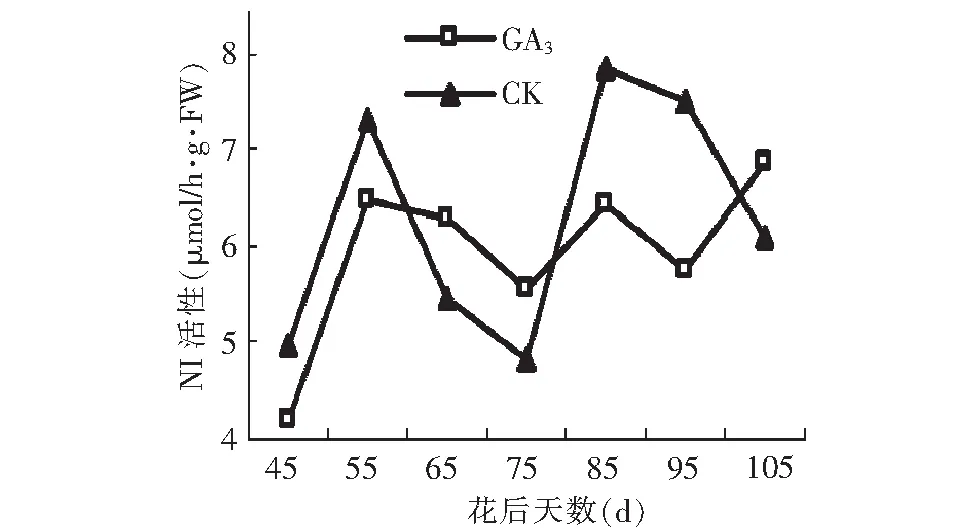
图13 GA3处理对假种皮中性转化酶活性的影响Fig.13 Effect of GA3 on activities of NI of sarcocarp
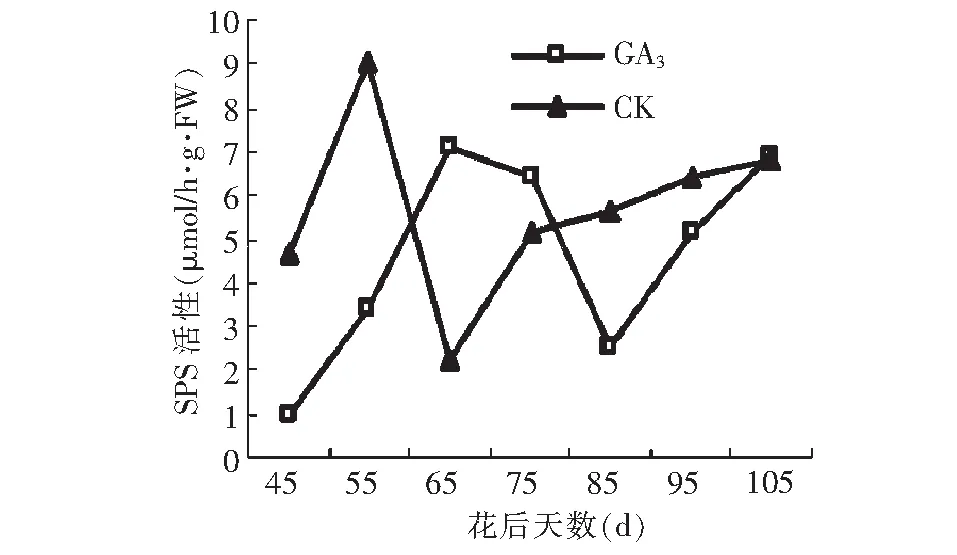
图14 GA3处理对假种皮磷酸蔗糖合成酶活性的影响Fig.14 Effect of GA3 on activities of SPS of sarcocarp
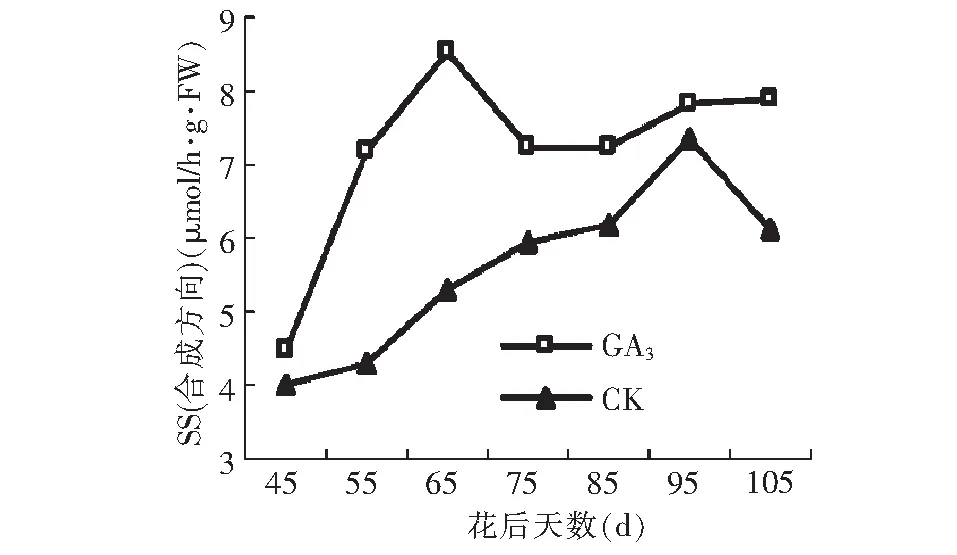
图15 GA3处理对假种皮蔗糖合成酶(合成方向)活性的影响Fig.15 Effect of GA3 on activities of SS(the composition direction)of sarcocarp
2.4 GA3处理后糖分积累与糖代谢相关酶的关系
GA3处理可以促进糖的积累,然而糖的积累和代谢离不开蔗糖代谢相关酶的调控和影响。在“石硖”龙眼假种皮生长发育期,GA3处理和对照的AI 、SS(分解方向)的酶活性均维持较高水平,大于其他酶类,因此,推测石硖龙眼糖代谢起主导作用的酶为AI和SS(分解方向)。假种皮缓慢生长期,GA3处理显著降低了 AI活性,然而其活性大小仍处于较高水平,故此期果糖和葡萄糖积累迅速,假种皮快速生长期,AI活性的降至较低水平,此时蔗糖迅速积累,到成熟期,蔗糖含量远高于果糖和葡萄糖,成为果实中的主要糖分。在假种皮缓慢生长期, GA3处理后 SS(分解方向)的活性从较低水平迅速升高至最高峰值,高活性的SS(分解方向)促进果糖和葡萄糖积累;从花后75 d逐渐减小,低活性的SS(分解方向)有利于蔗糖积累。NI、SPS、SS(合成方向)的活性大小均在8 μmol/h·g·FW以下上下波动,低活性的NI,高活性的SPS和SS(合成方向)均有利于蔗糖积累。蔗糖的积累是由蔗糖代谢相关酶的综合作用决定的。
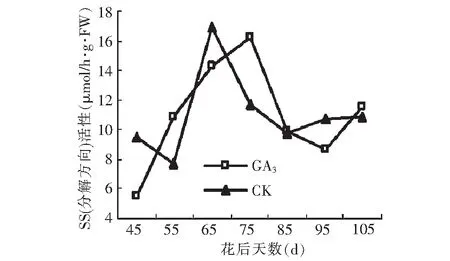
图16 GA3处理对假种皮蔗糖合成酶(分解方向)活性Fig.16 Effect of GA3 on activities of SS(the decomposition direction)of sarcocarp
3 讨 论
研究结果表明,GA3处理可以提高生长发育过程中单果重量,单果体积,果实纵横径和假种皮重量,但是没有改变其变化规律,这可能与GA3可刺激细胞壁松弛酶基因和细胞分裂基因的表达,也刺激细胞扩展过程中微管的重排,加快果肉细胞分裂,促进细胞伸长增大的作用有关。依据蔡可[17]的观点,外源GA3在植物体内会逐渐被降解、钝化,有效期为2周左右,故作者推测,外施GA3在植物生长发育过程中是并不都是直接发挥作用,它可以增加生长素的含量,通过影响生长素的运输和信号传导来促进细胞的扩展并吸引营养物质向处理部位运转,使果实成为强“库”,在营养竞争中处于有利地位,从而间接起到促进生长的作用,这与李鹏程[3]在葡萄上的结果吻合。
本研究表明,GA3处理使蔗糖积累的转折点从花后75 d提前到花后55 d,且加快了糖的积累效率,但没有显著改变成熟果实中糖的积累量,说明GA3并不能改变龙眼果实中糖的积累类型。前人的研究结果显示,GA3在植物体内有一定的时效性[18],说明GA3的作用只能维持一定的时间。本研究也发现,GA3对糖积累的促进作用体现在一定阶段,如,总糖在花后60 d之后,果糖在果实整个生长发育过程,葡糖糖在果实成熟前,而蔗糖在花后60~80 d,但快速积累的时间均在花后55~85 d,故作者推测GA3处理在花后55~85 d的促进作用最大,而后期糖含量增加是这种优势一直保持的结果。
果实糖分积累受蔗糖代谢相关酶的调控和影响,本研究中,GA3处理改变了蔗糖代谢相关酶活性的大小和变化趋势,显著降低了AI活性,提高了SS(合成方向)活性,而对SPS、NI、SS(分解方向)的活性和变化趋势的影响较为复杂。依据吕英民[19]的观点,GA3对蔗糖代谢相关酶的影响可能是在基因表达的不同水平上(转录翻译和翻译后),调节与代谢相关的关键酶和运输糖的载体作用的;分子生物学的研究证实,果实的糖积累与蔗糖代谢酶的基因表达有关[4];Rolland等也认为,糖响应与植物激素合成或信号转导突变体之间存在复杂的内在联系,很可能是内在的糖信号与外在信号共同作用调控了果实的发育与糖积累[3]。因此,作者推测GA3对糖分积累的影响是通过调节和改变蔗糖代谢相关酶的活性而间接作用的。
4 结 论
GA3处理可以提高“石硖”龙眼生长发育过程中单果重量,单果体积,果实纵横径,假种皮重量,但是没有改变其变化规律;GA3处理加快假种皮发育过程中糖的积累效率,但并不能改变糖的积累类型;GA3对糖分积累的影响是通过调节和改变蔗糖代谢相关酶的活性而间接作用的。研究糖与植物激素信号之间的联系,是揭示果实糖代谢和积累机制的重点,也是今后研究的方向。
[1]吴伟民,钱亚明,赵密珍,等. 赤霉素对魏可葡萄无核果实的诱导效果及对品质的影响[J]. 中外葡萄与葡萄酒,2010(3):38-39.
[2]杜晓彧,常海飞,冯晓东. 不同浓度赤霉素对金丝小枣叶片生长和果实品质的影响[J]. 延安大学学报(自然科学版),2014,33(1):50-53.
[3]李鹏程,郁松林,符小发,等. GA3对葡萄果实糖积累及蔗糖代谢酶的影响[J]. 西北农林科技大学学报(自然科学版),2011,10:177-183.
[4]陈俊伟,冯健君,秦巧平,等. GA3诱导的单性结实‘宁海白’白沙枇杷糖代谢的研究[J]. 园艺学报,2006,33(3):471-476.
[5]姜仕豪,庞基良,王利琳,等. 赤霉素促进花发育的分子机制[J]. 植物生理学通讯,2008,44(5):835-843.
[6]周 玲,魏小春,郑 群,等. 脱落酸与赤霉素对瓜尔豆叶片光合作用及内源激素的影响[J]. 作物杂志,2010(1):15-20.
[7]苏明华,刘志成,刘惠婉. 赤霉素对水涨龙眼花芽分化的效应[J]. 亚热带植物通讯,1990(2):35-39.
[8]彭 坚,李永红,席嘉宾,等. 化学调控四季花龙眼成花座果的初步研究[J]. 西北植物学报,2005,25(7):1440-1445.
[9]赵春香,彭乃容,李新福,等. 龙眼花粉生活力的研究[J]. 福建果树,2002(4):8-10.
[10]李 明. 诱导龙眼焦核果实的研究[D].福建农林大学,2008.
[11]李节法,杨 琪,虞秀明,等. 赤霉素对梨糖代谢及其关键酶基因表达的影响[J]. 上海交通大学学报(农业科学版),2015(3):21-28+35.
[12]孙盼盼,李建贵,王 娜. 叶面肥和赤霉素对骏枣果实生长发育中糖积累的影响[J]. 西北农业学报,2011,20(12):98-102.
[13]孙红梅,何 玲,王微微,等. IBA与GA3调控百合鳞片扦插繁殖的“淀粉-蔗糖”代谢机制[J]. 中国农业科学,2011,44(4):798-806.
[14]张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社, 2003.
[15]赵智中,张上隆,徐昌杰,等.蔗糖代谢相关酶在温州蜜柑果实糖积累中的作用[J].园艺学报,2001,28(2):112-118.
[16]张秀梅,杜丽清,孙光明,等.菠萝果实发育过程中糖积累与其代谢酶的关系[J].热带作物学报,2008,29(1):10-13.
[17]蔡 可. 植物激素和生长调节物质的物理、化学和生物学特性[J]. 植物生理学通讯,1984(6):49-55.
[18]吴建明,李杨瑞,杨 柳,等. 赤霉素诱导甘蔗节间伸长与内源激素变化的关系[J]. 热带作物学报,2009, 30(10):1452-1457.
[19]吕英民,张大鹏. 果实发育过程中糖的积累[J]. 植物生理学通讯,2000,36(3):258-265.
(责任编辑 李 洁)
Effects of GA3on Sugar Accumulation and Sugar Metabolism-related Enzyme Activities in Shixia ofDimocarpuslonganLour Fruits
LIU Li-qin1, LI Jian-guang2, SHU Bo1,WANG Yi-cheng1, WU Hong-xia1, JUE Deng-wei1, SHI Sheng-you1*
(1.South Subtropical Crops Research Institute, Chinese Academy of Tropical Agricultural Science, Key Laboratory Tropical Fruit's Biology, Ministry of Agriculture, Guangdong Zhanjiang 524091, China; 2.Institute of Fruit Tree Research,Guangdong Academy of Agricultural Sciences, Guangdong Guangzhou 510640,China)
To understand the effects of gibberellin on the Longan fruit development and sugar production, the index involving fruit development, the aril sugar content and the enzyme activities were measured in both GA3treated ‘Shixia’ of Lonagn and the control. The results showed that GA3treatment increased the weight, volume, longitudinal diameter and traverse diameter of fruit and the weight of aril. Sucrose is main sugar of ‘Shixia’ longan in aril , and sucrose metabolism was mediated mainly by AI and SS (the decomposition direction). GA3treatment did not affect sugar accumulation in aril but enhanced the efficiency of sucrose accumulation by activating the sucrose metabolism-related enzymes .
GA3;Fruit development;Sugar accumulation; Sucrose metabolism-related enzyme
1001-4829(2016)11-2669-06
10.16213/j.cnki.scjas.2016.11.034
2015-12-28
国家自然科学基金项目(31572087);广东省自然科学基金(2014A030307031);中央级公益性科研院所基本科研业务费专项(1630062015004);物种资源保护(农作物)项目(2016NWB053);物种资源保护费(农业野生植物保护)项目(1251416305010);国家热带果树种质资源平台运行服务项目(095)
刘丽琴,(1979-)女,助理研究员,硕士,主要从事热带果树遗传育种研究,Tel:0759-2859112, E-mail:lolitallq@163.com, *为通讯作者,Tel:0759-2859112, E-mail:ssy7299@163.com。
S667.2
A
