紫花地丁二型性种子的萌发特性及其生态意义
2016-12-09侯云云
苏 雪,高 婷,侯云云,2,吴 琼,孙 坤
(1.西北师范大学生命科学学院,甘肃 兰州 730070; 2.富平中学,陕西 渭南 711700)
紫花地丁二型性种子的萌发特性及其生态意义
苏 雪1,高 婷1,侯云云1,2,吴 琼1,孙 坤1
(1.西北师范大学生命科学学院,甘肃 兰州 730070; 2.富平中学,陕西 渭南 711700)
紫花地丁(Violaphilippica)具有开放花(CH)和闭锁花(CL)形成的二型性种子。在对紫花地丁CH种子和CL种子形态观测的基础上,比较了这两种类型种子在不同温度、光照以及贮藏时间下的萌发特性。结果表明,1)CH种子长、宽以及千粒重显著大于CL种子(P<0.05)。2)CH种子和CL种子最适萌发温度均为20 ℃,萌发率均在95%以上,在10 ℃条件下均不萌发,但在高温(30 ℃)条件下CL种子的萌发率显著高于CH种子(P<0.05),其它温度下,两者萌发率均无显著差异(P>0.05)。3)CH种子对光照条件敏感,黑暗条件对其萌发具有极显著的抑制作用(P<0.01),而CL种子在12 h光照和全黑暗条件下萌发差异不显著(P>0.05)。4)CH种子贮藏210 d后,萌发率显著降低(P<0.05),而CL种子则在贮藏510 d后萌发率才显著下降(P<0.05)。CH种子于春季成熟后即可迅速萌发,CL种子一部分于之后的季节萌发,另一部分成为短暂土壤种子库于次年春季萌发。紫花地丁通过二型性种子更好地适应时空异质性环境,提高了其适合度。
紫花地丁;种子二型性;萌发特性;生态适应
种子萌发阶段是植物种群更新和物种得以繁衍的关键性环节[1-2],但同时种子繁殖所产生的幼苗更易受不利环境的影响,因而具有更高的死亡风险[3]。所以,种子萌发阶段也是植物生活周期中最脆弱的阶段[4]。植物表现出复杂多样的萌发行为和策略,来提高物种的适合度。种子异型性(seed heteromorphism)是植物在种子阶段生态适应的一种特殊策略,它指的是同一植株不同部位产生两种或多种形态或行为的种子的现象[5-6],不同类型的种子在形态学特征、传播特点、萌发和休眠特性等方面往往存在明显差异[7-13]。近年来,种子异型性已成为国际领域植物适应对策研究中的热点问题和植物进化生态学的重要研究领域[14]。目前,我国对种子异型性的研究才刚刚起步,相关的研究报道较少[8-13],且多集中在荒漠、盐渍等极端环境下一年生植物类群中,对于一些非极端环境、特别是对以不同繁育策略形成的二型性种子的植物类群的相关研究鲜有报道。
紫花地丁(Violaphilippica)属堇菜科堇菜属的多年生草本植物,可形成开花受精(chasmogamous)花(CH)和闭花受精(cleistogamous)花(CL)两种有性繁殖的混合繁育系统[15]。CH花为虫媒花,花期为3月初到4月末;CL花无花瓣,为专性自交的完全闭锁花,4月中旬产生持续到9月中旬。紫花地丁的两种类型的花分别形成CH种子和CL种子[15-16],属典型的种子二型性植物。两种类型的种子通过蒴果开裂机械弹射得以传播,由于种子表面具有油质体,之后可通过蚁类的搬运实现二次传播[17-18]。虽然关于紫花地丁种子萌发有过少量研究报道[15,19-22],但并未对其二型性种子萌发特性进行过较系统的研究,更缺乏对CH和CL种子在萌发特性方面是否存在差异及其生态适应意义的了解。本研究对紫花地丁CH和CL种子的形态特征及萌发特性开展研究,以期了解:1)由CH花和CL花混合繁育系统形成的CH和CL二型性种子在形态和萌发特性上有何差异?2)这种形态和萌发特性的差异有何适应意义?这些工作将为紫花地丁生活史进化和种子异型性生态适应对策的研究提供参考依据。
1 材料与方法
1.1 试验材料
紫花地丁的CH和CL种子于2014年采自甘肃省兰州市安宁区党校内,在蒴果微裂但种子尚未弹出时采收。净种后自然风干,放于信封袋内,室温贮藏。随机选取风干后两种类型的种子各30粒,测量其长度和宽度。用电子天平进行称重,以1 000粒×3组的平均值为其千粒重。
1.2 试验方法
1.2.1 萌发试验 用0.3% KMnO4溶液对两种类型的种子进行表面灭菌10 min,蒸馏水冲洗3次后,将种子置于垫有两层滤纸的120 mm培养皿中进行萌发试验。每个处理3个重复,每个重复50粒种子。以胚根出现为种子萌发的标志,每隔24 h观察一次,统计萌发数,连续5 d不再有种子萌发视为萌发结束[23]。萌发结束后,计算各萌发指标:
萌发率=(发芽种子数/供试种子总数)×100%。
表示种子形成幼苗的潜力,衡量种子质量好坏或者休眠状态的指标。
萌发势=(前7 d萌发高峰期发芽种子数/供试种子总数)×100%。
萌发率反映种子萌发速度的快慢;
萌发指数=∑(Gt/Dt),反映种子发芽整齐度的指标。其中:Gt为t天的萌发数,Dt为相应萌发天数;萌发时滞(d):从种子着床到第一粒种子萌发所需天数;萌发历期(d):从第1粒种子萌发到萌发结束所需要天数。
1.2.2 温度处理 将新采集的CH种子和CL种子分别在10、15、20、25和30 ℃(光照12 h/黑暗12 h)5个恒温条件下进行萌发。
1.2.3 光照处理 在20 ℃恒温下,将新采集的CH和CL种子放于光照(光照12 h/黑暗/12 h)和黑暗(24 h)条件下进行萌发。24 h黑暗处理的种子于每天22:00进行观察,逐一取出培养皿迅速计数后放回培养箱。
1.2.4 贮藏时间处理 以新采收的种子为对照,将室温贮藏90、210、360和510 d的CH和CL种子在20 ℃恒温、12 h光照/12 h黑暗条件下进行萌发。
1.3 数据统计和分析
用Excel进行数据统计,萌发结果以萌发率±标准误差表示。用SPSS 20.0软件对数据进行正态分析和方差齐性检验,通过后对两种种子的长、宽以及千粒重进行独立样本t检验,对其它数据进行单因素方差分析。此外,采用二因素方差分析检验了种子类型与温度、光照以及贮藏时间对种子萌发率的交互作用。用Origin 9.0软件作图。
2 结果与分析
2.1 种子的形态特征
成熟的CH和CL种子色泽、形状等特征无明显差异,均为倒卵形、深褐色,种皮光滑有光泽。但CH种子长度、宽度及千粒重均显著大于CL种子(P<0.05)(表1)。
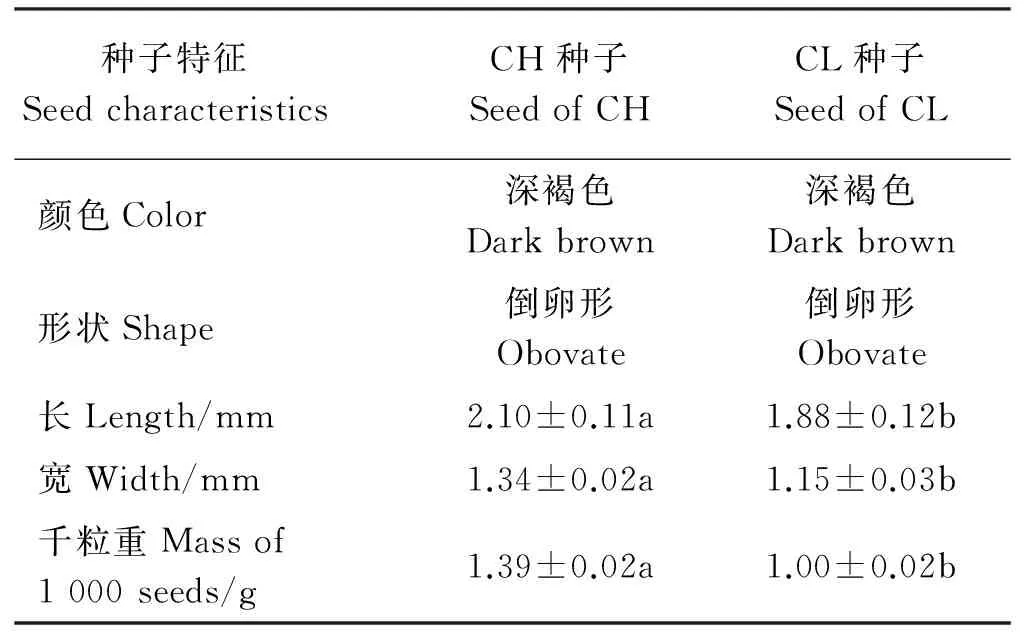
表1 紫花地丁开花授精(CH)和闭花受精花(CL)种子的特征Table 1 Characteristics of CH and CL seeds of V. philippica
注:同行不同小写字母表示两种种子间差异显著(P<0.05)。下同。
Note: Different lower case letters within the same row indicate significant difference between two seed types at 0.05 level. The same below.
2.2 温度对CH和CL种子的萌发特性的影响
紫花地丁新成熟的CH和CL种子的萌发率差异极显著(P<0.01);种子类型和温度对紫花地丁种子的萌发率有显著的交互作用(P<0.05)(表2)。随萌发温度(10~30 ℃)的升高,紫花地丁CH和CL种子萌发率都先升高后降低,最高萌发率均出现在20 ℃,分别可达到98.7%和96.0%(表3)。除在10 ℃条件下两种类型的种子均不萌发外,其它温度处理下,两种类型种子萌发率均大于50.0%。在30 ℃下,CL种子萌发率显著高于CH种子(P<0.05),其它温度下两者无显著性差异(P>0.05)(表3)。

表2 温度和种子类型对紫花地丁种子萌发率的影响Table 2 Analysis of variance for effects of temperature,types of seeds and their interaction on the germination rates of seeds of V. philippica
两种类型种子的萌发势和萌发指数在20 ℃均为最高,且与其它温度间差异极显著(P<0.01)(表3)。在30 ℃下CL种子萌发指数显著高于CH种子(P<0.05)。其它温度间,两种种子的萌发指数无显著差异(P>0.05)(表3)。
随萌发温度的升高,CH和CL种子萌发时滞均缩短,而萌发历期两者均呈先降低后升高趋势,最短历期均出现在20 ℃,且与其它温度下的萌发历期差异显著(P<0.05)(表3)。15 ℃下,CH种子萌发历期极显著短于CL种子(P<0.01),其它温度下两者无显著性差异(P>0.05)(表3)。
2.3 光照对CH和CL种子萌发特性的影响
种子类型和光照条件对紫花地丁种子的萌发率有显著的交互作用(P<0.05),CH和CL种子的萌发率差异极显著(P<0.01),表明两种类型种子的萌发特性显著不同(表4)。CH种子的萌发率、萌发势和萌发指数在光照条件下均极显著高于黑暗条件下(P<0.01),而CL种子的3个萌发指标在两种条件下均无显著差异(P>0.05)(图1)。虽然紫花地丁的萌发势、萌发率和萌发指数在12 h光照/12 h黑暗条件下CH和CL种子间无显著差异(P>0.05),但24 h黑暗条件下CL种子的各萌发指标极显著高于CH种子的(P<0.01)(图1)。
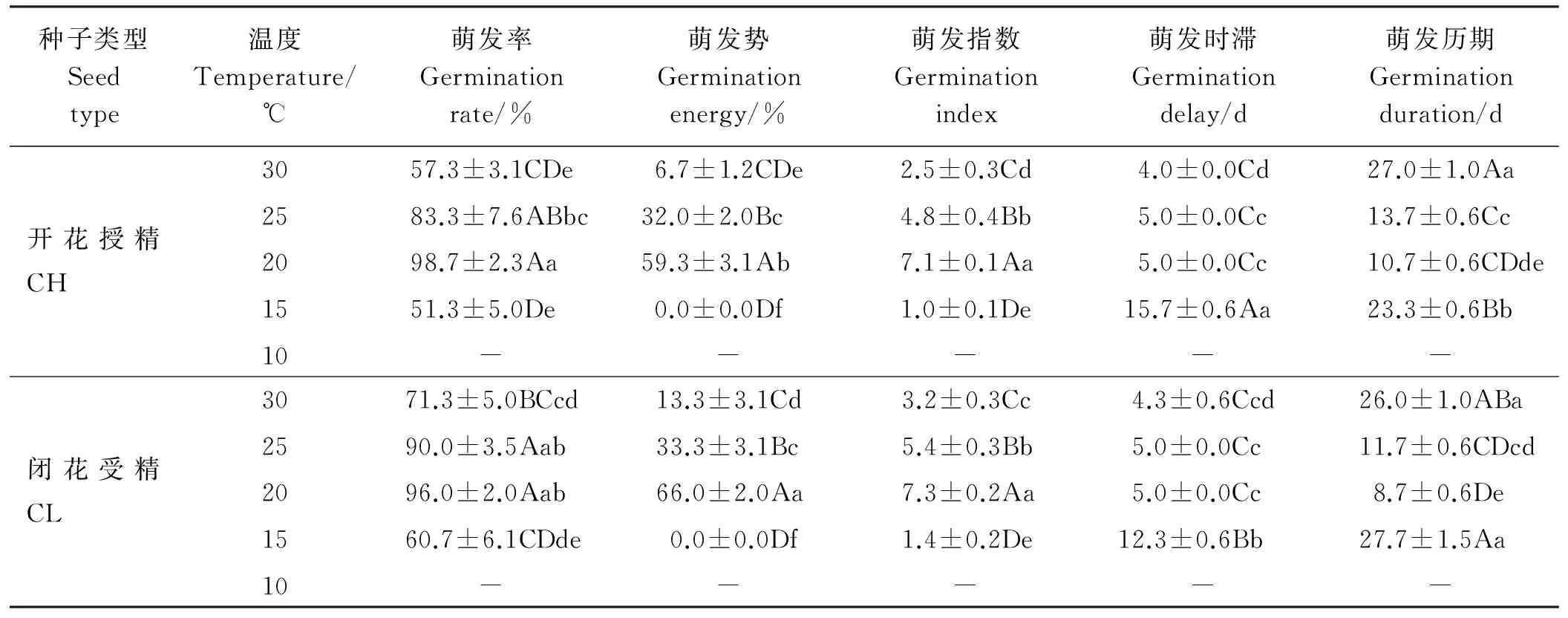
表3 紫花地丁两种类型种子在不同温度条件下萌发指标Table 3 Germination indicators of two types of seeds of V. philippica under different temperature conditions
注:同列不同大、小写字母分别表示差异极显著(P<0.01)和显著(P<0.05)。
Note: Different capital and lower case letters within the same column indicate significant difference at 0.01 and 0.05 level, respectively.
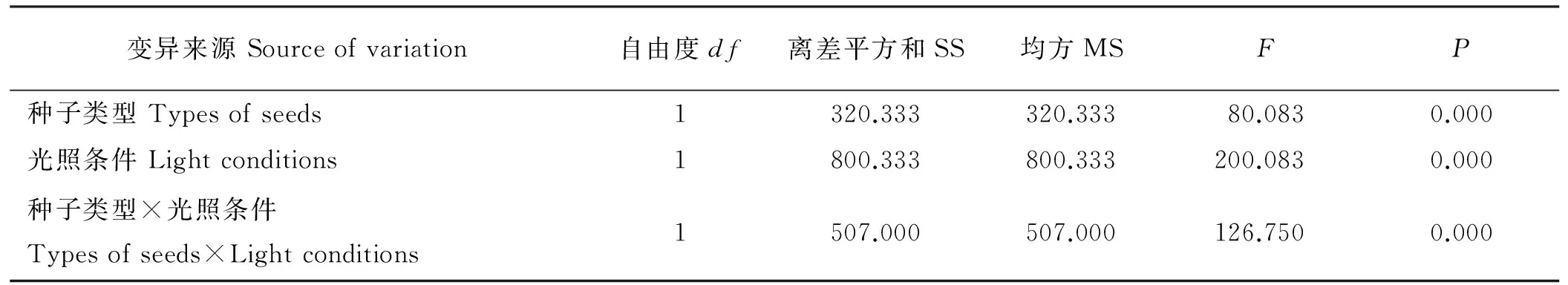
表4 光照条件和种子类型对紫花地丁种子萌发率的影响Table 4 Analysis of variance for effects of light conditions, types of seeds and their interaction on the germination rates of seeds of V. philippica
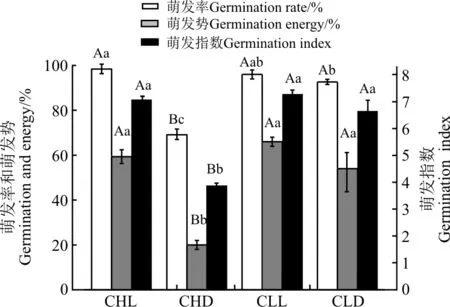
图1 不同光照条件对紫花地丁CH和CL种子萌发率的影响Fig.1 Effect of different light conditions on germination percentages of CH and CL seeds of V. philippica
注:CHL和CHD表示CH种子光照和黑暗;CLL和CLD表示CL种子光照和黑暗;大、小写字表示同一指标不同光照条件下差异显著(P<0.05)。
Note:CHL, CH Cight; CHD, CH dark; CLL, CL Cight; CLD, CL dark; capital and lower case letters indicate signifcant difference among different light conditions with same index at 0.05 level.
2.4 贮藏时间对CH和CL两种类型种子萌发的影响
种子类型和贮藏时间对紫花地丁种子的萌发率有显著的交互作用(P<0.05),CH种子和CL种子的萌发率差异极显著(P<0.01),表明两种类型种子的萌发特性明显不同(表5)。在20 ℃、12 h光照/12 h黑暗条件下,贮藏90 d的CH种子萌发率、萌发势和萌发指数与新采收的种子(对照)相比无显著差异(P>0.05),但此后随着贮藏时间的延长,各萌发指标均显著降低(P<0.05)(图2A)。贮藏210 d,CL种子萌发势和萌发指数均显著降低(P<0.05),但贮藏90、210、360d的CL种子,其萌发率与对照相比没有显著差异(P>0.05),均在90.0%以上(图2B)。与对照相比,CH种子贮藏210 d后,萌发率极显著降低(P<0.01),而CL种子则在贮藏510 d后萌发率才极显著下降(P<0.01)。
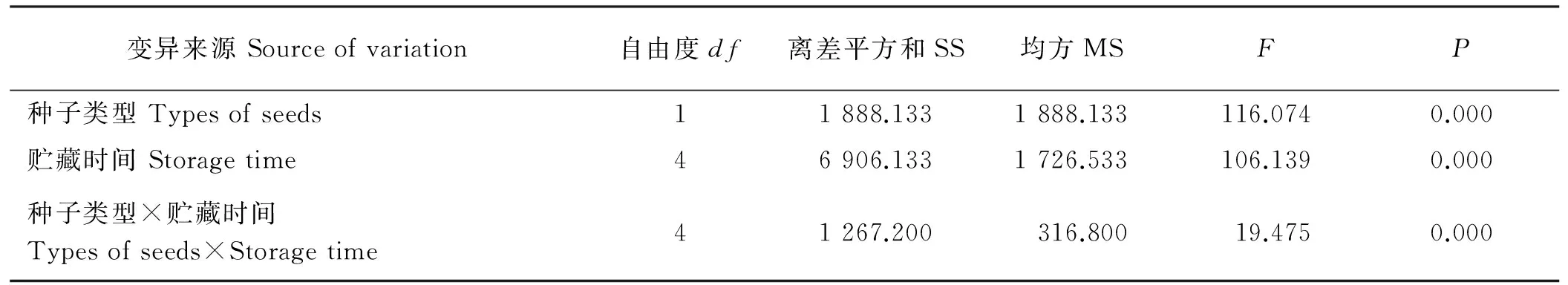
表5 储藏时间和种子类型对紫花地丁种子萌发率的影响Table 5 Analysis of variance for effects of storage time, types of seeds and their interaction on the germination rates of seeds of V. philippica

图2 贮藏时间对紫花地丁CH和CL种子萌发率的影响Fig.2 Effect of storage time on germination percentages of CH and CL seeds of V. philippica
3 讨论与结论
生活在多变环境中的植物,具有异型性种子的物种比只有单型种子的物种有更高的适合度,这是植物为抵御不良的生物和非生物环境在种子阶段所采取的重要策略[6,11]。种子异型性在避免密集负效应、减弱同胞子代间的竞争时采取两面下注(bet-hedging)策略以适应时空异质性等,具有重要的进化生态意义[3]。而报道的大部分异型性种子为同一繁育系统产生的不同位置、不同颜色以及不同大小的种子[3,14],采取混合繁育系统的植物所产生的二型性种子与传统的形态学上的二型性种子不同,他们一般会产生两种遗传组成的种子。自交使种子具有两套完全相同的基因,自交除了可提供繁殖保障,还可阻止对当地适应性基因合成的破坏,保持当地的种群优势;异交可产生遗传多样性的种子,促进了后代适应新环境的能力[24-25]。
同一植株可形成CH花,也可形成CL花,这一混合繁育系统是堇菜属等类群植物的一个著名的现象[26-28],这使得植株可产生两种类型的种子。Berg和Redbo torstensson[29]的研究发现,堇菜属植物V.hirta的CH种子质量大于CL种子,在同样具有此类繁殖策略的凤仙花科植物Impatiensbiflora中也发现了同样的现象[30],即CH种子重量大于CL种子。紫花地丁是常见于北半球温带地区的多年生草本,早春产生CH花,CL花在CH花之后的季节形成[15-16]。本研究发现,其CH和CL种子虽然于色泽、形状上无明显差异,但CH种子长度和宽度以及平均千粒重显著大于CL种子,可见紫花地丁种子在大小上也存在二型性。二型性种子产生的幼苗常存在显著差异,大种子产生的幼苗一般较大[31]。有研究发现,三角叶滨藜(Atriplextriangularis)大种子形成的幼苗甚至在以后的生长中一直保持这种优势[32]。大种子产生的较大个体的幼苗对光照和营养等资源具有较强的竞争能力,也增强了对不利环境条件的抵抗忍耐能力,对于幼苗及植被的更新具有更大的贡献[12,33-35]。相较之下,小种子则具有较强的传播、散布和拓殖优势[35]。紫花地丁形成的不同大小的CH和CL种子可能是其对时空异质性环境采取的适应性策略。
紫花地丁CH和CL两种种子的二型性不仅表现在种子大小上,在萌发特性上也存在明显的差异。研究表明,虽然CH和CL种子均不存在休眠、具有较广的萌发温度范围以及在适宜温度条件下迅速萌发等特点,但两种类型的种子在萌发特性上具有显著差异:CH种子不耐贮藏,在贮藏210 d后萌发率便极显著降低,且黑暗条件对其萌发有极显著的抑制作用,而CL种子在可萌发温度下均具有较高萌发率(>60.0%),贮藏360 d时其萌发率也仍未显著下降,其萌发对黑暗条件不敏感。通过两年观察发现,紫花地丁CH种子成熟期为4月中旬至5月末,兰州地区近几年4月平均温度和5月平均温度分别接近20和25 ℃,而CH种子在这两个温度下萌发率最高,萌发历期最短。CL种子在CH种子之后从夏季一直持续到秋末很长的时期内形成,研究结果显示,CL种子在高温(30 ℃)下萌发时滞最短,萌发率、萌发势和萌发指数均显著高于CH种子,这可能是其对成熟期所处高温环境的长期适应的结果。兰州地区每年的11月至次年2月份日平均温度基本低于10 ℃,种子不萌发以短暂土壤种子库的形式越冬[36],避免了种子萌发而幼苗不能越冬造成的生殖浪费。
上述两种种子的萌发特点表明,与CL种子相比,质量较大、不耐贮藏的CH种子于春季成熟后绝大部分即可迅速萌发,有利于其占据可用空间、利用有限资源[37-39],是对当年种群的有效补充,种子需光萌发的特点也有利于其在春季植被覆盖较低环境下的萌发。一部分CL种子在当年成熟后即可萌发,可对CH种子不足进行补充。CL种子不但在CH种子生产不足时提供了繁殖保障,从而提高了种群整体适合度[40],此外,通过观测发现其种子具有油质体、耐贮藏、萌发不需光等特性,使一部分CL种子可通过蚁运作用降低存在于地表被捕食和损坏的风险[41],成为越冬种子库的主要来源,为来年种群的更新以及扩散占领新生境提供了基础。可见,紫花地丁通过CH和CL花产生在大小、萌发特性不同的CH和CL二型性种子,不仅在传粉者不足和花粉限制时提供了繁殖保障[15,26,30,39],同时也拓宽了种子萌发的时间,以多重繁殖保障机制来适应时空异质性环境,保证了种群的成功扩散、定居和繁衍。
References:
[1] 何芳兰,赵明,王继和,尉秋实,张锦春.几种荒漠植物种子萌发对干旱胁迫的响应及其抗旱性评价研究.干旱区地理,2011,34(1):100-105.
He F L,Zhao M,Wang J H,Yu Q S,Zhang J C.Response to droughty stresses and drought-resistances evaluation of seed germination of four desert vegetation.Arid Land Geogarphy,2011,34(1):100-105.(in Chinese)
[2] 张景宽,张文辉,张洁明,刘宝玉,刘新成.种子萌发期4种植物对干旱胁迫的响应及其抗旱性评价研究.西北植物学报,2006,26(9):1811-1818.
Zhang J K,Zhang W H,Zhang J M,Liu B Y,Liu X C.Response to droughty stresses and drought resistances evaluation of four species during seed germination.Acta Botanica Boreal-Occidentalia Sinica,2006,26(9):1811-1818.(in Chinese)
[3] 马文宝,谭敦炎,薛建辉,刘兴良.果实(种子)多态性及生态学意义.四川林业科技,2011,32(6):34-43.
Ma W B,Tan D Y,Xue J H,Liu X L.Seed heteromorphism and its ecological significance.Journal of Sichuan Forestry Science and Technology,2011,32(6):34-43.(in Chinese)
[4] Rajjou L,Duval M,Gallardo K,Catusse J,Bally J,Job C,Job D.Seed germination and vigor.Annual Review of Plant Biology,2012,63:507-533.
[5] Imert E.Ecological consequences and ontogeny of seed herteromorphism.Perspectives in Plant Ecology,Evolution and Systematic,2002,5(1):13-36.
[6] Venable D L.The evolutionary ecology of seed heteromophism.American Naturalist,1985,126(5):577-595.
[7] Maun M A,Payne A M.Fruit and seed polymorphism and its relation to seedling growth in the genusCakile.Canadian Journal of Botany,1989,67:2743-2750.
[8] 刘鹏伟,魏岩.准噶尔荒漠异苞滨藜(Atriplexmcrantha)的种子二型性及其萌发行为.生态学报,2007,27(10):4233-4239.
Liu P W,Wei Y.Seed dimorphism and germination behavior ofAtriplexmicrantha,an annual inhabiting Junggar desert.Acta Ecologica Sinica,2007,27(10):4233-4239.(in Chinese)
[9] 高蕊,魏岩,严成.角果藜的地上地下结果性与种子萌发行为.生态学杂志,2008,27(1):23-27.
Gao R,Wei Y,Yan C.Amphicarpy and seed germination behavior ofCeratocarpusarenariusL.(Chenopodiaceae).Chinese Journal of Ecology,2008,27(1):23-27.(in Chinese)
[10] 刘艳芳,魏岩,严成.异子蓬二型种子的萌发与休眠特性及其生态适应.生态学报,2009,29(12):6609-6614.
Liu Y F,Wei Y,Yan C.Germination characteristics and ecological adaptation of dimorphic seeds ofBorszczowiaaralocaspica.Acta Ecologica Sinica,2009,29(12):6609-6614.(in Chinese)
[11] 吴泽昂,李予霞,庄丽,王海霞.异型狭果鹤虱的二型性种子萌发特性及其生态适应.新疆农业科学,2013,50(20):307-312.
Wu Z A,Li Y X,Zhuang L,Wang H X.Germination characteristics and ecological adaptation of dimiorphic seeds ofL.Semiglabra.Xinjiang Agricultural Sciences,2013,50(20):307-312.(in Chinese)
[12] 刘艳,周家超,张晓东,李欣,范海,王宝山,宋杰.盐地碱蓬二型性种子及其幼苗对盐渍环境的适应性.生态学报,2013,33(17):5162-5168.
Liu Y,Zhou J C,Zhang X D,Li X,Fan H,Wang B S,Song J.Adaptations of dimorphic seeds and seedlings ofSuaedasalsato saline environments.Acta Ecologica Sinica,2013,33(17):5162-5168.(in Chinese)
[13] 杨帆,曹德昌,杨学军,高瑞如,黄振英.盐生植物角果碱蓬种子二型性对环境的适应策略.植物生态学报,2012,36(8):781-790.
Yang F,Cao D C,Yang X J,Gao R R,Huang Z Y.Adaptive strategies of dimorphic seeds of the desert halophyteSuaedacorniculatain saline habitat.Chinese Journal of Plant Ecology,2012,36(8):781-790.(in Chinese)
[14] 王雷,董鸣,黄振英.种子异型性及其生态适应意义的研究进展.植物生态学报,2010,34(5):578-590.
Wang L,Dong M,Huang Z Y.Review of research on seed heteromorphism and its ecological significance.Chinese Journal of Plant Ecology,2010,34(5):578-590.(in Chinese)
[15] 刘绮丽,刘香梅,刘薇薇,廖万金.紫花地丁开放花和闭锁花繁殖特征的研究.北京师范大学学报:自然科学版,2006,42(6):605-609.
Liu Q L,Liu X M,Liu W W,Liao W J.Comparison between cleistogamy and chasmogamy in perennial herbViolayedoensis(Violaceae).Journal of Beijing Normal University:Natural Science Editione,2006,42(6):605-609.(in Chinese)
[16] 中国科学院植物研究所植物志编委会.中国植物志.北京:科学出版社,1991:63.
Editorial Committee of Chinese Journal of Plant of Chinese Academy of Sciences.Flora of China.Beijing:Science Press,1991:63.(in Chinese)
[17] 黄曼,王东.油质体在5种蚁播植物种子散布中的作用.生态学报,2015,35(17):5721-5727.
Huang M,Wang D.The role of elaiosome in seed dispersal of five ant-dispersed herbaceous plants.Acta Ecologica Sinica,2015,35(17):5721-5727.(in Chinese)
[18] 巩明明,苏雪,孙坤.青藏高原特有植物块茎堇菜地上地下结果性及散布机制.草业科学,2009,26(4):16-19.
Gong M M,Su X,Sun K.Amphicarpy and dispersal mechanism ofViolatuberiferaendemic to Qinghai-Tibetan Plateau.Pratacultural Science,2009,26(4):16-19.(in Chinese)
[19] 苏秋霞,李青丰.紫花地丁种子萌发检验标准化.种子,2014,33(10):113-116.
Su Q X,Li Q F.Standardization for seed germination testing ofViolaphilippicaCav.Seed,2014,33(10):113-116.(in Chinese)
[20] 罗铮,孙群,曲超,王永春,孙宝启,丁自勉.紫花地丁种子最适采收期及其贮藏方法研究.种子,2007,26(7):60-62.
Luo Z,Sun Q,Qu C,Wang Y C,Sun B Q,Ding Z M.Study on the optimal picking time and storage method ofViolayedoensisMakino seeds.Seed,26(7):60-62.(in Chinese)
[21] 刘明久,许桂芳,王鸿升.四种野生地被植物资源及种子特性研究.种子,2007,26(12):47-49.
Liu M J,Xu G F,Wang H S.The resources and seed germination chrachteristics of four wild ground cover plants.Seed,2007,26(12):47-49.(in Chinese)
[22] 董慧,王艳芳,刘群龙,牛铁荃,张建成,郝燕燕.紫花地丁种子萌发特性研究.山西农业科学,2015,43(7):831-833.
Dong H,Wang Y F,Niu Q L,Liu T Q,Zhang J C,Hao Y Y.Study on seed germination characteristics ofViolayeoensisMakino.Journal of Shanxi Agricultural Sciences,2015,43(7):831-833.(in Chinese)
[23] 王非,王金侠,李强,何淼.GA3和IAA处理对4种铁线莲种子萌发的影响.草业科学,2014,31(4):672-676.
Wang F,Wang J X,Li Q,He M.Effects of GA3and IAA on the germination of four species ofClematiseseeds.Pratacultural Science,2014,31(4):672-676.(in Chinese)
[24] Charlesworth D.Evolution of plant breeding systems.Current Biology,2006,16:726-735.
[25] 何亚平,刘建全.植物繁育系统研究的最新进展和评述.植物生态学报,2003,27(2):151-163.
He Y P,Liu J Q.A review on recent advances in the studies of plant breeding system.Acta Phytoecologica Sinica,2003,27(2):151-163.(in Chinese)
[26] Forrest J,Thomson J D.Pollen limitation and cleistogamy in subalpineViolapraemorsa.Botany,2008,86:511-519.
[27] Cortes-Palomec A C,BallardH E.Influence of annual fluctuations in environmental conditions in chasmogamous flower production inViolastriata.Journal of the Torrey Botanical Society,2006,133(2):312-320.
[28] 郝楠,苏雪,吴琼,常立博,张世虎,孙坤.祁连山高寒草甸两型闭锁花植物块茎堇菜的繁殖分配.草业科学,2015,32(4):586-592.
Hao N,Su X,Wu Q,Chang L B,Zhang S H,Sun K.Reproductive allocation of dimorphic cleistogamousViolatuberiferain alpine meadow of Qilian Mountain.Pratacultural Science,2015,32(4):586-592.(in Chinese)
[29] Berg H,Redbo-Torstensson P.Offspring performance in three cleistogamousViolaspecies.Plant Ecology,1999,145:49-58.
[30] Schemske D W.Evolution of reproductive characteristics inImpatiens(Balsaminaceae):The significance of cleistogamy and chasmogamy.Ecology,1978,59:596-613.
[31] 李伟强,刘小京,毛任钊,安萍,乔海龙,黄玮,李志刚.植物种子二型性(多型性)研究进展.生态学报,2006,26(4):1234-1241.
Li W Q,Liu X J,Mao R Z,An P,Qiao H L,Huang W,Li Z G.Advances in plant seed dimorphism (or polymorphism) research.Acta Ecologica Sinica,2006,26(4)):1234-1241.(in Chinese)
[32] Ellison A M.Effect of seed dimorphism on the density-dependent dynamics of experimental populations ofAtriplextriangularis(Chenopodiaceae).American Journal of Botany,1987,74(8):1280-1288.
[33] Gross K L.Effects of seed size and growth form on seeding establishment of six monocarpic plants. Journal of Ecology,1984,72(2):369-387.
[34] 赵可夫,范海,江行玉,宋杰.盐生植物在盐渍土壤改良中的作用.应用与环境生物学报,2002,8(1):31-35.
Zhao K F,Fan H,Jiang X Y,Song J.Improvement and utilization of saline soil by planting halophytes.Chinese Journal Environmental Biology,2002,8(1):31-35.(in Chinese)
[35] 武高林,杜国祯.植物种子大小与幼苗生长策略研究进展.应用生态学报,2008,19(1):191-197.
Wu G L,Du G Z.Relationships between seed size and seedling growth strategy of herbaceous plant a review.Chinese Journal of Applied Ecology,2008,19(1):191-197.(in Chinese)
[36] 于顺利,陈宏伟,郎南军.土壤种子库的分类系统和种子在土壤中的持久性.生态学报,2007,27(5):2099-2108.
Yu S L,Chen H W,Lang N J.The classification systems of soil seed banks and seed persietence in soil.Acta Ecologica Sinica,2007,27(5):2099-2108.(in Chinese)
[37] Ross M A,Harper J L.Occupation of biological space during seedling establishment.Journal of Ecology,1972,60:77-88.
[38] 吴冬,黄姝博,李宏庆.意大利苍耳二型性种子萌发、植株生长差异及生态适应性.生态学报,2009,29(10):5258-5263.
Wu D,Huang S B,Li H Q.Dimorphic seed germination,plant growth difference,and ecological adaptability ofXanthiumitaficum.Acta Ecologica Sinica,2009,29(10):5258-5264.(in Chinese)
[39] 李雪华,蒋德明,刘志民,李晓兰.温带半干旱地区一年生植物种子的萌发特性.生态学报,2006,26(4):1194-1199.
Li X H,Jiang D M,Liu Z M,Li X L.Seed germination characteristics of annual species in temperate semi-arid region.Acta Ecologica Sinica,2006,26(4):1194-1199.(in Chinese)
[40] Waller D M.The relative costs of self- and cross- fertilized seeds inImpatienscapensis(Balsaminaceae).American Journal of Botany,1979,66:313-320.
[41] 祝艳,王东.蚂蚁觅食与搬运行为对阜平黄堇和小花黄堇种子散布的影响.生态学报,2014,34(17):4938-4942.
Zhu Y,Wang D.Seed dispersal ofCorydaliswilfordiiandCracemosa(Papaveraceae):Effect of ant foraging and behavior and seed characteristics.Acta Ecologica Sinica,2014,34(17):4938-4942.(in Chinese)
(责任编辑 苟燕妮)
Germination characteristics and ecological significance of dimorphic seeds ofViolaphilippica
Su Xue1, Gao Ting1, Hou Yun-yun1,2, Wu Qiong1, Sun Kun1
(1.College of Life Science, Northwest Normal University, Lanzhou 730070, China;2.Fuping High School, Weinan 711700, China)
Violaphilippicahas dimorphic seeds which come from chasmogamous flowers and cleistogamous flowers. Based on the morphology observation of CH and CL seed, we compared the germination characteristics of the two types of seeds at different temperatures, light conditions and the storage time. The results showed that: 1) The length, width and weight of CL seeds were significantly lighter (P<0.05) than CH seeds. 2) The optimum germination temperatures of CH and CL seeds were 20 ℃, and both the germination rates were above 95%. Both of the two types of seeds did not germinate at a low temperature (10 ℃), but germination rate of CH seeds was significantly lower (P<0.05) than CL seeds at a high temperature (30 ℃). As to the moderate temperature (15-25 ℃), the germination rates of CH and CL seeds did not differ significant(P>0.05). 3) CH seeds were sensitive to light and its germination rate would be inhibited in the dark (P<0.01), but the germination rate of CL seeds had no significantly difference at 12 h light and full dark conditions(P>0.05). 4) Compared with the control, the germination rates of CH and CL seeds significantly decreased after 210 d and 510 d storage (P<0.05), respectively. CH seeds maturing in the spring can quickly germinate. CL seeds germinated later in the season than CH seeds. The part of CL seeds could germinate, another part became transient soil seed bank and could germinate next spring. The differences in germination characteristics between dimorphic seeds ofV.philippicaimproved the species’ fitness to heterogeneous habitats.
Violaphilippica; seed dimorphism; germination characteristic; ecological adaptation
Su Xue E-mail:suxue@nwnu.edu.cn
10.11829/j.issn.1001-0629.2016-0036
2016-01-16 接受日期:2016-06-14
国家自然科学基金项目(31260054)
苏雪(1974-),女,辽宁辽阳人,副教授,硕士,主要从事植物繁殖生态学的研究。E-mail:suxue@nwnu.edu.cn孙坤(1965-),男,甘肃民勤人,教授,博士,主要从事植物系统进化和生物多样性等研究。E-mail:kunsun@nwnu.edu.cn
S314;Q945.34
A
1001-0629(2016)10-2025-08*
Sun Kun E-mail:kunsun@nwnu.edu.cn
苏雪,高婷,侯云云,吴琼,孙坤.紫花地丁二型性种子的萌发特性及其生态意义.草业科学,2016,33(10):2025-2032.
Su X,Gao T,Hou Y Y,Wu Q,Sun K.Germination characteristics and ecological significance of dimorphic seeds ofViolaphilippica.Pratacultural Science,2016,33(10):2025-2032.
