浅海筏式养殖对周边海域潮流和悬浮体特征影响研究*
2016-12-12张泽华黄海军刘艳霞严立文王恩康
张泽华,黄海军,刘艳霞,严立文,王恩康
(1.中国科学院 海洋研究所,海洋地质与环境重点实验室,山东 青岛 266071;2.中国科学院大学 北京,100049;3.国家海洋局 第一海洋研究所,山东 青岛 266061)
浅海筏式养殖对周边海域潮流和悬浮体特征影响研究*
张泽华1,2,黄海军1*,刘艳霞1,严立文1,王恩康3
(1.中国科学院 海洋研究所,海洋地质与环境重点实验室,山东 青岛 266071;2.中国科学院大学 北京,100049;3.国家海洋局 第一海洋研究所,山东 青岛 266061)
根据黑泥湾近岸海域养殖期与无养殖期多站同步分层潮流泥沙实测资料,对黑泥湾大规模海带养殖区及周边海域的潮流动力特征以及悬浮体分布规律和时间变化进行了动态的研究。通过对比海带养殖期与无养殖期潮流动力与悬浮体特征的时空变化规律,分析了浅海海带筏式养殖对周边海域悬浮体动力特征的影响作用。研究结果表明,研究区大范围分布的海带筏式养殖群落使得潮流动力减弱,并改变了潮流流速和悬浮体质量浓度的垂向分布特征,海底边界层的潮流底切应力被削弱,抑制了海底泥沙的再悬浮作用,但在一定程度上加强了水体的垂向混合作用。
筏式养殖;水动力;泥沙输运;悬浮体;黑泥湾
海带是世界上养殖面积和产量最大的大型经济藻类。近年来,随着大规模海水养殖技术的发展与需求的不断扩大,我国近海海带养殖的范围与规模逐年扩大,所以人工海水养殖对于浅海地区环境的影响日益引起人们的重视。大型海藻群落内的水动力的研究观测始于Jackson和Winant[1],他们通过在美国加利福尼亚圣迭戈湾海藻床内、外的海流观测,发现海藻群落内的流速仅为外部的 1/3。Jackson[2]在之后的研究中发现,在海岸地区的海藻床与周边海域的物质交换中,横向流比沿岸流起到更重要的作用。随后很多学者[3-8]相继通过野外调查与室内试验的方法对海藻群落内的水动力特征进行了研究,揭示了海藻床的存在对潮流、波浪、湍流以及物质输运的影响。这些研究都针对河口浅海区海藻床的研究,而筏式养殖群落作为一种悬浮的植被存在于水体的中上层,其对水动力与物质输运的影响和附底的海藻群落相比存在很大的差别。Grant等[9]观测了萨尔达尼亚湾内贻贝筏式养殖对海水的阻力作用,通过研究对比发现筏式养殖阻力约为底播养殖的 30倍,Boyd和Heasman[10]的研究进一步验证了这个结果。Plew等[11-12]通过实地观测新西兰黄金湾附近离岸贻贝筏式养殖区内水动力特征的水平及垂向变化,研究了悬浮冠层对层流的影响作用。樊星等[13-14]根据实测资料研究分析了桑沟湾养殖活动影响下的潮流动力结构,并建立了一维双阻力水动力模型对潮流垂直结构及其相应的剪应力垂直结构进行模拟,通过控制模型参数研究了养殖活动对潮流垂直结构的影响机制。目前对海区水动力环境变化的研究主要针对于对养殖容量的估算,而很少涉及大规模的养殖活动造成的悬浮泥沙输运及其动力机制。本文以山东半岛东南端的典型大规模海带筏式养殖区——黑泥湾为研究对象,根据不同养殖时期近岸海域多站同步分层潮流泥沙实测资料,对该海域海带筏式养殖区及周边海域的潮流以及悬浮体分布特征进行了研究。通过对比海带养殖期与无养殖期水动力与悬沙浓度的时空变化规律,分析了浅海海带筏式养殖对周边海域悬浮体输运的影响作用。
1 数据与方法
1.1 研究区概况
黑泥湾地处山东省威海市,位于山东半岛东南端楮岛和镆铘岛之间,东向濒临黄海。黑泥湾为基岩岬湾海岸,由于其两侧基岩岬角突出,将其与其他海湾分隔,形成了相对独立的泥沙运动系统,其北部褚岛岬角北侧为桑沟湾,那里高密度养殖着牡蛎、贝类、鱼和海带等[15];西南部镆铘岛西侧为海带养殖为主的石岛湾。黑泥湾顶海岸主要为连岛和潟湖沙坝堆积地貌类型,侵蚀主要发生在岬角处[16]。湾中部有礁石分布,坡度较陡,10 m等深线直逼海岸,其将海湾分为南北两个水下地形坡度较缓的海湾,坡度小于0.5‰,向海过渡至水深20 m附近有个地形突变的水下岸坡[17-18]。
黑泥湾附近海域的潮汐类型主要为不规则半日潮,日潮不等现象显著,多年平均潮差2.39 m。潮流性质为规则半日潮流,具有明显的往复流性质,涨潮偏南向、落潮偏北向。研究区夏季常浪向为S和SW向,冬季多为N向,强浪向主要为S、SE、SSE及NNE向,平均波高为0.6~1.2 m,最大波高为4~7 m。研究区近岸为自北向南的黄海沿岸流区[19]。黑泥湾外围河流多为单独入海的小河,其对黑泥湾的泥沙供应量很小。海岸潮滩沉积物类型为粉砂质砂,海域沉积物类型主要为黏土质粉砂,为第四纪泥质沉积区[20]。
黑泥湾所属的荣成市素有“中国海带之乡”称号,是全国最大的海带养殖地,目前海带养殖区面积已达七千多公顷,鲜海带产量约占全国的50%[21]。黑泥湾附近5~20 m等深线范围内的海域均为海带养殖架群填充,养殖面积约30 km2。黑泥湾海带养殖周期一般为11月至次年的5或6月,离岸较远的水域最晚可至7月。到12月时,黑泥湾海域表层海水已被海带群落以及养殖筏架覆盖,此时海带筏式养殖分布范围如图1所示,而到6-7月海带收割时,海带已可以生长到4~5 m长。图1养殖范围基于5 m分辨率的SPOT5假彩色合成影像提取得到[22],影像获取时间为2004-12-08,通过对比研究区相近几年的遥感影像,海带养殖范围无太大变化,因此影像提取的养殖范围具有很好的代表性。
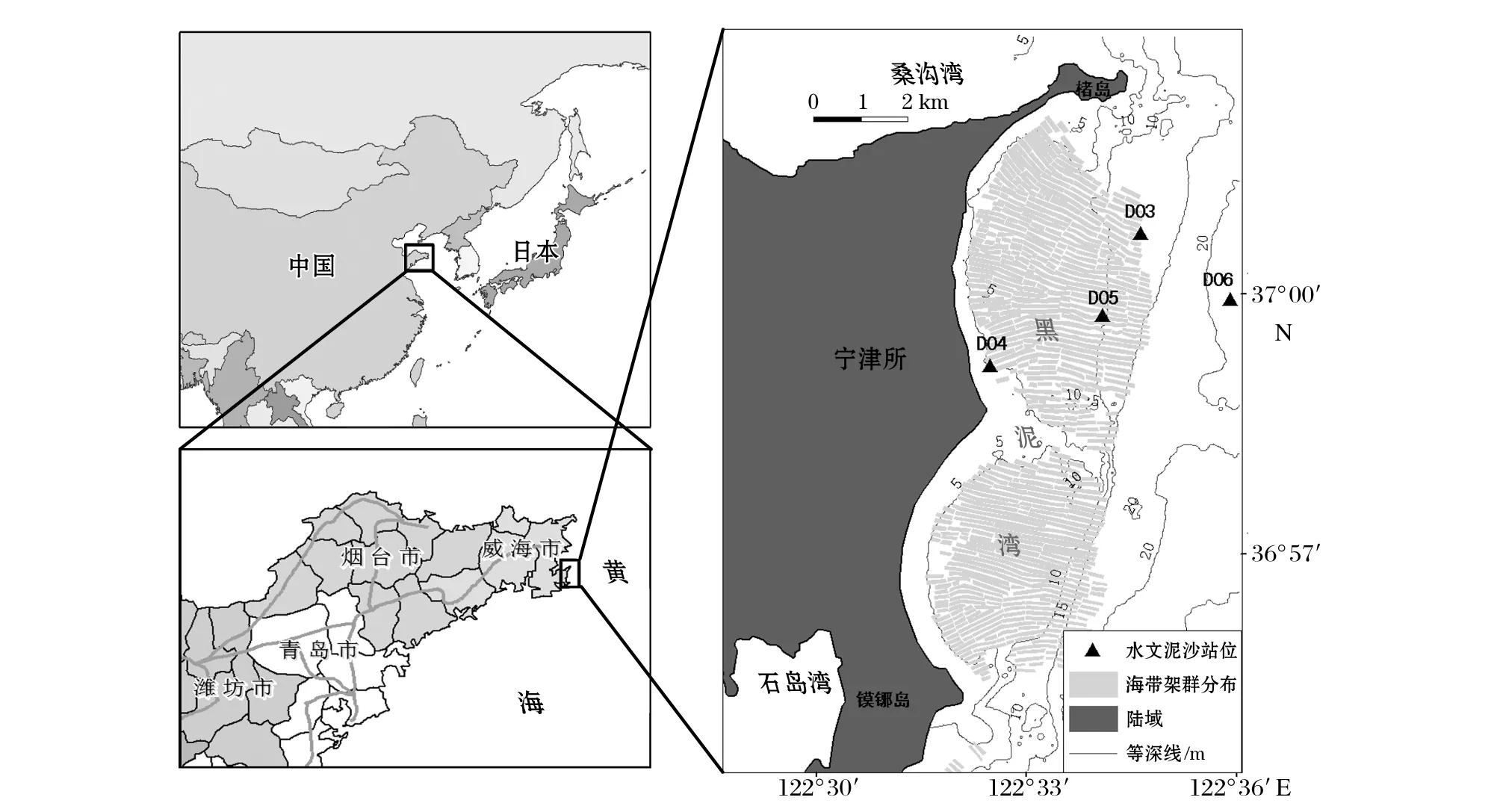
图1 研究海区位置及取样站位布置图
1.2 现场观测与采样
为对比探讨浅海筏式养殖对潮流动力以及悬浮体输移的影响,在研究海区布置了4个潮流、泥沙观测站位,分别在2006-07和2006-12进行大、中、小潮共6个潮周期的定点同步分层潮流、泥沙观测,用于研究养殖与非养殖期海域流场与泥沙运动特征,观测站位布置见图1。2006-07调查时,除外海一小范围海域(D03站位)尚余海带之外,黑泥湾大部分海域海带已收获,D04,D05和D06站位均可视为无养殖影响的自然海域。2006-12调查时,4个站位分别位于海带养殖区外缘(D03)、养殖区内缘(D04)、养殖区中部(D05)和养殖区外围的自然海域(D06)。7月和12月观测期间,最大风速分别为3.2和5.7 m/s,天气均状况较好。
具体观测与取样、分析方法:在连续潮周期内的整点时刻,采用挪威生产的安德拉RCM-9MKII型和RCM4s型自容式海流计,以五点测量法分5层(表层、0.2H、0.6H、0.8H和底层,H为整点时刻实测水深)观测流速、流向数据,并使用直立式采水器同步采集双水样进行室内粒度测试和含沙量分析。另外在大、中、小潮落憩时刻取各观测站的底质样品。水样在每个潮次观测结束后现场过滤,在实验室中完成滤膜烘干、称重、含沙量计算、盐度计算,使用的滤膜孔径为0.45 μm,直径为60 mm。对涨急、落急、涨憩、落憩等典型时刻的悬浮体样品以及各站位的底质样品进行了粒度分析,分析步骤为:将取回的水样过滤,过滤出的悬浮颗粒物加入六偏磷酸钠在超声波振荡仪内振荡、分散2 h,然后采用Cilas 940L型室内激光粒度仪进行粒度分析。
2 结果与分析
2.1 研究海区冬夏两季潮流特征
研究海区各站实测潮流均表现出较强的往复流性质,主流向基本与岸线平行,涨潮流偏S向,落潮流偏N向。总体上涨潮平均流速大于落潮平均流速,且涨落潮平均流速大潮期>中潮期>小潮期。
由各站位冬夏季流速特征值(表1)可以看出,研究海区整体上由外海向近岸随水深变浅潮流平均流速逐渐降低,四个调查站位中D06站位(平均水深约24 m)平均流速最大,D04站位(平均水深约为6.5 m)平均流速最小,D03、D05站位水深均为12 m,平均流速介于D04、D06两者之间。比较各站位的冬夏季平均流速差别,D03、D06站位冬季平均流速均大于夏季,而位于养殖区内的D04、D05站位冬季平均流速均小于夏季。
由研究海区各站位各层潮流流速的垂向变化来看(图2),D06站位分层时均流速在冬夏两季均表现出近似对数剖面的流速分布特征,海水表层流速最大,冬季为51.7 cm/s,夏季为52.3 cm/s,随水深增加而逐渐减小,近底层的平均流速冬季为36.0 cm/s,夏季为23.7 cm/s。表层最大流速冬季为112 cm/s,夏季为132 cm/s;底层最大流速冬季为96 cm/s,夏季为61 cm/s。D04、D05两站位分层时均流速在夏季时也表现出这种流速剖面特征,表层分别为14.0和35.4 cm/s;而在冬季时由于表层养殖阻力的存在流速剖面发生变化,平均流速最大值出现在水深中部0.6H~0.8H,其中D04站位出现在水深4 m左右,流速约为9.9 cm/s,D05站位出现在水深8 m左右,流速约为24.7 cm/s。两站最大流速剖面特征与各自时均流速剖面特征相似,D04冬季流速最大值为22 cm/s,D05冬季流速最大值为44 cm/s,分别出现在水深4和8 m左右。D03站位分层时均流速在夏季表现出中层高,表、底层低的特征,最大值约为24.5 cm/s,出现在水深9 m左右。

表1 研究海区各站位冬夏季流速特征值(cm·s-1)比较Table 1 The characteristic values of current velocity (cm·s-1) at each sampling station in summer and winter
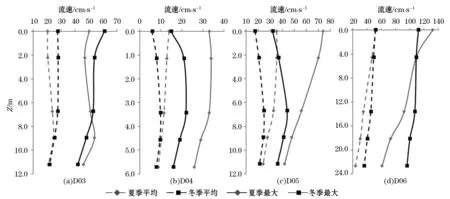
图2 研究海区各站位流速剖面特征图
2.2 研究海区冬夏两季悬浮体分布特征
表2为研究海区各调查站位冬夏季垂向平均含沙量变化范围,由表可以看出,D04站位水深较浅,冬夏两季悬浮体质量浓度值均较低,夏季平均为14.5 mg/L,冬季平均为10 mg/L。另外3个站位的悬浮体质量浓度冬夏季的差异更为明显,在夏季D03,D05和D06站位悬浮体质量浓度平均值相差不大,分别为26.7,29.0和23.1 mg/L,D05站位略高于其他两个站位;而在冬季,D06站位的悬浮体质量浓度明显高于D03和D05站位,前者平均值为37.5 mg/L,后两者分别为21.2和22.2 mg/L。夏季调查期间各站位的含沙量小潮期>大潮期>中潮期;而冬季的含沙量大潮期>中潮期>小潮期。各站位冬夏季垂向平均含沙量对比,D06站位冬季大于夏季,其他3个站位均是夏季略大于冬季。
研究海区悬浮体质量浓度垂向分布在冬夏两季也存在着很显著的差异,主要表现为夏季比冬季悬浮体质量浓度垂向变化梯度更为明显(图3)。其中D05站位冬夏差异最为显著,夏季表层、底层悬浮体质量浓度分别为16.0和63.3 mg/L;冬季分别为20.7和24.5 mg/L。D06站位夏季表层、底层悬浮体质量浓度分别为9.5和43.6 mg/L;冬季分别为34.8和40.2 mg/L。

表2 研究海区各站位冬夏季垂向平均含沙量(mg·L-1)比较Table 2 The depth-averaged silt content (mg·L-1) at each sampling station in summer and winter

图3 研究海区各站位时均悬浮体质量浓度垂向变化图
3 讨 论
3.1 研究海区潮流流速特征季节性差异
为了研究筏式养殖活动对海域潮流特征的影响而选取了2个养殖季节进行研究,然而海域自然状态下的水动力特征本身不可避免地也存在一定的季节差异,因此分析养殖活动的影响之前应首先排除季节性差异造成的水动力的变化。
D06站位调查结果可以很好地反映研究区海域自然状态下潮流流速垂向分布特征的季节性变化。从平均流速的变化上来看,研究区海域观测期间冬季流速略大于夏季流速。冬、夏两季海水表层平均流速相近,而底层平均流速冬季略大于夏季,夏季比冬季具有更大的流速梯度。在夏季,海水的层化作用较强,阻碍了海水的垂向混合,致使流速剖面具有较大的梯度;而冬季则相反,由于海面风混合作用、海面冷却造成的垂向对流以及潮流在底边界层内的强湍流混合作用等的结果,使得水体垂向上混合均匀,从而使水体垂向各层的流速差别不大[21]。
3.2 筏式养殖对潮流的影响
由D04、D05站位冬、夏两季不同潮期潮流流速对比发现,冬季海带养殖时期潮流流速相比较夏季无养殖时期发生很明显的衰减,D04站位夏季和冬季平均流速分别为11.3和8.0 cm/s,D05站位夏季和冬季平均流速分别为31.7和22.2 cm/s,两站位平均流速冬季比夏季均降低了约30%(图2)。而由于养殖筏架和海带群落主要集中在海水表层(观测期间平均水深小于1 m),这种流速的衰减在水体表层最为明显,D04站位夏季和冬季表层平均流速分别为14.0和6.0 cm/s,D05站位夏季和冬季平均流速分别为35.4和17.4 cm/s,两站位表层平均流速冬季比夏季降低了50%~60%。随着水深的加深影响逐渐减小,0.2H层平均流速冬季比夏季降低了40%左右,0.6H层平均流速降低20%左右,0.8H和底层平均流速冬夏季差值在15%以内。
从各站位潮流流速的垂向分布来看,对比海带群落内部和外部,D06站位冬季和夏季以及D04和D05站位夏季,潮流各层平均流分布符合对数变化的剖面形态;而冬季时D04和D05站位分层平均流速在剖面存在中间层位流速大表层和底层流速小的特征(图2)。樊星等[13]建立了一种一维双阻力水动力模型从理论上研究了这种水流流速剖面模式,它从壁面率假设出发,将顶覆筏式海带养殖群落的摩擦阻力类比海底边界层的摩擦阻力,得到一种双对数分布形式的流速剖面。由图2中D04和D05站位流速剖面分布特征可以看出,虽然养殖筏架和海带群落主要集中在海水表层(观测期间平均水深小于1 m),但是海表养殖阻力形成的顶边界层的影响可以深入到0.6H~0.8H水深处。
然而这种流速剖面仅适用于海带群落分布于海水表层的情况,冬季调查期间,D04和D05站位的流速分布即很好的反映了这种分布特征,但是,当海带群落充分生长并覆盖了海水垂向较大深度范围的水体之后,海带群落造成的阻力作用影响到海水水体的中上部,此时流速剖面特征不再呈单纯的双对数分布特征。D03站位附近海域在夏季调查期间仍存留一定范围的海带群落,且海带大部分生长到4~5 m长度,此期间调查得出该站位时均流速剖面呈S型分布(图2),即海水时均流速由海表层至底层呈减小—增大—再减小的变化特征。
图4是D05站位冬季大潮和D03站位夏季大潮半个潮周期的潮流流速北分量垂向分布,时间间隔为1 h,流速北方向为正。由图可以看出各时刻的潮流流速垂向分布模式与相应的时均流速剖面模式是一致的(图2),即D05站位在冬季流速遵循双对数剖面模式,D03站位在夏季流速遵循S型剖面模式。D05站位冬季时各层海水几乎同时涨落,说明海带藻体在约2 m长的时期对海域表层水体的阻力与海底的摩擦作用量级相当。而在D03站位,底层海水先涨先落,表层海水后涨后落,海带已生长至4~5 m长,体积增大,其对表层海水的阻力作用将大于海底摩擦作用,从而影响水流的垂向结构。

图4 大潮半个潮周期内潮流流速北分量垂向分布变化
将研究海区各站位各潮期的实测海流资料经过磁差等订正后,进行潮流准调和分析[23],得出各站位各潮期主要分潮的潮流调和常数及椭圆要素。由于研究区海域潮流类型为规则半日潮流,因此其潮流运动形式主要以M2分潮流的椭圆率作近似分析。表3为D05站位冬夏季大潮期各层M2分潮流的椭圆率K值。由表可以看出,D05各层旋转率|K|值均为0.06~0.16,属典型的往复流,而且冬季有海带养殖群落存在的表层M2分潮流|K|值小于冬季其他层位以及夏季各层位,表明养殖群落使潮流的旋转性特征减弱而更趋向于往复流。可见冬季表层海带养殖群落的存在不仅可以改变潮流流速大小,而且还对潮流性质产生影响。

表3 D05站位冬夏季大潮期各层M2分潮流K值表Table 3 The ellipticity value (K) of M2 current component at each depth layer of the D05 station
3.3 研究区海域悬浮体特征季节性差异
在潮流为主要驱动力的浅海海域,无风无浪条件下潮致底应力被认为是沉积物周期性再悬浮的最重要的驱动力。半日潮流占优的水域悬浮物浓度升高通常对应着涨急落急时刻,并呈现出M4周期变化[24]。当沉积物进入水体后,水体湍流混合强度大小决定了沉积物是否继续随垂向湍流扩散到上层水体中去参与水平悬移输运,或者沉降下来回到沉积态或参与推移质输运。
由D06站位冬夏季时均悬浮体质量浓度垂向变化(图3)可以看出,研究区海域冬夏季悬浮体质量浓度垂向分布特征有着显著的差异。夏季悬浮体质量浓度值垂向变化很大,水体下部悬浮体质量浓度很高,底层悬浮体质量浓度平均值为42.6 mg/L,但上部浓度则较低,表层平均浓度为9.5 mg/L。而冬季悬浮体质量浓度各层分布均匀,表层和底层悬浮体平均浓度分别为34.8和40.2 mg/L。冬夏两季水体底层悬浮体平均浓度虽然相差不大,但水体悬浮体垂线平均浓度冬季(37.5 mg/L)要比夏季(23.1 mg/L)高很多。
自然状态下冬季水体垂向混合近乎均匀,水体中动量及悬浮颗粒物的垂向交换非常通畅,沉积物一旦被潮流卷起,立即在湍流垂向混合作用下扩散到水体中部甚至表层。然而在夏季,由于水体层化作用,悬浮颗粒物的垂向分布、聚集及湍扩散却是另一番景象,水体动量以及悬浮体质量浓度在垂向上具有很强的梯度分布特征。尽管海底沉积物的起动取决于起动临界应力,但其再悬浮进入水体中后垂向输运的高度和在水体中停留的时间取决于水体中湍扩散系数。如果水体中的湍流强度不足以克服悬浮物的重力沉降,悬浮颗粒物马上沉积下来。研究区海域D06站位位于距离养殖区较远的自由水体,实测结果表明该站位悬浮体垂向分布特征季节性变化符合以上规律,然而位于养殖区边缘及内部的几个站位的悬浮体分布季节特征存在一定的改变。
3.4 筏式养殖对悬浮体动力特征的影响
从悬浮泥沙来源分析,研究海区附近岸段无较大河流入海,且季节性较强,泥沙量较少,陆域来沙对整个区域的贡献较小。该海域海岸类型均为沙质,从波浪动力和海岸性质而言,海岸可能存在沿岸输沙,但从目前该岸线形状以及多年的海岸线和岸滩变化判断,沿岸输沙的量级应该很有限。研究区海域受潮流和波浪共同作用,在正常天气下主要受潮流影响,泥沙主要以悬移运动为主,在较大风浪条件下,近岸及破波带水域泥沙较易起动,并随潮流、波生流等运动。因此,就该海域而言,本地掀沙是其最主要的泥沙来源。
比较D03,D05和D06三个站位冬夏两季垂向平均悬浮体质量浓度随时间的变化(图5)可以看出,在夏季各站位悬浮物浓度升高几乎都对应着涨落急时刻,呈现M4周期变化特征,且3个站位的浓度值在相似的量级上变化,特别是D05、D06站位浓度变化过程具有很高的一致性。由此可见,研究区海域自然状态下潮致底应力是沉积物周期性再悬浮的最重要的驱动力。在冬季处于养殖区外部自然海域的D06站位保持较高的悬浮体质量浓度和较大的浓度变化波动,而D03和D05站位则受到养殖的影响,悬浮体质量浓度降低且在20~30 mg/L波动。另一方面,D06站位悬浮体质量浓度依然具有与潮流流速相关的M4型的周期变化特征,而D03和D05的浓度变化无明显的周期特征。
比较D03,D05和D06三个站位冬夏两季底层悬浮体质量浓度的时间变化,在夏季D05、D06站位底层悬浮体质量浓度在较高的范围波动,最大值可以达到130 mg/L左右,悬浮物浓度的高值均对应着涨落急时刻,可见在夏季自然海域潮流是泥沙再悬浮主要的动力因素。在冬季,处于自然海域的D06站位底层悬浮体质量浓度值波动剧烈,而受到筏式养殖影响的D03和D05站位底层悬浮体质量浓度较小、波动较弱。由于养殖阻力的作用使养殖区内部平均流速发生衰减,近底潮流流速剪切作用减弱抑制了泥沙的再悬浮过程,最终导致研究区冬季养殖区内底层悬浮体质量浓度较低且波动较弱的情况。同样地,由于夏季D03站位附近仍有一定范围养殖筏架的存在,在无风浪状态下海底层的泥沙再悬浮量减少,近底层的悬浮体质量浓度仅为20~40 mg/L。
在养殖区内部的D05站位,虽然悬浮体质量浓度与流速密切相关且表现出相同的周期性变化,在冬季夏季的相同潮周期内流速极大值都出现在涨急和落急时刻,但是在养殖季节悬浮体质量浓度峰值往往出现在涨憩和落憩时刻,说明悬浮体质量浓度的变化存在一定的滞后性,原因一方面是悬沙沉降至底床或从底床向上再悬浮需要一定时间,另一方面是流速从极大值开始降低后,仍有一段时间不低于再悬浮的临界速度,悬沙浓度会继续升高。通过对D05站位涨急、落急、涨憩、落憩等典型时刻的悬浮体粒度分析(表4),得到涨憩落憩时底层悬浮体中值粒径Md值偏小,涨急落急时底层悬浮体中值粒径Md值偏大,也可以说明这一特征。然而在冬季养殖期间,D05底层悬浮体质量浓度和粒径参数变化均没有很强的周期性和规律性,说明在冬季养殖期间,养殖区内的潮流流速衰减的情况下,潮流流速大小已不再是悬浮体质量浓度波动的控制因素。但是,本次调查中并未对波浪进行观测,无法说明波浪作用对养殖区内泥沙再悬浮的影响。

图5 研究海区各站位冬、夏两季悬浮体质量浓度变化图
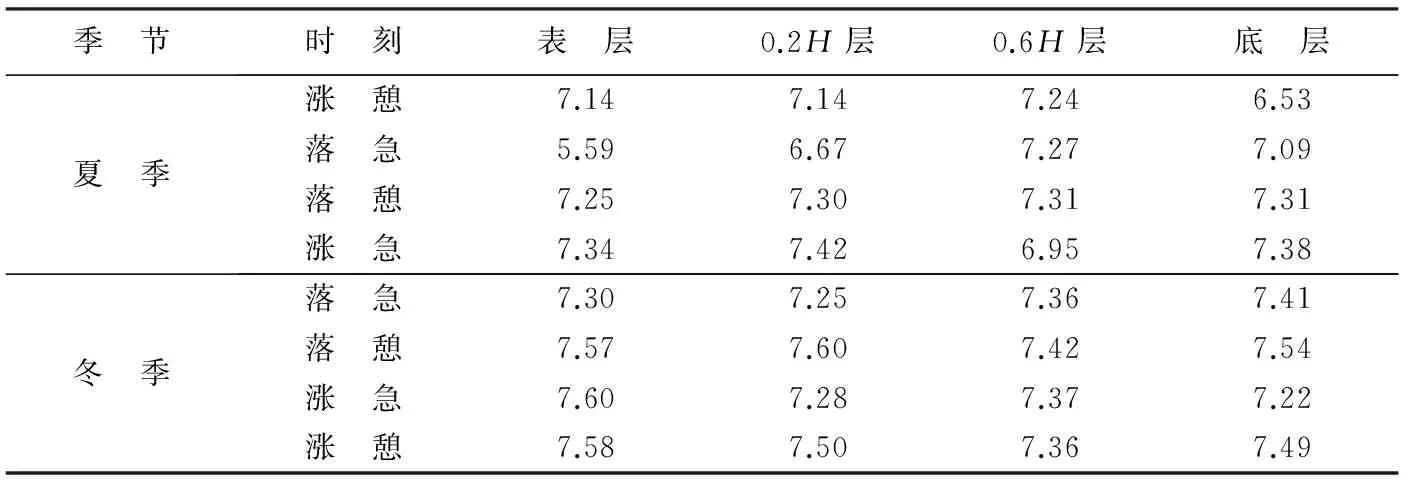
表4 D05站位冬夏季大潮典型时刻悬浮体中值粒径Table 4 The median size of SSC at the D05 station at typical moments of spring tide
由以上分析,研究区筏式养殖覆盖的水体,由于养殖活动的存在使得海域流速明显衰减,致使水底潮流切应力相应减弱,从而造成再悬浮作用减弱;相比于自然水域,其近底层以及垂向平均悬浮体质量浓度均显著降低。但是与夏季无养殖期间水体各层悬浮体质量浓度存在较大浓度梯度不同的是,D05站位在冬季养殖期间大部分时间悬浮体质量浓度垂向分布是十分均匀的,而且调查期间冬季表层水体悬浮体平均浓度(20.7 mg/L)反而略大于夏季(16.0 mg/L)(图3)。这种冬季水体悬浮体质量浓度垂向均匀分布的特征与自然水域(D06站位)的垂向分布特征是一致的。而比较夏季时D03和D05站位悬浮体垂向分布差别,底层悬浮体平均浓度前者远低于后者的同时,表层悬浮体平均浓度前者略大于后者。这说明了养殖区内的垂向混合作用并不只是季节性原因造成的,筏式养殖对水体的垂向混合也有重要作用。研究区夏季自然状态下水体层化作用较强,而养殖区内却存在很强的混合作用,这可能是由于筏式养殖增强了水体的湍流扩散作用。在一般的近岸环境中,顶边界层和底边界层中流速的剪切作用产生的湍流是湍流生成的主要的机制。Rosman等[7]通过对海藻床内外理查森数(Ri)的研究,指出海藻群落内部由于流速降低进而速度剪切显著减弱,以至于流速剪切不足以超过局部层化作用而产生垂向的混合。然而海带群落内部却存在着湍流生成的其他机制,可能包括单个海带束与潮流相互作用而在其后部产生湍流[25-26]。另一方面,与海藻床不同的是,悬浮式的海带养殖主要集中于水体的上半部分,而海带养殖之下存在一定深度相对自由的水体,这部分水体收到养殖阻力的影响较小而表现出相对较高的流速。Waite[27]对贻贝养殖区内流速的观测发现养殖体以下2 m存在强烈的水流波动,指出近表层阻滞水流与养殖区以下较快速的水流之间形成了一个混合层。这种混合层会加强水体的垂向混合作用。由此可见,筏式养殖群落在一定程度上加强了水体的垂向混合作用。
3.5 筏式养殖对冲淤演化的影响
严立文[28]利用海图以及实测水深数据建立了研究海区的DEM并计算出该海域两个不同历史时期的底床冲淤演化特征(图6)。由于1949-1980年期间研究区海域受养殖活动的扰动较少,基本上反映了海域自然演化的状态。由该时期冲淤演化特征看出,研究区湾外水深15 m以深海域以侵蚀作用为主,而湾内水深15 m以浅海域则以淤积作用为主。在自然状态下,研究海区冲淤演化特征受到沉积物水动力及地形地貌条件的综合作用,湾外水域在较强的水动力条件下,沉积物很容易发生再悬浮,由潮流作携带向近岸扩散的过程中,随水深变浅、水动力减弱而逐渐落淤。80年代以后筏式养殖活动大规模开展,研究海区大范围分布的海带群落减弱了湾内的水动力,增加了泥沙沉积的同时减弱了再悬浮作用,海带养殖区外围水深15~20 m海域表现最为明显,因为外海携带来的泥沙首先在此处发生淤积。
然而研究海区的筏式养殖并非全年覆盖水域,海带群落存在一个生长周期,由于海带群落密度、影响深度和覆盖率的不断变化,其对水动力和悬浮体扩散的影响是一个复杂的过程,同时也将影响研究海区冲淤状态的年内变化。11月至次年6或7月海带养殖期间,由外海搬运来的泥沙在养殖区外缘发生沉降淤积,而养殖区内部由于处于弱水动力状态,泥沙很难进入缺乏补给,本身底质又较少发生扰动再悬浮,所以处于一种较稳定的冲淤平衡状态。而海带收获以后直到11月新的养殖活动开始这段时间,研究海区处于自然状态,于是在养殖期间于养殖区外缘淤积的大量沉积物开始在水流的作用下往浅水区搬运。如此周期往复的过程最终形成了图6所示的冲淤格局。

图6 研究海区不同历史时期的冲淤演化图[28]
4 结 论
通过对比黑泥湾近岸海域海带养殖期与无养殖期潮流动力与悬浮体特征的时空变化规律,分析了浅海海带筏式养殖对周边海域悬浮体动力特征的影响作用,得出以下结论:
1)研究区自然状态下冬夏两季潮流流速在潮周期内的变化几乎一致,而在养殖季节,海带群落内部潮流流速发生很明显的衰减,平均流速冬季比夏季均降低了约30%,表层平均流速冬季比夏季降低了50%~60%。
2)研究区冬季海带养殖群落仅覆盖水体表层,时均流速剖面表现出明显的双对数剖面模式,而随着海带的生长,覆盖的水体范围加深,流速剖面转化为S型剖面模式。海带养殖群落的存在不仅可以改变潮流流速大小,而且还对潮流性质产生一定影响。养殖群落覆盖的表层水体潮流的旋转性减弱而更趋向于往复流。
3)筏式养殖的存在,使海底边界层的潮流底切应力被削弱,抑制了海底泥沙的再悬浮作用,使得底层悬浮体质量浓度值大大降低且波动减弱。筏式养殖群落的存在一定程度上加强了水体的垂向混合作用,使得养殖海域的悬浮体垂向分布更为均匀。
4)研究区大范围分布的筏式养殖群落减弱了湾内的水动力,使得悬浮泥沙更容易沉降淤积,近岸水域特别是养殖区外围的淤积作用增强。
[1] JACKSON G A, WINANT C D. Effect of a kelp forest on coastal currents[J]. Continental Shelf Research, 1983, 2(1): 75-80.
[2] JACKSON G A. Currents in the high drag environment of a coastal kelp stand off California[J]. Continental Shelf Research, 1998,17(15):1913-1928.
[3] ELWANY M, REILLY W O, GUZA R, et al. Effects of southern California kelp beds on waves[J]. Journal of Waterway, Port, Coastal, and Ocean Engineering, 1995,121(2): 143-150.
[4] UTTER B, DENNY M. Wave-induced forces on the Giant Kelp Macrocystis pyrifera (Agardh): field test of a computational model[J]. The Journal of Experimental Biology,1996, 199: 2645-2654.
[5] LOVAS S M, TORUM A. Effect of the kelp Laminaria hyperborea upon sand dune erosion and water particle velocities[J]. Coastal Engineering, 2001, 44(1):37-63.
[6] GAYLORD B, DENNY M, KOEHL M. Modulation of wave forces on kelp canopies by alongshore currents[J]. Limnology and Oceanography, 2003, 48(2): 860-871.
[7] ROSMAN J H, KOSEFF J R, MONISMITH S G, et al. A field investigation into the effects of a kelp forest (Macrocystis pyrifera) on coastal hydrodynamics and transport[J]. Journal of Geophysical Research, 2007, 112, C02016, doi:10.1029/2005JC003430.
[8] GAYLORD B, ROSMAN J H, REED D C, et al. Spatial patterns of flow and their modification within and around a giant kelp forest[J]. Limnology and Oceanography, 2007, 52(5): 1838-1852.
[9] GRANT J, STENTON-DOZEY J, MONTEIRO P, et al. Shellfish culture in the Benguela system: a carbon budget of Saldanha Bay for raft culture of Mytilus galloprovincialis[J]. Journal of Shellfish Research, 1998, 17: 41-49.
[10] BOYD A J, HEASMAN K G. Shellfish mariculture in the Benguela System: water flow patterns within a mussel farm in Saldanha Bay, South Africa[J]. Journal of Shellfish Research, 1998, 17:25-32.
[11] PLEW D R, STEVENS C L, SPIGEL R H, et al. Hydrodynamic implications of large offshore mussel farms[J]. IEEE Journal of Oceanic Engineering, 2005, 30(1):95-108.
[12] PLEW D R, SPIGEL R H, STEVENS C L, et al. Stratified flow interactions with a suspended canopy[J]. Environ Fluid Mech, 2006, 6:519-539.
[13] FAN X. Preliminary studies on the features of tidal-dynamic structure in a typically high density mariculture coastal bay-observation and simulations[D].Qingdao: Ocean University of China, 2008. 樊星.典型养殖海区潮动力结构特征的初步研究--观测与数值模拟[D].青岛:中国海洋大学,2008.
[14] FAN X, WEI H, YUAN Y, et al. Vertical structure of tidal current in a typically coastal raft-culture area[J]. Continental Shelf Research, 2009, 29: 2345-2357.
[15] GRANT J, BACHER C. A numerical model of flow modification induced by suspended aquaculture in a Chinese bay[J]. Journal of Fisheries and Aquatic Sciences, 2001,58:1003-1011.
[16] The editorial committee for records of bays in China. Records of bays in China: Volume 3[M].Beijing: China Ocean Press,1993.中国海湾志编纂委员会.中国海湾志:第三分册[M].北京:海洋出版社,1993.
[17] LIU J P, MILLIMAN J D, GAO S, et al. Holocene development of the Yellow River's subaqueous delta, North Yellow Sea[J]. Marine Geology, 2004, 209: 45-67.
[18] YANG Z S, LIU J P. A unique Yellow River-derived distal subaqueous delta in the Yellow Sea[J]. Marine Geology, 2007, 240: 169-176.
[19] LIU J, SAITO Y, KONG X H, et al. Geochemical characteristics of sediment as indicators of post-glacial environmental changes off the Shandong Peninsula in the Yellow Sea[J]. Continental Shelf Research, 2009, 29: 846-855.
[20] HU L M, SHI X F, GUO Z G, et al. Sources, dispersal and preservation of sedimentary organic matter in the Yellow Sea: The importance of depositional hydrodynamic forcing[J]. Marine Geology, 2013, 335: 52-63.
[21] WANG G G. Rongcheng mariculture status and standing development countermeasures[D]. Qingdao: Ocean University of China, 2009. 王国钢.荣成海水养殖现状与可持续发展对策[D].青岛:中国海洋大学,2009.
[22] LIU X, HUANG H J, YANG X G, et al. Method to extract raft-cultivation area based on SPOT image[J]. Science of Surveying and Mapping, 2013, (38): 41-43.刘晓,黄海军,杨曦光,等.基于SPOT影像的筏式养殖区提取方法研究[J]测绘科学,2013, (38): 41-43.
[23] CHEN Z Y. Tidology[M]. Beijing: China Science Press,1980.陈宗镛.潮汐学[M].北京:科学出版社,1980.
[24] YUAN Y. Observations of suspended sediment dynamics in Chinese coastal seas by acoustic instruments[D]. Qingdao: Ocean University of China, 2009. 原野.基于声学方法的中国近海沉积物和悬浮颗粒物动力过程观测研究[D].青岛:中国海洋大学,2009.
[25] NEPF H M, MUGNIER C G, ZAVISTOSKI R A. The effects of vegetation on longitudinal dispersion[J]. Estuarine, Coastal and Shelf Science, 1997, 44(6):675-684.
[26] NEPF H M. Drag, turbulence, and diffusion in flow through emergent vegetation[J]. Water Resources Research, 1999, 35(2): 479-489.
[27] WAITE R P. The nutritional biology of Perna canaliculus with special reference to intensive mariculture systems[D]. Christchurch:University of Canterbury,1989.
[28] YAN L W. Sedimentary environment evolution in representative kelp (Laminaria Japonica)-cultured region(Harny Bay) and inner hydrodynamic mechanism[D].Qingdao: Institute of Oceanology, Chinese Academy of Sciences, 2008.严立文.浅海区海带养殖的沉积环境效应及动力机制——以山东半岛黑泥湾为例[D].青岛:中国科学院海洋研究所,2008.
Received: March 25, 2015
Effects of Suspended Culture of the Seaweed Laminaria japonica Aresch on the Flow Structure and Suspended Sediment Transport
ZHANG Ze-hua1,2, HUANG Hai-jun1, LIU Yan-xia1, YAN Li-wen1, WANG En-kang3
(1.KeyLaboratoryofMarineGeologyandEnvironment,InstituteofOceanology,ChineseAcademyofSciences,Qingdao 266071, China; 2.UniversityofChineseAcademyofSciences, Beijing 100049, China;3.TheFirstInstituteofOceanography,SOA, Qingdao 266061, China)
Based on the synchronous hydrological and sediment in-situ observations at fixed stations in Heini Bay during culture period and non-culture period, we studied the hydrodynamic characteristics and the distribution and variation of SPM in the Bay. By comparing the hydrodynamic and suspended sediment characteristics between culture and non-culture periods, the paper described the influence of suspended kelp aquaculture on the suspension dynamic characteristics of surrounding areas. The analytical results showed that widespread distribution of suspended culture community in the study area decreased the current velocity in culture area, and changed the suspension concentration vertical distribution structures. At the same time, the bottom shear stress of seabed boundary layer is greatly weakened, which restrains the seabed sediment re-suspension and thereby to strengthen the vertical mixing action of water to a certain extent.
suspended culture; hydrodynamics; sediment transport; SPM; Heini Bay
2015-03-25
国家自然科学基金项目——浅海大规模海带养殖活动对沉积环境的影响研究(41076031)和悬浮颗粒矿物成分对近岸水体光学参数的影响及光谱响应研究(41306190)
张泽华(1988-),男,山东济宁人,博士研究生,主要从事海洋环境地质方面研究.E-mail: zew00r@163.com*
黄海军(1963-),男,湖南湘潭人,研究员,博士,主要从事海洋遥感和海洋环境地质方面研究. E-mail: hjhuang@ms.qdio.ac.cn
(陈 靖 编辑)
P73
A
1671-6647(2016)01-0037-13
10.3969/j.issn.1671-6647.2016.01.004
