宁夏贺兰山种子植物区系垂直分布格局及其气候解释
2016-12-14张东杰曾小强张化永
张东杰,徐 翔,曾小强,张化永
(华北电力大学工程生态学与非线性科学研究中心,北京 102206)
宁夏贺兰山种子植物区系垂直分布格局及其气候解释
张东杰,徐 翔,曾小强,张化永*
(华北电力大学工程生态学与非线性科学研究中心,北京 102206)
根据我国种子植物区系地理划分,结合当地考察资料,探讨了宁夏贺兰山自然保护区种子植物及不同分布区类型的垂直分布格局,并利用SPSS软件对物种丰富度和气候因子进行了回归分析。结果表明,宁夏贺兰山种子植物以温带区系成分为主,约占73.9%,植物物种丰富度沿海拔梯度呈单峰分布,在2 000 m左右达到峰值,在高海拔地区(2 600 m以上)物种丰富度变化不明显。回归分析结果表明,种子植物和温带区系成分垂直分布格局主要受年均温和年潜在蒸散量影响,世界分布垂直分布格局受年均温、年潜在蒸散量、年降水量和最冷月均温影响,热带区系成分垂直分布格局受年均温、年潜在蒸散量、年降水量、最冷月均温和最冷季降水量影响。
宁夏贺兰山自然保护区;种子植物;区系成分;气候因子;垂直分布格局
物种多样性的空间分布格局及其形成机制是宏观生态学和生物地理学研究的核心问题之一,也是保护生物学研究的基础[1]。由于海拔梯度包含了多种环境因子(温度、降雨、光照等)的梯度效应,因此它在一定程度上可以显著影响植物种群分布格局[2]。许多研究表明,物种沿海拔梯度的变化趋势主要有4种,即随海拔梯度先升高后降低(单峰分布格局)、先降低后升高、随海拔升高而降低、随海拔升高而递增和无规律性,但其具体成因尚未达成一致的观点[3-5]。近年来,随着生态学的发展,许多学者试图确定造成多样性分布差异的决定因素,并因此提出了许多假说和理论[6-12]。例如,岛屿假说认为,物种多样性与其分布面积之间存在显著关系[13]。Currie等[14]通过对北美洲树木多样性的研究认为,实际蒸散量是影响其地理格局的主导因子。水分-能量动态假说认为物种多样性的大尺度格局主要受到水分和能量的影响,水分可以调节生物体对能量的利用过程[6]。Wang等[15]研究了多个气候因素对我国东北地区森林分布格局的影响,结果显示生长季降水和年潜在蒸散量在解释森林分布时占主导地位。冯建孟[16]在对我国种子植物地理分布格局的研究中认为,在大尺度的物种多样性格局的研究中,气候因子的空间变异和季节性分异可能比反映总体水平的气候因子更值得关注。尽管已经做了很多研究,但迄今为止,还没有一种理论或假说可以完全解释所有地区的物种多样性分布格局,因此,物种多样性的空间分布格局及其影响因素有待进一步深入研究。
植物分布区理论认为物种分布区的形成是物种形成过程的空间反映,它不仅是物种迁移、系统分化等历史过程的积累,也是生态环境及其异质性格局影响的结果。参照吴征镒等[17]关于我国种子植物属的分布区类型的界定,种子植物可以划分为15个植物分布区类型,分别为世界分布(1),泛热带区系成分(2),热带亚洲和热带美洲间断分布(3),旧世界热带区系成分(4),热带亚洲至热带大洋洲分布(5),热带亚洲至热带非洲分布(6),热带亚洲分布(7),北温带区系成分(8),东亚和北美洲间断分布(9),旧世界温带区系成分(10),温带亚洲分布(11),地中海区,西亚至中亚分布(12),中亚分布(13),东亚分布(14),中国特有分布(15)。其中,2~7统称为热带区系成分,8~15为温带区系成分。植物区系研究在理论上可以揭示和解决植物学的一些疑难问题,同时也为保护和持续利用植物资源提供重要的参考资料。
目前,利用动植物志等资料研究生物多样性分布格局是一种较为成熟的方法[16,18]。笔者根据宁夏贺兰山自然保护区科学考察文献中的种子植物名录资料,结合我国种子植物区系划分,分别探讨了种子植物、世界分布、热带区系成分和温带区系成分物种丰富度的垂直分布格局,并利用年均温、年降水量、年潜在蒸散量等水热气候因子对其进行解释。该研究期望为当地植物物种保护提供理论基础,同时对研究相似气候条件下植物物种多样性空间分布格局提供参考。
1 研究对象与方法
1.1 研究区域概况 宁夏贺兰山自然保护区位于宁夏西北部,地处我国温带草原与荒漠两大植被区域的过渡带,属中温带干旱气候区,具有典型的大陆季风气候特点。保护区总面积1 935.35 km2,海拔1 400~3 500 m,最高峰俄博疙瘩3 556.1 m。由于山体陡峭、地形复杂,保护区内山地气候明显。气象站常年数据显示,贺兰山年平均气温为-0.7 ℃,最冷月均温为-13.9 ℃,最热月均温为12.1 ℃,气温年较差达26.0 ℃。年平均降水量400 mm左右,降水的季节变化大,夏季降水量最多可达全年降水量的60%以上。受到气候、土壤等因素影响,贺兰山保护区植被具有明显的垂直分异,按植被类型可划分成4个植被垂直带:山前荒漠与荒漠草原带;山麓与低山草原、灌丛带;中山针叶林带;高山、亚高山灌丛、草甸带[19]。保护区内主要森林群系为青海云杉林、油松林,另有杜松、山杨、白桦等零星分布。
1.2 数据来源与处理 植物数据和海拔分布信息主要来源于《宁夏贺兰山国家级自然保护区综合科学考察》植物名录记载[19],其中包含海拔信息的种子植物共56科196属402种。根据吴征镒等[17]对我国种子植物区系地理成分的划分,将宁夏贺兰山自然保护区种子植物分为世界分布、热带区系成分和温带区系成分,其中世界分布共20科25属66种,热带区系成分共13科19属39种,温带区系成分共45科152属297种。
将贺兰山按海拔间隔100 m分为不重合的22段,分别为<1 400 m、1 400~1 500 m、 …… 、3 300~3 400 m、≥3 400 m,并根据种子植物海拔分布信息统计不同海拔段的植物物种丰富度。对于只记录了中间海拔的植物,定义这些物种只分布在该海拔段。由此得到宁夏贺兰山种子植物、世界分布、热带区系成分和温带区系成分在各个海拔段上的物种丰富度数据。根据美国航空航天局(NASA)和日本经济产业省(METI)联合发布的全球30 m×30 m分辨率海拔数据(ASTER GDEM V2)和各个高度带的海拔范围,使用ArcGIS裁剪出贺兰山各个海拔段的位置矢量图,利用WorldClim数据库所提供的全球30″分辨率气候数据[20]提取各个海拔段的气候数据。
研究所选用的气候因子包括年均温和年降水量,其中年均温反映热量因子对物种多样性格局的影响,年降水量则反映水分因子的影响。同时,根据Thornthwaite潜在蒸散量计算方法,计算年潜在蒸散量并作为水热综合因子进行研究。根据寒冷忍耐假说[8],物种多样性受到“恶劣”环境的影响严重,环境的“恶劣”主要指一些极端低温,因此笔者选取了最冷月均温作为极端条件下的热量因子进行研究,选取最冷季降水量作为极端条件下的水分因子(表1)。
1.3 研究方法 研究通过描述植物物种丰富度和气候因子之间的关系来解释物种丰富度沿海拔梯度的变化规律,使用SPSS对数据进行统计分析,对选取的气候因子和不同分布区类型植物物种丰富度进行初步分析;再通过逐步回归分析建立物种丰富度和气候因子之间的多元线性回归模型,并对结果进行讨论。

表1 影响植物物种多样性的气候因子
2 结果
2.1 种子植物物种丰富度沿海拔梯度的变化规律 宁夏贺兰山种子植物及不同分布区类型植物物种丰富度随海拔变化如图1所示。
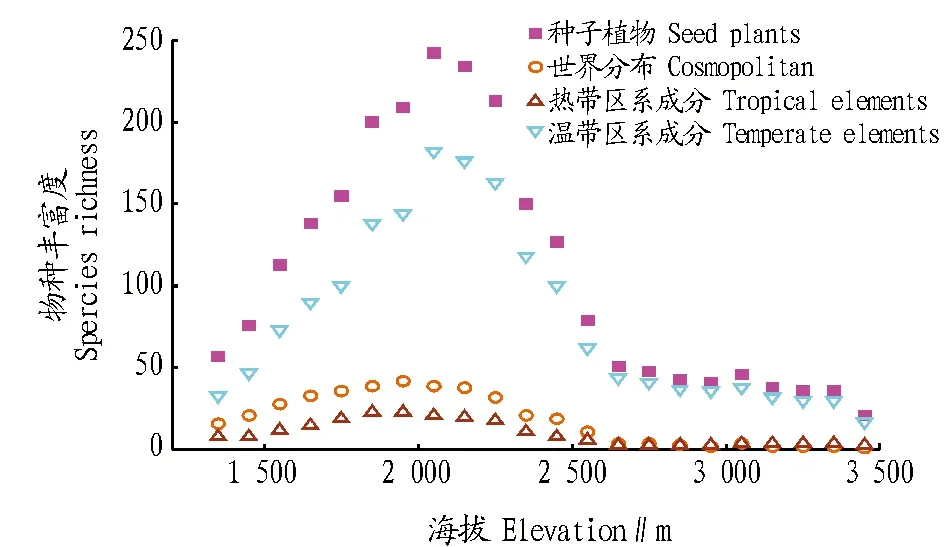
图1 种子植物及不同区系成分物种丰富度的垂直分布格局Fig.1 Altitudinal patterns of species richness for seed plants and different floristic composition
如图1所示,贺兰山种子植物、世界分布、热带区系成分和温带区系成分物种丰富度沿海拔梯度基本呈单峰分布(先升高后降低),峰值出现在2 000 m左右,在高海拔地区(2 600 m以上)物种丰富度随海拔升高变化不明显。在各个海拔段上,温带区系成分均占主导地位,在种子植物中的比例基本超过60%,且所占比例随着海拔升高逐渐增加,到海拔2 600 m及以上地区,温带区系成分占种子植物比例可达85%以上。总体而言,植物物种丰富度大小关系为温带区系成分>世界分布>热带区系成分。
2.2 气候因子对种子植物空间分布格局的影响 对植物物种丰富度和气候因子做相关性分析。植物物种丰富度和气候因子均存在一定相关性,和热量因子(AMT、MTCM)、水热综合因子(PET)呈正相关,和水分因子(AP、PCQ)呈负相关(表2)。
表2 植物物种丰富度和气候因子之间的相关性分析
Table 2 Correlation analysis of plant species richness and climatic factors
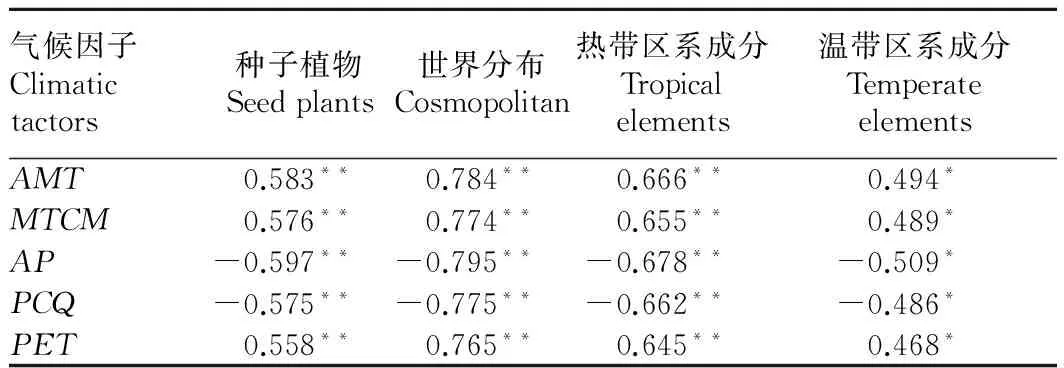
气候因子Climatictactors种子植物Seedplants世界分布Cosmopolitan热带区系成分Tropicalelements温带区系成分TemperateelementsAMT0.583**0.784**0.666**0.494*MTCM0.576**0.774**0.655**0.489*AP-0.597**-0.795**-0.678**-0.509*PCQ-0.575**-0.775**-0.662**-0.486*PET0.558**0.765**0.645**0.468*
注:**在0.01水平(双侧)上显著相关;*在 0.05水平(双侧)上显著相关。
Note:** stands for significant correlation at 0.01 level;* stands for significant correlation at 0.05 level.
再以气候因子作为解释变量,以不同分布区类型植物物种丰富度数据作为因变量,利用逐步回归的方法,筛选出满足统计检验的自变量,并以此为基础建立种子植物、世界分布、热带区系成分和温带区系成分物种丰富度垂直分布格局的回归模型。回归模型及相关检验统计量如表3所示。
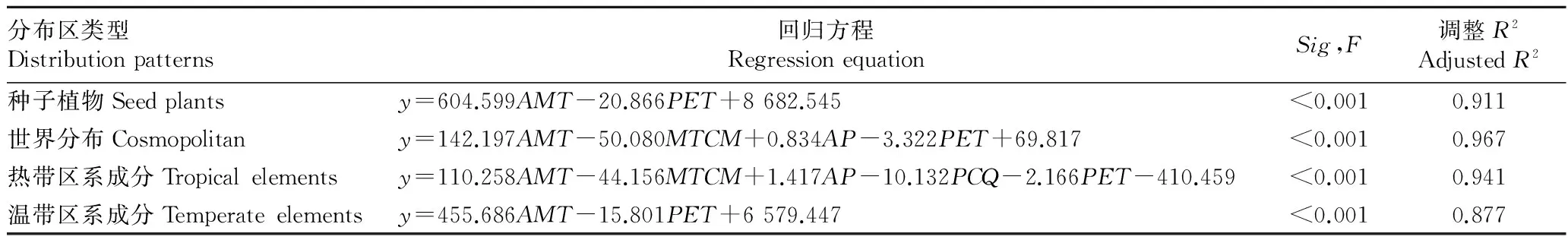
表3 基于逐步回归的物种丰富度回归模型
回归分析结果表明:影响种子植物垂直分布格局的气候因子为年均温和年潜在蒸散量,两者可以解释种子植物物种丰富度沿海拔梯度91.1%的变化。影响世界分布垂直分布格局的气候因子为年均温、最冷月均温、年降水量和年潜在蒸散量,它们的总解释率为96.7%。影响热带区系成分垂直分布格局的气候因子为年均温、最冷月均温、年降水量、年潜在蒸散量和最冷季降水量,它们的总解释率为94.1%。影响温带区系成分垂直分布格局的气候因子和种子植物相同,年均温和年潜在蒸散量可以解释物种分布格局变化的87.7%。
在逐步回归过程中,首先将自变量按相关性大小逐个引入方程,再将效应显著的自变量留在回归方程内,循此继续遴选下一个自变量。如果效应不显著,停止引入新自变量。由于新自变量的引入,原已引入方程中的自变量由于变量之间的相互作用,其效应有可能变得不显著者,经统计检验确证后要随时从方程中剔除,只保留效应显著的自变量,直至不再引入和剔除自变量为止,从而得到最优的回归方程。
该研究在逐步回归过程中,进入回归方程的解释变量顺序如下:首先引入方程的气候因子为和植物物种丰富度相关性最大的年降水量,随后在计算过程中引入年均温,又引入年潜在蒸散量之后,年降水量受变量之间相互作用影响被剔除,这时可以得到种子植物和温带区系成分植物关于物种丰富度和气候因子之间的回归模型,接着再引入热量因子最冷月均温。在新变量的影响下,年降水量再次进入模型,并得到世界分布植物物种丰富度沿海拔梯度的气候模型,再引入最冷季降水量,得到热带区系成分植物的回归模型。
3 讨论
植物区系是在一定地理环境和自然历史等综合条件下长期发展和演化的结果。在以往的研究中,将种子植物按区系进行划分并探究其分布格局往往只研究了物种丰富度沿海拔的分布格局,并没有深层次地探讨形成这种格局的原因[19,21]。该研究依据吴征镒等[7]对我国种子植物属的区系划分,将种子植物分为世界分布、热带区系成分和温带区系成分,分析了贺兰山种子植物垂直分布格局,并结合现代气候假说及相关研究[6-9],定量研究了一些重要气候因素对贺兰山种子植物和不同区系成分物种丰富度的影响,并得出相关结论。
迄今为止,有关山地植物物种多样性沿海拔梯度的变化模式尚无统一认识,但单峰曲线是一种普遍格局[3-5]。从研究结果来看,贺兰山种子植物及不同分布区类型物种多样性变化格局基本符合这一规律,物种丰富度峰值出现在海拔2 000 m左右,这和之前的研究一致[22],但在高海拔地区(海拔2 600 m以上),物种丰富度变化趋于平缓。由此可见,代表诸多环境因子综合作用的海拔梯度对种子植物多样性格局的解释偏弱(Pearson相关系数|r|<0.6),除此之外的其他因素对物种多样性垂直格局的形成可能发挥着重要作用。宁夏贺兰山为中温带干旱地区,热量和水分条件都受到区域限制,随着海拔升高总体呈现气温降低而降水增多的气候格局,在这样的气候条件下形成单峰格局的生态学解释如下。在温度因子不占主导地位的中低海拔段,增温会增强蒸散量和突然干旱程度,从而对植物生长起到限制作用;随着海拔升高到中高海拔段,降水增加,水分不再是植物生长的限制因素,物种丰富度逐渐增加[23]。对于较高海拔区,尽管降水总量大于低海拔区,但由于温度降低,潜在蒸散量下降,大大限制了植物的生长,植物多样性也因此下降。贺兰山高海拔地区(海拔2 600 m以上)年均温在0 ℃以下,植物体内可用的水分被冻结,根系从土壤中吸收水分的能力也大大减小,很多植物都无法生存在这样的环境中,尤其是对水热条件要求较高的热带区系成分,因而植物物种丰富度也呈一种较少的状态。
回归分析结果表明,年均温和年潜在蒸散量是影响种子植物和温带区系成分垂直分布格局的主要气候因子,说明种子植物和温带区系成分垂直分布格局受到热量因子影响。当热量充足时,沿海拔梯度种子植物和温带区系成分,物种多样性将会增加,而在水热综合因子的影响下,这种增长将受到限制。年均温、年潜在蒸散量,还有最冷月均温和年降水量是影响世界分布垂直分布格局的气候因子,说明水分和热量充足会增加世界分布在海拔梯度上的物种多样性,但这种增长受到水热综合因素和极端条件下热量因素的限制。年均温、最冷月均温、年降水量、年潜在蒸散量和最冷季降水量是影响热带区系成分垂直分布格局的气候因子,说明热带区系成分生长不仅需要充足的水分和热量,而且“恶劣”环境下的水热条件也会限制其物种多样性。
以上研究的结果有望为贺兰山植物保护提供理论基础,如在冬季时需对贺兰山世界分布和热带区系成分保温保湿等,同时希望能对类似气候条件下的中温带干旱地区物种多样性研究提供参考。
4 结论
该研究依据吴征镒等[17]对我国种子植物属的区系划分,将贺兰山种子植物分为世界分布、热带区系成分和温带区系成分,阐述了宁夏贺兰山种子植物及不同分布型在海拔梯度上的垂直分布格局,并得到了沿海拔梯度物种丰富度和气候因子之间的回归模型。贺兰山种子植物以温带区系成分为主,占比约73.9%,世界分布和热带区系成分比例较小,分别为16.4%和9.7%,物种丰富度随海拔升高先升高后降低,并在高海拔地区基本保持不变。对物种丰富度和选取的气候因子进行回归分析,得到的回归方程分别可以解释种子植物、世界分布、热带区系成分和温带区系成分物种丰富度随海拔梯度变化的91.1%、96.7%、94.1%和87.7%。贺兰山种子植物、温带区系成分物种多样性主要受热量因子和水热综合因子的限制,而世界分布和热带区系成分植物生长不仅受水热条件限制,而且“恶劣”环境也会对其物种丰富度造成影响。
[1] GASTON K J.Global patterns in biodiversity[J].Nature,2000,405(6783):220-227.
[2] 唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
[3] OHLEMULLER R,WILSON J B.Vascular plant species richness along latitudinal and altitudinal gradients:A contribution from New Zealand temperate rainforests[J].Ecology letters,2000,3(4):262-266.
[4] MCCAIN C M,GRYTNES J A.Elevational gradients in species richness[M]//Encyclopedia life sciences.Chichester:John Wiley & Sons,2010.
[5] MARINI L,BONA E,KUNIN W E,et al.Exploring anthropogenic and natural processes shaping fern species richness along elevational gradients[J].Journal of biogeography,2011,38(1):78-88.
[6] O’BRIEN E M.Biological relativity to water-energy dynamics[J].Journal of biogeography,2006,33(11):1868-1888.
[7] BROWN J H,GILLOOLY J F,ALLEN A P,et al.Toward a metabolic theory of ecology[J].Ecology,2004,85(7):1771-1789.
[8] 王志恒,唐志尧,方精云.物种多样性地理格局的能量假说[J].生物多样性,2009,17(6):613-624.
[9] HAWKINS B A,ALBUQUERQUE F S,ARAUJO M B,et al.A global evaluation of metabolic theory as an explanation for terrestrial species richness gradients[J].Ecology,2007,88(8):1877-1888.
[10] ZHAO S,FANG J.Patterns of species richness for vascular plants in China’s nature reserves[J].Diversity and distributions,2006,12(4):364-372.
[11] HAWKINS B A,RODRIGUEZ M,WELLER S G.Global angiosperm family richness revisited:Linking ecology and evolution to climate[J].Journal of biogeography,2011,38(7):1253-1266.
[12] ROWE R J.Environmental and geometric drivers of small mammal diversity along elevational gradients in Utah[J].Ecography,2009,32(3):411-422.
[13] FRASER R H,Currie D J.The species richness-energy hypothesis in a system where historical factors are thought to prevail:Coral reefs[J].American naturalist,1996,148(1):138-159.
[14] CURRIE D J,PAQUIN V.Large-scale biogeographical patterns of species richness of trees[J].Nature,1987,329(6137):326-327.
[15] WANG X P,TANG Z Y,FANG J Y.Climatic control on forests and tree species distribution in the forest region of northeast China[J].Journal of integrative plant biology,2006,48(7):778-789.
[16] 冯建孟.中国种子植物物种多样性的大尺度分布格局及其气候解释[J].生物多样性,2008,16(5):470-476.
[17] 吴征镒,孙航,周浙昆,等.中国种子植物区系地理[J].生物多样性,2011,19(1):124-124.
[18] 郑智,龚大洁,孙呈祥,等.秦岭两栖、爬行动物物种多样性海拔分布格局及其解释[J].生物多样性,2014,22(5):596-607.
[19] 王小明.宁夏贺兰山国家级自然保护区综合科学考察[M].银川:阳光出版社,2011.
[20] HIJMANS R J,CAMERON S E,PARRA J L,et al.Very high resolution interpolated climate surfaces for global land areas[J].International journal of climatology,2005,25(15):1965-1978.
[21] 赵鸣飞,王宇航,邢开雄,等.黄土高原山地森林群落植物区系特征与地理格局[J].地理学报,2014,69(7):916-925.
[22] 朱源,江源,刘全儒,等.基于等面积高度带划分的贺兰山维管植物物种丰富度的海拔分布格局[J].生物多样性,2007,15(4):408-418.
[23] 潘红丽,李迈和,蔡小虎,等.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722-730.
Altitudinal Patterns of Seed Plants Floristic Composition and Their Climatic Explanation in Helan Mountain Nature Reserve, Ningxia Province
ZHANG Dong-jie, XU Xiang, ZENG Xiao-qiang, ZHANG Hua-yong*
(Research Center for Ecological Engineering and Nonlinear Science, North China Electric Power University, Beijing 102206)
Based on floristics of seed plants from China and documented data, we explored the patterns of seed plants richness along an altitudinal gradient in Mountain Helan Nature Reserve, Ningxia Province, China. Regression analysis of species richness and climatic factors was carried out by SPSS software. The results indicated that the main component of seed plants was temperate elements (73.9%). The species richness of seed plants and its floristic composition showed a unimodal pattern along the altitude in Mountain Helan Nature Reserve, up to peak value at about 2 000 m, species richness was not significantly changed at high altitude region (above 2 600 m). Annual mean temperature and annual potential evapotranspiration was the variable to explain the altitudinal pattern of seed plants and temperate elements species richness. In addition to these two climatic factors, cosmopolitan was influenced by min temperature of coldest month and annual precipitation, and tropical elements was influenced by min temperature of coldest month, annual precipitation and precipitation of coldest quarter.
Ningxia Helan Mountain Nature Reserve; Seed plants; Floristic composition; Climatic factors; Altitudinal patterns
国家水体污染控制与治理科技重大专项(No.2015ZX07203-011;No.2015ZX07204-007)。
张东杰(1992- ),男,安徽芜湖人,硕士研究生,研究方向:生态数学。*通讯作者,教授,博士,博士生导师,从事理论生态学研究。
2016-09-09
Q 948
A
0517-6611(2016)32-0053-04
