苹果枝干轮纹病抗性的相关性及遗传规律研究
2016-11-28李小静张瑞萍阎振立陈迪新张恒涛
李小静+张瑞萍+阎振立+陈迪新+张恒涛+刘珍珍+过国南
摘要:使用树干分段级数加权的方法进行田间自然病情调查,并结合田间接种和离体枝条接种的方法对秦冠×富士杂交F1代进行抗病性鉴定,分析了各抗性指标的次数分布和正态分布,以及各抗性指标间的相关性,研究了苹果杂交F1代枝干轮纹病抗性的遗传规律。结果表明,皮孔密度和大小与轮纹病抗性各指标间没有显著相关性;田间自然感病的级数加权感病高度和加权感病级数与田间接种的感病指数均呈显著正相关;苹果杂交F1代枝干轮纹病感病指数和离体侵染率均表现广泛分离,呈偏正态分布。可见:皮孔密度和大小不宜作为枝干轮纹病抗性的直接鉴定指标;田间接种较离体接种更适合与田间自然病情调查相结合对枝干轮纹病抗性进行评价;苹果枝干轮纹病抗性是数量性状,可进一步对其进行QTL定位等研究,以期更早实现分子抗病性育种。
关键词:苹果杂交F1代;枝干轮纹病;抗性;遗传规律
中图分类号: S661.1 文献标志码: A
文章编号:1002-1302(2016)09-0171-04
苹果枝干轮纹病是由轮纹病菌[Botryosphaeria dothidea(Moug.) Ces. et de Not.]在苹果枝干上侵染而引起的一种病害[1],是目前我国苹果生产上三大病害之一,病害严重时可削弱树势,造成死枝死树。虽然传统防治方法如药剂防治能达到一定的防治效果,但是有些药剂会产生环境污染等问题,应用抗病品种能明显减少病害的发生,可以减少药剂防治可能产生的环境污染等问题的出现,因此开发和利用苹果本身的抗病基因,进行抗病育种和遗传改良,是解决病害的有效途径。
鉴定和评价苹果种质资源对轮纹病的抗病性是选育和利用抗病品种防治轮纹病的基础,苹果抗病性不仅与品种的遗传性有关,与环境条件也有关,即便是同一品种,在不同地点环境下抗病性鉴定结果也可能不一致[2-3]。所以,采用简便、准确、实用、可靠的评价方法对苹果种质资源进行抗病性评价十分必要。目前苹果抗病性的研究方法主要有田间自然病情调查和人工接种鉴定,此外还可通过抗病与感病品种枝条的性状差异进行鉴定。有研究表明,皮孔是病菌入侵的主要途径,通过调查富士苹果枝干轮纹病不同部位的轮纹病发生频率,发现皮孔部位发病频率为92%,因此认为皮孔密度与轮纹病发生程度关系密切[4-5]。李广旭等认为皮孔密度与枝干轮纹病病情指数存在明显正相关[5],阎振立等认为皮孔密度、皮孔密度和皮孔直径之积均与苹果枝干轮纹病的自然发病程度存在显著正相关,可以此来判断品种对枝干轮纹病的抗病程度[6],但韩园园等以红玉×金冠杂种后代为试材发现枝条皮孔密度与枝干接种轮纹病后的病情指数间均未见显著相关性[7]。
迄今为止,国内有关苹果种质资源对轮纹病抗病性的研究报道较多,不同苹果种质资源对苹果轮纹病的抗病性存在明显差异,生产上的富士、金冠、华冠等主栽品种均为感病品种,而秦冠、北之幸、鸡冠对枝干轮纹病具有较强的抗性[6,8-9]。除了了解品种的抗性外,掌握果树性状的遗传规律对于育种工作者来说也很重要,它是进行果树杂交育种的前提和基础。搞清楚性状的遗传特点对于进行基因连锁分析、基因定位和基因图谱的绘制以及有目的地进行基因分离和克隆也是有必要的,但是由于果树基因高度杂合,对其进行病害的遗传研究有一定难度,目前关于轮纹病抗性遗传研究报道相对较少。为了初步分析苹果枝干轮纹病抗性的遗传规律,比较苹果枝干轮纹病抗性的多种评价方式,了解各抗性指标间的相关性,本研究以抗病品种秦冠为母本、感病品种富士为父本的杂交F1代为试材,使用树干分段级数加权的方法进行田间自然病情调查,并结合田间接种和离体枝条接种的方法对苹果杂交F1代轮纹病抗性进行研究,以期获得更简单、有效、准确的苹果轮纹病评价方法,为苹果枝干轮纹病抗性育种提供研究基础。
1 材料与方法
1.1 试验材料
供试材料为中国农业科学院郑州果树研究所苹果育种课题组以秦冠×富士杂交培育的202个实生后代F1代为试材。供试材料于2003年杂交,2004年春播种,立地条件及管理水平相同。
1.2 试验方法
1.2.1 病原菌制备 苹果枝干轮纹病病原菌是由中国农业科学院郑州果树研究所病害研究课题组提供的ZZ26菌株,该菌株从郑州市十里铺果园枝干典型病瘤分离获得[6],通过苹果轮纹病菌的遗传多样性分析,被认为是优势类群[10]。
菌株保存在4 ℃的PDA培养基(葡萄糖20 g、琼脂20 g、马铃薯200 g、蒸馏水1 000 mL)中,接种前1周在25 ℃条件下活化、扩繁,选取长势均匀一致的轮纹病菌用于接种。
1.2.2 田间自然病情调查 田间自然病情调查于2014年5月份进行,采用观察法[11]并结合分段级数加权的方法,逐株调查枝干轮纹病自然发病情况,发病程度按6级标准(表1)进行分级。
测量每棵树树干的总感病高度;判断其由根部开始往上感病部位的不同等级,并测量每个等级所对应的树干长度,二者相乘加权得出级数加权感病高度;级数加权感病高度除以总感病高度得出加权感病级数。
级数加权感病高度=∑(感病高度×感病级数);
加权感病级数=级数加权感病高度/总感病高度。
1.2.3 田间接种抗性鉴定 参照阎振立等的方法[6],用直径为5 mm带有病原菌丝体的培养基块直接接种于枝条表面,并参考其分级标准进行病情调查,计算平均感病指数:
感病指数=100×∑(接种数×该接种处感病级数)/(接种总数×发病最高一级代表值)。
1.2.4 皮孔密度及大小测定 参照于秋香等的方法[12],每株实生树取1年生枝5条,每条枝截取直径为0.6 cm、长为2.0 cm的枝段,40倍显微镜下调查皮孔数目,计算皮孔密度;每根枝条测定5个皮孔纵横径,取平均值计算皮孔面积。
皮孔密度(个/cm2)=皮孔数目/(枝段长×枝段宽);
皮孔面积(mm2)=π×皮孔纵径×皮孔横径/4。
1.2.5 离体接种抗侵染鉴定 离体接种参照周增强的方法[2],以田间光滑无病瘤病斑的粗0.3 cm、长10 cm的一年生枝条作为接种材料,每条枝接种宽0.3 cm、长2 cm菌块,重复3次;接种3 d后参照孙月丽等的方法[13]进行单皮孔培养,每个培养皿接种20个,每个试材接种3皿,25 ℃恒温培养,24 h后于40倍显微镜下观察皮孔周围是否有菌丝生成,有菌丝生成即表示皮孔被侵染,统计侵染情况。
侵染率=被侵染的皮孔数/调查的皮孔数×100%。
1.2.6 统计分析 应用Excel 2003进行数据整理,用SPSS 22.0进行相关性分析。
2 结果与分析
2.1 田间自然抗病性遗传规律
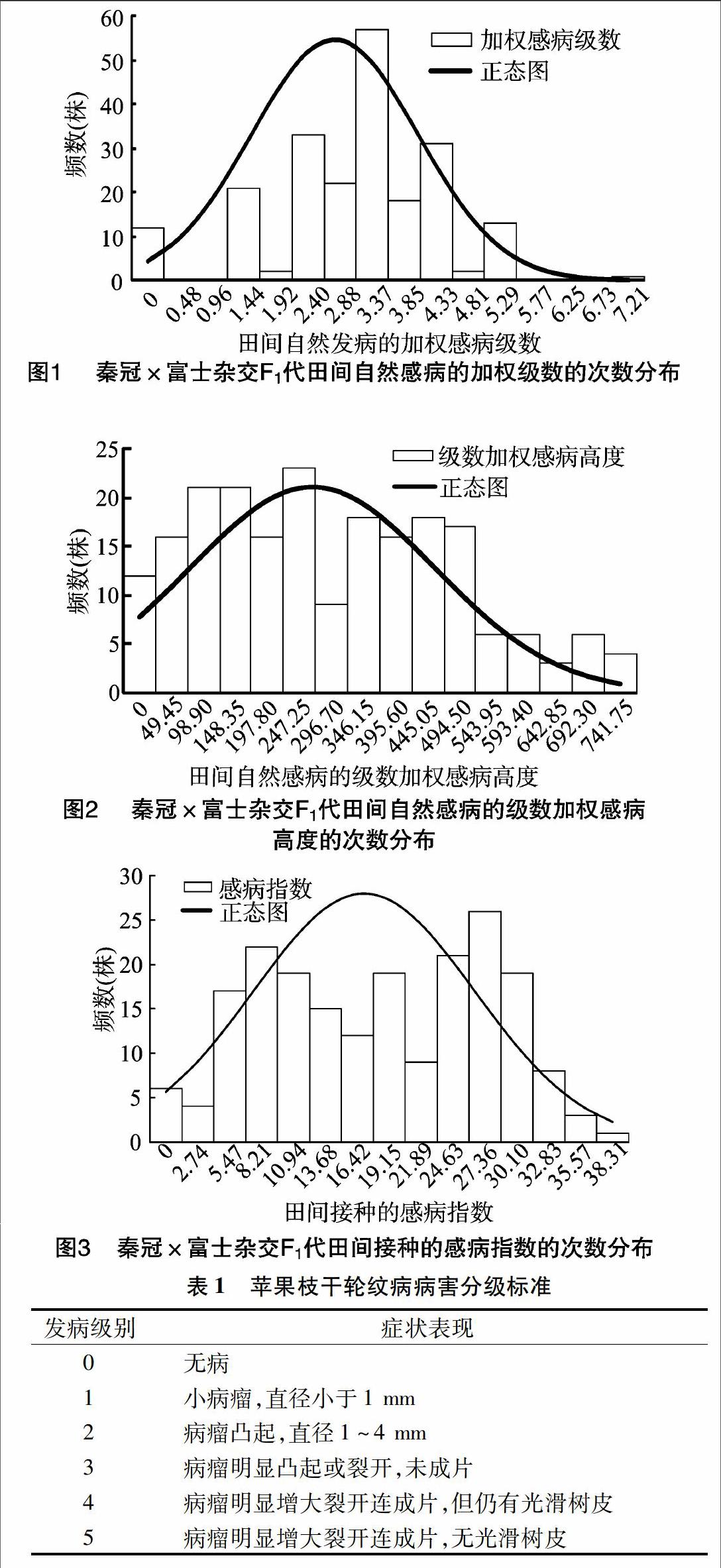
田间自然抗病性鉴定表明,供试的202个单株均发病,加权感病级数在0~7.5之间。不同试材的田间加权感病级数存在较大分离,呈连续、偏正态分布,分布曲线向低加权感病级数偏移(图1)。以加权感病级数≥1为感病、加权感病级数<1为抗病进行分级,抗病/感病的实生树株数的分离比为54 ∶148,卡方检验结果符合1 ∶3的孟德尔分离规律。以级数加权感病高度≥100为感病,级数加权感病高度<100为抗病分级,抗病/感病的实生树株数的分离比为49 ∶153,卡方检验结果符合1 ∶3的孟德尔分离规律(图2)。以上结果可初步确定秦冠×富士杂交F1代田间自然抗病性有数量性状遗传的特点,苹果枝干轮纹病抗性受1对主效基因控制。
2.2 田间接种抗病性遗传规律
采用田间活体枝条接种方法对杂交后代单株进行轮纹病抗病性鉴定,结果表明供试的202个单株的感病指数在0~38.89之间。田间接种的感病指数存在较大分离,呈连续性分布,其次数分布图呈偏正态分布,分布曲线向低感病指数偏移(图3)。以感病指数≥8为感病,以感病指数<8为抗病分级,抗病/感病的实生树株数的分离比为48 ∶154,卡方检验结果符合1 ∶3的孟德尔分离规律。以上结果可初步确定秦冠×富士杂交F1代田间接种抗病性有数量性状遗传的特点,苹果枝干轮纹病抗性受1对主效基因控制。
2.3 皮孔密度和大小
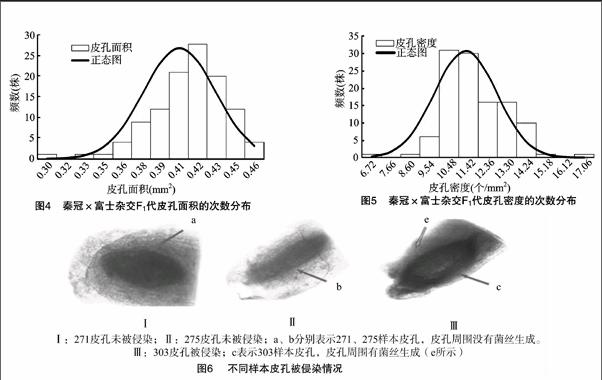
不同试材皮孔密度和大小存在明显差异,调查的113个单株试材间皮孔密度差异较大,从6.72~16.71个/cm2不等,皮孔大小从0.30~0.49 mm2不等,呈连续分布(图4、图5)。
2.4 离体抗侵染能力
对试材的1年生枝条接种后进行单皮孔再分离培养,通过显微镜观察发现,不同试材皮孔侵染情况存在差异,如单株271、单株275皮孔未被侵染,皮孔周围没有菌丝生成(图6-a和图6-b),单株303皮孔被侵染,皮孔周围有菌丝生成(图6-e)。
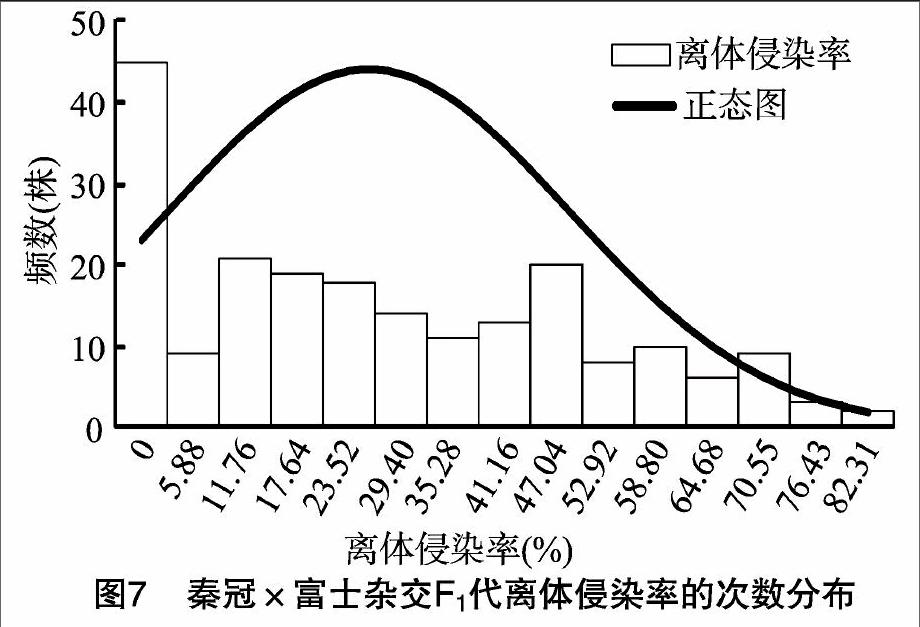
供试的209个单株离体侵染率存在较大分离,其数值从0~85.00%不等,呈连续性分布。以离体侵染率为抗病性指标的次数分布图呈偏正态分布,分布曲线向低离体侵染率偏移(图7)。以侵染率≤30%为抗侵染试材,侵染率>30%为易感染试材进行划分,抗侵染试材/易感染试材的实生树株数的分离比为78 ∶68,卡方检验结果符合1 ∶1的孟德尔分离规律。以上结果可初步确定秦冠×富士杂交F1代离体接种抗轮纹病菌侵染能力有数量性状遗传的特点,苹果枝干轮纹病抗性受1对主效基因控制。
2.5 各鉴定指标间的相互关系
由表2可知,田间接种的感病指数与自然发病的级数加权感病高度、自然发病的加权感病级数的相关系数分别是0.188**、0.159*,呈显著性相关;与皮孔面积的相关系数为0.031,而与离体侵染率和皮孔密度呈负相关。自然发病的级数加权感病高度和加权感病级数的相关系数为0.829**,表现出极显著的相关性。离体侵染率与田间接种感病指数、自然发病的级数加权感病高度、自然发病的加权感病级数和皮孔密度均呈负相关,相关系数分别为-0.215**、-0.216**、-0.307**和-0.093,与皮孔面积呈正相关,相关系数为0.056。皮孔面积与田间接种感病指数、自然发病的级数加权感病高度和自然发病的加权感病级数之间的相关系数为0.031、-0.016和-0.037。皮孔密度与田间接种感病指数、自然发病的级数加权感病高度和加权感病级数之间的相关系数为-0.112、0.160和0.143。自然发病的级数加权感病高度和加权感病级数均与田间接种感病指数均呈显著性相关,说明田间接种的抗性表现情况与单株在自然状态下的抗性情况较为相近。皮孔面积、皮孔密度与离体侵染率以及田间接种和自然发病2种感病方式下的抗性指标并没有同时呈现出显著相关性,仅有皮孔面积与离体侵染率呈正相关,说明皮孔面积和密度并不从根本上决定苹果枝干轮纹病的抗病,而植株本身的抗性所起的决定作用更大。同时也认为不宜直接把皮孔的形态特征指数作为评价苹果枝干轮纹病菌抗性的鉴定指标。
3 讨论与结论
3.1 抗性评价方式选择
在植物抗病性遗传研究中,田间自然感病情况能够十分直观地体现植株的综合抗性,虽然无法排除树势、其他病害或环境条件等因素的影响,但它仍然是抗病研究中较为重要的参考指标[14]。但也有研究表明杂种实生树抗病性采用活体接种鉴定方法所得结果的准确性好[4,15],许多学者将它与田间自然病情调查结合起来对轮纹病抗性进行评价。本研究借鉴前人的田间调查法,测定了级数加权感病高度,然后计算出加权发病级数。该测定方法考虑到了主干发病高度的差异、相同发病高度下发病级数的差异,旨在尽量消除植株本身的长势差异导致的树干本身高度的不同,更客观地体现出植株本身对轮纹病的抗性。
本研究中田间接种和自然发病的抗性趋势一致、呈显著正相关,这与周增强等的研究结果[2]一致。阎振立等研究显示自然调查和田间接种鉴定结果具有极显著相关性[6],本研究中二者相关性未达到极显著水平,可能是由于本研究选的材料为重复发病的多年生植株,不仅枝干轮纹病菌群复杂,而且树体生长又受到其他病害影响。
本试验采用离体枝条接种单皮孔再分离的方法对轮纹病抗侵染能力进行了鉴定,研究结果表明离体接种与自然调查鉴定结果没有显著相关性。孙月丽等研究结果发现不同抗性试材的抗侵染能力与抗病性基本一致,但也存在抗侵染能力与抗病性不统一的情况[13]。
前人研究认为皮孔密度与轮纹病发生程度关系密切,皮孔密度或皮孔密度与直径之积可以作为鉴定轮纹病抗性的形态指标[3-4]。本研究结果表明皮孔形态指标与轮纹病抗性指标间无显著相关性,不宜作为枝干轮纹病抗性的直接鉴定指标,这与韩园园等以红玉×金冠后代分离群体为试材的研究结果[7]一致。
3.2 苹果枝干轮纹病抗病性遗传规律
本研究中秦冠×富士杂交F1代个体轮纹病抗性有较大的分离,呈偏正态分布,属于1对主效基因控制多基因共同作用的数量性状遗传。苹果枝干轮纹病抗性在杂交后代有较大分离,同时分离呈偏正态分布,这与郭兴科等以西府海棠×珠美海棠、西府海棠×S19、西府海棠×P22杂交后代为试材的研究结果[3]相同。本研究认为苹果轮纹病抗性为1对主效基因控制多基因共同作用的数量性状,而刘志认为苹果轮纹病为单一主效基因控制的质量性状[16]。在苹果斑点落叶病的研究中,Saito等认为苹果斑点落叶病的抗病性由隐性单基因控制[17],而赵磊等认为该病抗性由2对主效基因控制多基因共同作用,同时受加性效应控制[18],这种差异可能是抗病性评价标准不同及杂种实生群体的遗传背景不同造成的[19]。本试验田间自然总感病指数和田间接种感病指数的孟德尔分离比均为1 ∶3,离体侵染率的孟德尔分离比为1 ∶1,这说明秦冠的苹果枝干轮纹病的抗性和抗侵染性的控制基因及其遗传规律是不同的。
参考文献:
[1]Tang W,Ding Z,Zhou Z Q,et al. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is botryosphaeria dothidea[J]. Plant Disease,2012,96(4):486-496.
[2]周增强,侯 珲,王 丽,等. 枝干苹果轮纹病人工接种方法与品种抗性评价[J]. 果树学报,2010,27(6):952-955.
[3]郭兴科,史 娟,李方方,等. 不同苹果砧木杂交后代对轮纹病的抗性及对“红富士”苹果抗病性的影响[J]. 北方园艺,2014(4):22-25.
[4]李广旭,高艳敏,杨 华,等. 轮纹病菌在苹果枝干上侵入途径的扫描电镜观察[J]. 果树学报,2005,22(2):169-171.
[5]李广旭,沈永波,高艳敏,等. 皮孔组织结构及密度与苹果枝干粗皮病发生的关系[J]. 果树学报,2004,21(4):350-353.
[6]阎振立,张全军,张顺妮,等. 苹果品种对轮纹病抗性的鉴定[J]. 果树学报,2005,22(6):654-657.
[7]韩园园,庄 艳,李春敏,等. 苹果实生树枝干对轮纹病菌抗病性反应的差异[J]. 中国农业大学学报,2012,17(5):69-75.
[8]张玉经,王 昆,王 忆,等. 苹果种质资源果实轮纹病抗性的评价[J]. 园艺学报,2010,37(4):539-546.
[9]沙守峰,伊 凯,刘 志,等. 不同苹果品种及杂交组合对粗皮病发生程度的影响[J]. 北方园艺,2005(2):52-53.
[10]彭 斌,刘丽锋,吴会杰,等. 苹果轮纹病菌种内遗传多样性研究[J]. 中国农业科学,2011,44(6):1125-1135.
[11]刘海涛,李彩丽,赵永波,等. 苹果育种杂种初选轮纹病抗性评价及遗传分析[J]. 江苏农业科学,2011,39(2):241-244.
[12]于秋香,邵建柱,徐继忠. 苹果砧木杂交后代皮孔性状与抗轮纹病菌侵染的关系[J]. 河北农业大学学报,2010,33(2):21-25.
[13]孙月丽,于秋香,徐继忠. 皮孔的组织结构与苹果枝干轮纹病抗性的关系[J]. 河北农业大学学报,2011,34(6):55-59.
[14]党志国,赵政阳,万怡震,等. 苹果杂交F1代抗褐斑病遗传趋势及抗性选择研究[J]. 西北农林科技大学学报:自然科学版,2010,38(5):137-142.
[15]阎振立,张恒涛,张全军,等. 苹果抗轮纹病遗传的初步研究[J]. 河南农业科学,2009(8):105-108.
[16]刘 志. 苹果抗轮纹病种质资源和基因筛选以及NO介导的防御响应[D]. 泰安:山东农业大学,2009.
[17]Satio K I,Niizeki M. Fundamental studies on breeding of the apple:Ⅺ. Genetic analysis of resistance to Alternaria blotch(Alternaria mali Roberts)in the interspecific crosses[J]. Bulletin of the Faculty of Agriculture,Hirosaki University,1988,50:27-34.
[18]赵 磊,赵政阳,党志国,等. 秦冠、富士苹果杂交后代抗早期落叶病的遗传分析[J]. 西北农业学报,2008,17(2):197-201.
[19]吕 松,王 忆,张新忠,等. 苹果斑点落叶病抗病性种质评价及遗传分析[J]. 中国农业大学学报,2012,17(4):68-74.
