浙江低山地区多用途植物无患子的开花物候特征
2016-11-28刁松锋姜景民岳华峰董汝湘孙洪刚邵文豪
刁松锋,姜景民,伊 焕,岳华峰,董汝湘,孙洪刚,邵文豪,*
1 国家林业局泡桐研究开发中心,郑州 450003 2 中国林业科学研究院亚热带林业研究所,杭州 311400 3 中国林业科学研究院经济林研究开发中心,郑州 450003 4 河南省梁园区林业局,商丘 476000
浙江低山地区多用途植物无患子的开花物候特征
刁松锋1,2,3,姜景民2,伊 焕4,岳华峰1,董汝湘2,孙洪刚2,邵文豪2,*
1 国家林业局泡桐研究开发中心,郑州 450003 2 中国林业科学研究院亚热带林业研究所,杭州 311400 3 中国林业科学研究院经济林研究开发中心,郑州 450003 4 河南省梁园区林业局,商丘 476000
无患子(SapindusmukorossiGaertn.)是我国长江以南地区传统的重要绿化树种,其果皮富含皂苷,种仁富含油脂,是国家林业局审定的新型木本油料树种之一。为研究多用途树种无患子在浙江低山地区的开花特征,2012年和2013年连续两年对位于浙江省天台县9年生无患子人工林在群体、个体、花序和单花水平进行开花物候观测和比较,并运用开花日期、相对开花强度和同步性等指数研究了无患子开花物候特征。观察结果显示:无患子花呈浅黄白色,花的类型有雄花和两性花,没有发现雌花。雄花较两性花大(花径分别为5.09 mm和3.72 mm),雄蕊多为8枚,个别7或9、10枚;雌蕊退化仅留下浅绿色凸起。两性花花萼稍抱拢,花药藏于花被片下,雄蕊大多8枚,极少数7枚或9枚,柱头高于花药并伸出花苞,子房一般具3室,极少数4室或仅2室。无患子2012年的开花进程略早于2013年,其花期集中在在5月中旬至6月上旬,单花从花蕾膨大到花朵凋谢一般为8—9 d。在2012年和2013年,无患子在群体、个体和花序水平的花期约为30 d、20 d、11 d和28 d、19 d、13 d。个体水平的开花振幅均呈单峰曲线,年际间相似性较高;开花同步性在个体水平同步性较高(同步指数为0.868),表现出一种大量、集中的开花式样;相对开花强度在单株间分布范围相对宽泛,但主要分布在30%—40%,在年际间和年际内均呈现极显著差异。花期同步指数在两年的变异范围分别是0.81—0.97和0.70—0.98,不同单株开花同步性在年际内差异极显著,但在年际间差异性则不显著。由此可见,无患子的生殖资源分配存在明显的时空差异,较长的花期可以减少非法花粉的干扰、保持其种群基因多样性,遗传因子是决定无患子开花物候的主要因素,生态环境对无患子开花物候的影响还需进一步研究,本研究以期为探索无患子开花的主要限制因子奠定基础。
无患子;花序生长;开花物候;相对开花强度;同步性指数
无患子(SapindusmukorossiGaertn.)为无患子科(Sapindaceae)无患子属(Sapindus)落叶乔木,具大型圆锥花序,是江南地区重要的绿化彩化树种,分布于东亚至南亚多个国家,中国分布于秦岭——淮河以南低山丘陵地区[1]。无患子果皮中的皂苷含量可达10.76%[2],是一种天然的非离子型表面活性剂,具有良好的起泡性和去污能力,可用来替代石化产品原料生产洗涤剂[3];另外,无患子皂苷还具有抗病毒、降血压等药理作用[4- 5]。无患子种仁油脂含量可达42.73%,其中不饱和脂肪酸含量高达86.63%[6],具有开发生物柴油的潜力[7]。2012年,无患子被国家林业局列为重要生物质能源树种,并发布了其原料林培育技术规程。可以预见,无患子可望成为支撑新兴绿色产业的新型经济林树种。
植物开花物候不仅能反映植物开花式样和非生物因子间的关系,而且可以反映植物开花的遗传基础和自然选择的关系[8],又能表征开花植物群体的适合度[9- 11]。植物的开花物候受遗传和环境因子共同影响,在群体、个体、花序和单花水平上具有不同的差异性[12],不同的物候特征对植物的生殖成功有重要影响[13]。植物花在一个季节不同时间开放,会改变花产生果实和种子的量[14],开化进程不同会影响果实和种子的总产量[15]。目前,国内外学者对无患子的研究大多集中在其皂苷化学活性成分检测[16- 17]及提取工艺[18]、药理效果与临床应用[4- 5]、种实表型变异[19- 20]、经济性状区域差异[2,21]、果实发育及其生理和光合变化[22- 23]等方面,而关于无患子开花物候还未见报道。因此,本试验以9年生无患子人工林为研究对象,连续两年从以上4个水平研究无患子开花物候特征及各物候参数的时空变异程度,研究无患子开花物候特征及其受环境因子影响的程度,探索无患子群体开花物候和面对外界逆境时的生殖适应策略,以期为研究无患子繁育系统及杂交育种提供理论依据,为无患子果用林高产、丰产奠定基础。
1 材料与方法
1.1 试验地概况
试验林地位于浙江省天台县,28°57′02″N、120°41′24″E,海拔55—95 m。该试验林为采自当地无患子老树种子育苗营建的、林龄9年的人工实生林,面积约100 hm2,株行距4 m×4 m,林分郁闭度约为75%,树高平均为4.82 m,胸径平均为8.67 cm。基地年平均气温16.8 ℃,降雨量1320 mm,1月份和7月份平均温度分别5.1 ℃和28.5 ℃,年无霜期平均232 d。地形海拔差异不大、地势较为平坦、林分立地条件基本一致,林地属黏性红黄壤,土层厚度20—70 cm,pH约6.0。林下散生着枸骨(Ilexcornuta)、络石(Trachelospermumjasminoides)、小果蔷薇(Rosacymosa)、铁芒萁(Dicranopterislinearis)、山莓(Rubuscorchorifolius)等植被。根据天台县气象资料记载:本县2012年5月1日—2012年6月30日和2013年5月1日—2013年6月30日的平均气温为26.5 ℃和22.7 ℃,最高温度34.4 ℃和32.1 ℃,最低温度20.3 ℃和16.5 ℃,两个月内的降雨量分别为368 mm和354 mm。
1.2 试验方法
参照Dafni[24]和马文宝[11]等方法,在林内标记长势良好、大小基本一致的30棵单株,并在每单株的中上部、阳面标记3枝发育正常的花序,分别于2012年和2013年对无患子开花物候在群体、单株、花序和单花4个水平上进行了观察,记录花序和植株上每天开花的数量。
1.2.1 单花水平
在每单株已确定的3个花序上分别标记30个花蕾,调查花蕾形成、至迅速膨大、至始放的时间,以及完全萎蔫的时间,记录花部各器官形态变化。调查花序不同空间部位开花次序及持续时间,统计花的寿命。并用解剖镜和电子显微镜观测花各部分形态特征,记录花径长度(FD, mm)、花药长度(AL, mm)、花药宽度(AW, mm)、花丝长度(FL, mm)、柱头长度(SH, mm),统计雄蕊数量(ST, n)、子房室数(CO, n)和萼片数(SN, n)。
1.2.2 群体、单株和花序水平
花序和单株水平的开花物候参数以所标记的全部花序和单株的平均值计算。观察和统计的开花期参数主要有始花期(FFS)、开花高峰期(PFD)、持续时间(duration)和末花期(LFD)。花序和单株的始花期为第1朵花在花序开花的日期,花序和单株开花数大于或等于50%时进入开花高峰期,基本再无新花开放后进入末花期;群体水平(以样地为标准)以25%的个体开花时则进入始花期,50%的个体达到开花高峰时则群体进入开花高峰期,95%的植株开花结束时则为群体的终花期。始花期的统计参考Pickering[25]的方法。在本研究中,2012年以5月16日为第1天(计为1),5月17日为第2天(计为2);同理,2013年以5月21日计为1,5月22日计为2,依此类推。

1.3 数据处理
采用EXCEL 2010软件进行数据统计和作图,采用SPSS 20.0软件进行数据统计和方差分析。
2 结果与分析
2.1 无患子花类型及其特征
无患子花呈浅黄白色,花的类型有雄花和两性花,没有发现雌花。雄花较两性花大,由花萼、花瓣和雄蕊组成,花萼5深裂,裂片卵形或长圆卵形,花瓣5,多披针形,雄蕊多为8枚,个别7或9、10枚;花丝呈放射状散开,下部多绒毛;花药呈黄色,纵裂;雌蕊退化仅留下浅绿色凸起(图1)。两性花花萼稍抱拢,花丝较短,多被绒毛围绕,花药藏于花被片下,雄蕊大多8枚,极少数7枚或9枚,柱头高于花药并伸出花苞,子房一般具3室,极少数4室或仅2室,未发现1室的子房(图1,图2)。表1为无患子两种类型花的形态结构数量特征,由此可知无患子雄花的花径、花药长度和花丝长度均比两性花大,其中花丝长度约是两性花的2.31倍,而雄蕊数和花药宽度则相差不大。

图1 无患子花类型Fig.1 The type of flower of S. mukoross

图2 无患子两性花中雄蕊数和心室数差异Fig.2 The different number of stamen and chamber of bisexual flower of S. mukorossi

花类别Flowertype数值类型Valuetype花径FD/mm雄蕊ST/枚花药长度AL/mm花药宽度AW/mm花丝长度FL/mm柱头高度SH/mm子房/室CO两性花Bisexualflower最大值3.9490.830.511.836.0173最小值3.4470.650.371.574.3222均值 3.7280.720.441.695.0243雄花Maleflower最大值5.37100.940.514.52——最小值4.4270.810.423.78——均值 5.0980.870.453.91——
FD:花径长度 flower diameter;AL:花药长度 anther length;AW:花药宽度 anther width;FL:花丝长度filamen length;SH:柱头长度 stigma height;ST:雄蕊数量 stamen number;CO:子房室数chamber of an ovary;SN:萼片数 sepal number
2.2 无患子单花发育及形态变化

图3 无患子单花发育不同阶段表型特征Fig.3 The development and it′s phenotypic characteristic of single flower of S. mukorossi
无患子花一般是由花序中部向底部,然后向顶部依次开放。单花从花蕾膨大到花朵凋谢一般为8—9 d。单花物候依据其表型变化可分为5个时期,即:盛蕾期、花前期、初开期、全开期、凋萎期(表2)。
在开花过程中,花丝和花柱都有伸长,但其生长速率不同,空间位置发生动态变化,这种位置的变化是随着单花物候进展而进行的(图3)。在盛蕾期,花苞为绿色,萼片紧闭,花柱和花丝均未露出花苞,花柱被花丝覆盖,此过程约持续2 d。在花前期,萼片微微张开,柱头微露、呈黄色,花丝和花柱近于平行状态,此过程约持续1 d。在初开期,萼片张开,花苞有“O”形状张口,柱头伸出花苞,柱头呈淡黄色,略高于花药,8枚雄蕊中的2—3枚先伸出花苞,并紧贴花柱,此过程在1 d内结束。在全开期,花苞完全张开,柱头完全伸展,呈黄灰色,花药位于柱头中下部,1—2枚花药脱离花丝,此过程约持续2 d。在凋萎期,萼片脱落,花药几乎全部脱落,柱头呈现黑色。调查过程中发现,由于无患子花序花量很大,花败育情况比较普遍,在风雨天气,出现大量花蕾凋落。

表2 无患子单花发育动态及表型变化
2.3 无患子群体、个体和花序水平开花物候
无患子花期为每年的5月中下旬至6月下旬。由表3可知,在2012年和2013年,无患子在群体、单株和花序3个水平的花期分别为30 d和28 d,20 d和19 d,11 d和13 d。其中,2012年无患子在群体水平、单株水平和花序水平的始花期、花期持续时间、开花高峰日期和终花期分别比2013年早4 d、6 d、5 d,2 d、1 d、2 d,3 d、5 d、4 d和3 d、5 d、6 d。2012年的开花物候进程早于2013年,这与2012年气温回升较2013年早有关。并且3个水平的花期持续时间的天数在2012年和2013年的差值小于3个水平在始花期、开花高峰日期和终花期的差值,这表明无患子开花物候期受环境影响较大。在3个水平中,无患子2012年的开花振幅均小于2013年(表3),两年的开花比例曲线具有相同的趋势,均呈单峰曲线,即均表现为急速上升至高峰,然后缓慢下降,最高峰分别出现在开花后的第8天和第6天(图4)。

表3 无患子群体、单株和花序水平的开花物候(2012年和2013年)
2.4 无患子开花物候参数特征
根据表4和图5可以看出,不同单株的相对开花强度变异范围在2012年和2013年分别为14.71%—88.86%和12.38%—95.09%。t检测发现单株间的相对开花强度在年际内和年际间均表现为极显著差异(P<0.001)。花期同步指数在两年的变异范围分别是0.81—0.97和0.70—0.98,但单株间花期同步性仅在年际内具有极显著差异(P<0.01),在年际间差异则不显著(P=0.483>0.05)。2012年和2013年,无患子平均花期同步指数为0.868,表现出一种大量、集中的开花式样。图6显示,无患子单株开花强度在2012年和2013年的主要分布范围均为30%—40%,其分布频度的偏斜率分别为1.75和1.72。
3 结论与讨论
全面、系统地了解植物花序生长和开花物候特性,既是进行植物生活史研究的前提,也是开展种质资源保存、利用及育种等相关研究的基础[26]。无患子多分布在江南低山丘陵地区,浙江省是无患子的自然分布区之一,是无患子人工林栽植面积最大的地区之一,选择江南地区生态环境具有代表意义的天台县的无患子人工林为研究对象,研究结果可为其他区域作为参考。
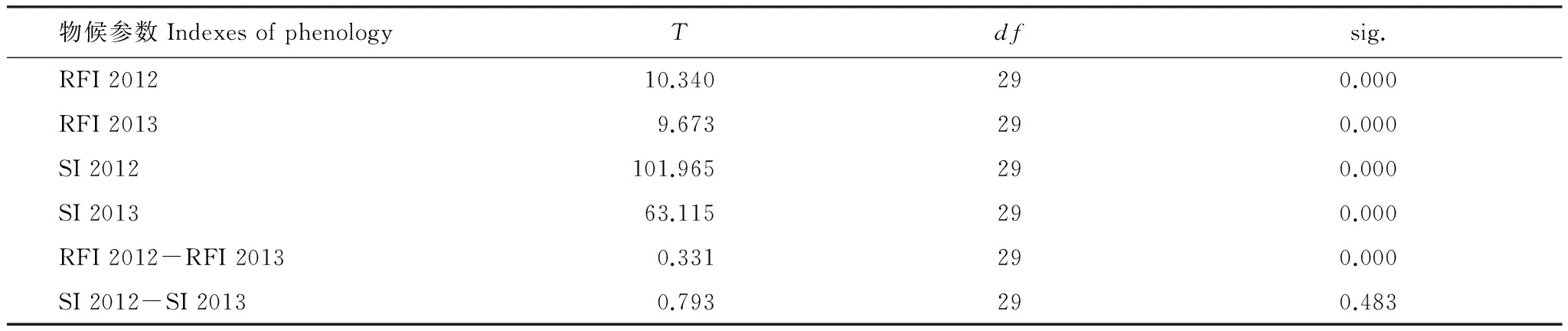
表4 相对开花强度和同步性指数在年度间的t检测
RFI: 相对开花强度relative flowering intensity;SI: 和开花同步指数synchrony index
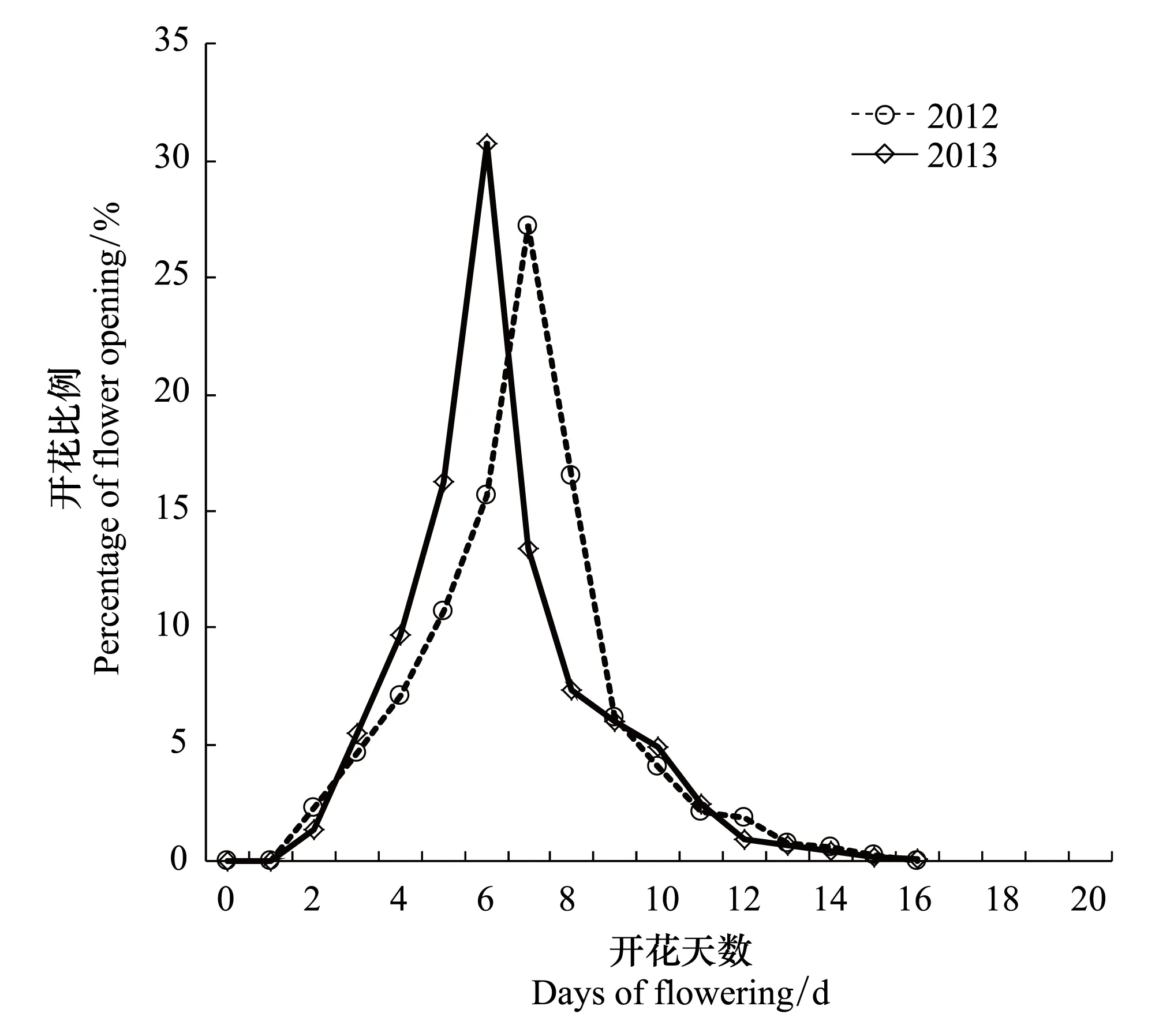
图4 无患子单株开花振幅曲线(2012和2013年) Fig.4 Flowering amplitudes curves of S. mukorossi individual plant (2012 and 2013)

图5 无患子单株相对开花强度和花期同步指数(2012和2013年)Fig.5 The relative flowering intensities and flowering synchrony indices of S. mukorossi (2012 and 2013)

图6 无患子单株相对开花强度分布 Fig.6 Frequency distributions of relative flowering intensity in S. mukorossi individual plants
植物个体物候的总和是一个群体开花物候,不同植物开花持续时间也不同[13]。锥栗(Castaneahenryi)雄花序花期4—5月,单花序花期10—12 d,群体花期23—26 d(其中盛花期13—15 d)[27]。无患子科植物中,无患子科植物中,花序开放顺序多为雄花→雌花/两性花→雄花,其中龙眼(Dimocarpuslongan)人工林花期为3月下旬到5月上旬,群体花期约为50 d[28];文冠果(Xanthocerassorbifolia)人工林花期约24 d,5月初为始花期,5月上旬为盛花期,5月中下旬为开花末期和落花期[29]。不同环境和立地条件开花物候也有差异,北京植物园文冠果比定西地区早一周左右[29- 30];位于福州的于国家龙眼品种资源圃中的龙眼比位于广东省湛江市的龙眼花期短15 d左右[28,31]。本研究发现位于浙江省天台县的无患子人工群体的花期约为50 d。
植物个体的开花物候常在两个极端之间变化,一个极端是物种表现为“集中开花模式”,此模式下,个体每天或一个星期左右产生大量新的花;另一极端是物种具有“持续开花模式”,此模式下,群体中的个体基本上每天或几个星期产生少量新的花[32- 33]。在2012年和2013年连续两年的研究中,无患子个体水平的开花物候进程(开花振幅曲线)表现出较高的相似性,都呈单峰曲线模式,都存在一个开花高峰期,且有很高的开花同步指数0.868,表现出集中开花模式,这种开花模式在很多种植物中都存在[10- 11,32]。无患子为虫媒花植物,集中开花模式是无患子应对江南地区5—6月份多雨多风的的条件,减少气象因素对虫媒传粉频率和效率的影响,提高散粉效率的一种生殖适应性。
相对开花强度是植物花资源空间分布的一个指标[34],它可能会影响到植物花粉的运动模式[23],研究发现多数植物具有较低的相对开花强度[32,34]。无患子也具有较低的相对开花强度,分布频率集中在30%—40%之间;在2012年和2013年,个体开花强度的偏斜率为分别为1.75和1.72。相对开花强度在年际内和年际间均表现为极显著差异,说明无患子相对开花受个体遗传和环境共同决定,而花期同步指数仅在年际内具有极显著差异,在年际间差异则不显著,表明遗传因素较环境因素对无患子花期同步性的影响大,这与刘曲等[35]对分布在浙江低山地区的油茶(Camelliaoleifera)人工林和陈芬等[36]对浙江金华市薄壳山核桃(CaryaillinoensisK. Koch)人工林的开花物候研究结果一致。无患子的开花强度偏斜率略高于Herrera[32]研究26种植物个体开花强度平均偏斜率(1.59)。无患子单株开花强度频率分布偏斜严重,表明在研究群体中具有相对开花强度高的单株。无患子始花日期早的单株比始花日期晚的单株具有更长的花期,而开花数多的单株比开花数少的单株具有更长的花期,这与许多研究结果相吻合[10- 11,34- 37]。Zimmerman等[15]认为花期开花进程影响植物总的雌性适合度,进而影响结实量,这可能是无患子出现“花而不实”、“千花一果”现象的重要原因之一。
植物个体开花物候在很大程度上是由其遗传因子决定的,但开花物候在年度间出现的变异,受年际间气象因素影响较大[15]。根据天台县气象资料可知2012年和2013年同期降水量基本持平,但2013年最高温度、最低温度和平均温度均明显低于2012年的同期水平。温度通常被认为是决定植物体开花的主要因素,对于相同物种积温高有利于更早开花[38],李新蓉等[37]研究表明低温不仅会对单花和花序水平的物候特征产生较大影响,而且也会显著影响个体的开花进程。但无患子各水平开花物候在年际间与环境,特别是积温差的关系还需进行多年的数据积累并结合生态环境因子的模拟研究。
[1] 中国植物志编辑委员会. 中国植物志第四十七卷(第1分册). 北京: 科学出版社, 1985.
[2] 邵文豪, 姜景民, 董汝湘, 岳华峰, 刁松锋. 不同产地无患子果皮皂苷含量的地理变异研究. 植物研究, 2012, 32(5): 627- 631.
[3] Ghagi R, Satpute S K, Chopade B A, Banpurkar A G. Study of functional properties ofSapindusmukorossias a potential bio-surfactant. Indian Journal of Science and Technology, 2011, 4(5): 530- 533.
[4] Ibrahim M, Khaja M N, Aara A, Khan A A, Habeeb M A, Devi Y P, Habibullah C M. Hepatoprotective activity ofSapindusmukorossiandRheumemodiextracts:Invitroandinvivostudies. World Journal of Gastroenterology, 2008, 14(16): 2566- 2571.
[5] Verma N, Amresh G, Sahu P K, Mishra N, Singh A P, Rao C V. Antihyperglycemic activity, antihyperlipedemic activity, haematological effects and histopathological analysis ofSapindusmukorossiGaerten fruits in streptozotocin induced diabetic rats. Asian Pacific Journal of Tropical Medicine, 2012, 5(7): 518- 522.
[6] 黄素梅, 王敬文, 杜孟浩, 张金萍, 姜景民. 无患子籽油脂肪酸成分分析. 中国油脂, 2009, 34(12): 74- 76.
[7] Sun S D, Ke X Q, Cui L L, Yang G L, Bi Y L, Song F F, Xu X D. Enzymatic epoxidation ofSapindusmukorossiseed oil by perstearic acid optimized using response surface methodology. Industrial Crops and Products, 2011, 33(3): 676- 682.
[8] Ollerton J, Lack A J. Flowering phenology: an example of relaxation of natural selection? Trends in Ecology & Evolution, 1992, 7(8): 274- 276.
[9] Abe T. Flowering phenology, display size, and fruit set in an understory dioecious shrub,Aucubajaponica(Cornaceae). American Journal of Botany, 2001, 88(3): 455- 461.
[10] 肖宜安, 何平, 李晓红. 濒危植物长柄双花木开花物候与生殖特性. 生态学报, 2004, 24(1): 14- 21.
[11] 马文宝, 施翔, 张道远, 尹林克. 准噶尔无叶豆的开花物候与生殖特征. 植物生态学报, 2008, 32(4): 760- 767.
[12] Toräng P, Ehrlén J, Ågren J. Habitat quality and among-population differentiation in reproductive effort and flowering phenology in the perennial herbPrimulafarinosa. Evolutionary Ecology, 2010, 24(4): 715- 729.
[13] Rathcke B, Lacey E P. Phenological patterns of terrestrial plants. Annual Review of Ecology and Systematics, 1985, 16: 179- 214.
[14] Barrett S C H. Sexual reproduction inEichhorniacrassipes(water hyacinth). II. Seed production in natural populations[J]. Journal of Applied Ecology, 1980, 17(1): 113- 124.
[15] Zimmerman M, Gross R S. The relationship between flowering phenology and seed set in an herbaceous perennial plant,PolemoniumfoliosissimumGray. American Midland Naturalist, 1984, 111(1): 185- 191.
[16] Kuo Y H, Huang H C, Yang Kuo L M, Hsu Y W, Lee K H, Chang F R, Wu Y C. New dammarane-type saponins from the galls ofSapindusmukorossi. Journal of Agricultural and Food Chemistry, 2005, 53(12): 4722- 4727.
[17] 刁松锋, 邵文豪, 栾启福, 董汝湘, 姜景民. 无患子果皮皂苷含量近红外光谱检测研究. 林产化学与工业, 2014, 34(5): 91- 96.
[18] Du M H, Huang S M, Zhang J P, Wang J W, Hu L S, Jiang J M. Isolation of total saponins fromSapindusmukorossigaerth. Open Journal of Forestry, 2014, 4(1): 24- 27.
[19] 刁松锋, 邵文豪, 姜景民, 董汝湘, 孙洪刚. 基于种实性状的无患子天然群体表型多样性研究. 生态学报, 2014, 34(6): 1451- 1460.
[20] 刁松锋, 邵文豪, 姜景民, 董汝湘, 孙洪刚. 无患子实生群体种实表型性状变异研究. 西北农林科技大学学报: 自然科学版, 2014, 42(5): 75- 83.
[21] 邵文豪, 刁松锋, 董汝湘, 姜景民, 岳华峰. 无患子种实形态及经济性状的地理变异. 林业科学研究, 2013, 26(5): 603- 608.
[22] 邵文豪, 刁松锋, 董汝湘, 孙洪刚, 姜景民, 肖可青, 徐金良. 无患子果实发育动态及内含物含量变化. 林业科学研究, 2014, 27(5): 697- 701.
[23] 刁松锋, 邵文豪, 董汝湘, 孙洪刚, 姜景民. 无患子光合生理日变化及其与生理生态因子的关系. 西北植物学报, 2014, 34(4): 828- 834.
[24] Dafni A, Kevan P G, Husband B C. Practical Pollination Biology. Cambridge, Ontario, Canada: Enviroquest Ltd., 2005: 3- 26.
[25] Pickering C C. Variation in flowering parameters within and among five species of Australian alpineRanunculus. Australian Journal of Botany, 1995, 43(1): 103- 112.
[26] 关文灵, 李叶芳, 陈贤, 杨德. 蝴蝶花花器结构和开花授粉生物学特性. 园艺学报, 2009, 36(10): 1485- 1490.
[27] 范晓明, 袁德义, 唐静, 田晓明, 张旭辉, 王碧芳, 谭晓风. 锥栗开花授粉生物学特性. 林业科学, 2014, 50(10): 42- 48.
[28] 刘丽琴, 李伟才, 王一承, 魏永赞, 石胜友. 龙眼2个品种花部特征比较研究. 热带作物学报, 2012, 33(1): 37- 40.
[29] 柴春山, 芦娟, 蔡国军, 王子婷. 文冠果人工种群的果实发育与落花落果特性研究. 植物研究, 2012, 32(1): 110- 114.
[30] 张敏, 王頔, 张雷, 吕雪芹, 唐亮, 王莉. 文冠果雌雄花发育过程形态结构比较. 电子显微学报, 2012, 31(2): 154- 162.
[31] 王长春, 柯冠武, 黄进华. 东壁龙眼花序发育和花朵分化次序的观察. 福建省农科院学报, 1988, 3(2): 68- 71.
[32] Herrera J. Flowering and fruiting phenology in the coastal shrublands of Doana, south Spain. Vegetatio, 1986, 68(2): 91- 98.
[33] Okullo J B L, Hall J B, Obua J. Leafing, flowering and fruiting ofVitellariaparadoxasubsp.niloticain savanna parklands in Uganda. Agroforestry Systems, 2004, 60(1): 77- 91.
[34] Buide M L, Díaz-Peromingo J A, Guitián J. Flowering phenology and female reproductive success inSileneacutifoliaLink ex Rohrb. Plant Ecology, 2002, 163(1): 93- 103.
[35] 刘曲, 姚小华, 王开良, 任华东, 高焕章, 滕建华, 尹润富, 洪友君. 低海拔地区浙江红花油茶无性系的开花物候特性. 林业科学研究, 2015, 28(2): 249- 254.
[36] 陈芬, 姚小华, 高焕章, 常君, 王开良, 刘曲. 薄壳山核桃不同无性系开花物候特性观测和比较. 林业科学研究, 2015, 28(2): 209- 216.
[37] 李新蓉, 谭敦炎, 郭江. 迁地保护条件下两种沙冬青的开花物候比较研究. 生物多样性, 2006, 14(3): 241- 249.
[38] Blionis G J, Halley J M, Vokou D. Flowering phenology of Campanula on Mt Olynipos, Greece. Ecography, 2001, 24(6): 696- 706.
Flowering phenology of the multipurpose tree speciesSapindusmukorossiGaertn. in low mountain areas of Zhejiang Province
DIAO Songfeng1,2,3, JIANG Jingmin2, YI Huan4, YUE Huafeng1, DONG Ruxiang2, SUN Honggang2, SHAO Wenhao2,*
1ChinaPaulowniaResearchCenter,Zhengzhou450003,China2ResearchInstituteofSubtropicalForestry,ChineseAcademyofForestry,Hangzhou311400,China3No-timberForestryResearchandDevelopmentCenter,ChineseAcademyofForestry,Zhengzhou450003,China4DepartmentofLiangyuanDistrictForestryAdminisration,Shangqiu476000,China
SapindusmukorossiGaertn. is a traditional and important virescent tree species in southern China with peel and seed oil rich in saponins. This tree species is one of the newly developed woody-oil species approved by the State Forestry Administration of China. To investigate the flowering phenology and characteristics of this multipurpose tree species, we observed and compared 9-yr-old plantations ofS.mukorossiat population, individual, inflorescence, and single-flower levels in Zhejiang Province, Tiantai County, from 2012 to 2013, based on the first flowering date, relative flowering intensity, flowering synchrony, and so on. The results showed that the flower color ofS.mukorossiwas yellowish white and the flowers were either male or bisexual. The male flower (5.09 mm in diameter) was larger than the bisexual flower (3.72 mm in diameter). There were usually eight stamens in the male flower; however, a few flowers had seven, nine, or ten. The pistil of the male flower was degenerated and only a slight, light-green protuberance was preserved. The calyx of the bisexual flower folded together lightly and the anthers were under the perianthes. There were generally eight stamens in the bisexual flower, but some also had seven or nine. The chapiter was higher than the anther and was outside the bud. Generally, the ovary had three chambers, but had two or four in rare cases.S.mukorossiflowered earlier in 2012 than in 2013, and the flowering period of the test populations was from the middle of May to late June. The development of the single flower was 8—9 d long from the flower bud to wilting stages. In 2012 and 2013, the florescences of the population, individuals, and inflorescences were approximately 30, 20, and 11 d, and 28, 19, and 13 d, respectively. The flowering amplitude of individuals and inflorescences showed a single peak, sharing a high similarity from year to year. The temporal patterns of flowering ofS.mukorossiwere very similar at the population level, with a well-defined peak and marked synchrony. The index of synchronicity was about 0.868. The relative flowering intensity was distributed over a wide range, but was concentrated between 30% and 40%. The flowering intensity showed an extremely significant (P=0.01) correlation annually and over different years. The range of variation of flowering synchrony was 0.81—0.97 and 0.70—0.98 in 2012 and 2013, respectively. The flowering synchrony correlation was extremely significant (P=0.01) annually, but not in different years. This suggested that there were temporal and spatial differences in the allocation ofS.mukorossireproductive resources, with longer flowering periods and higher flowering synchronization. Long flowering periods can alleviate the interference of illegal pollen and maintain population gene diversity. Heredity is the main factor affecting flowering phenology inS.mukorossipopulations. The effects of ecological and environmental factors on flowering phenology need to be researched further.
SapindusmukorossiGaertn.; inflorescence growth; flowering phenology; relative flowering intensity; flowering synchrony
林业公益性行业科研专项(201404104, 200804032);浙江省-中国林科院省院合作项目(2013SY01);浙江省重大科技专项重点农业项目(2011C12015)
2015- 01- 13;
2015- 08- 28
10.5846/stxb201501130107
*通讯作者Corresponding author.E-mail: wenhaoshao@yeah.net
刁松锋,姜景民,伊焕,岳华峰,董汝湘,孙洪刚,邵文豪.浙江低山地区多用途植物无患子的开花物候特征.生态学报,2016,36(19):6226- 6234.
Diao S F, Jiang J M, Yi H, Yue H F, Dong R X, Sun H G, Shao W H.Flowering phenology of the multipurpose tree speciesSapindusmukorossiGaertn. in low mountain areas of Zhejiang Province.Acta Ecologica Sinica,2016,36(19):6226- 6234.
