一株葡萄藻的分离、鉴定及产烃性能评价
2016-11-12陈传红尹顺吉马建源
陈传红 吴 洪 尹顺吉 马建源 李 青
( 新奥科技发展有限公司,煤基低碳能源国家重点实验室,廊坊 065001)
一株葡萄藻的分离、鉴定及产烃性能评价
陈传红吴洪尹顺吉马建源李青
( 新奥科技发展有限公司,煤基低碳能源国家重点实验室,廊坊 065001)
研究从我国海南博鳌海边的淡水池塘水样分离获得一株绿藻ENN41。显微形态观察表明,ENN41的形态特征属于葡萄藻。进一步克隆ENN41的核糖体小亚基18S rRNA片段,利用分子生物学软件进行比对分析,结果表明ENN41的18S rRNA基因序列与布朗葡萄藻(Botryococcus braunii)同源性最高,说明ENN41为一株布朗葡萄藻(B. braunii)。ENN41在柱式反应器中培养16d,单位体积产率为0.483 g/(L·d),粗烃占干重的含量为56.6%; 主要脂肪酸为油酸(C18:1)、十八碳四烯酸(C18:4)和棕榈酸(C16:0),三者之和占总脂肪酸的72.6%; 利用尼罗红染色,清晰可见大量的油脂分布在细胞内和胞外基质中。ENN41在板式反应器中培养16d,单位体积产率为0.234 g/(L·d),粗烃占干重的含量为20.0%。上述研究表明,ENN41是具有较高的生长速度和粗烃积累能力的布朗葡萄藻(B. braunii)藻株,具有产业化应用潜力。
ENN41;18S rRNA;布朗葡萄藻;单位体积产率;粗烃含量
随着全球化石能源的短缺和化石能源使用带来的环境问题,寻找新型可替代能源势在必行,微藻具有生物量大、生长周期短及含油脂类物质高等特点,在CO2减排和生物燃料利用上都有广阔的开发前景[1—3]。
目前利用微藻生产生物柴油主要有两种研发思路,一种是利用微藻合成的脂肪酸酯化来生产生物柴油,大多数的微藻通过脂肪酸合成途径能够积累大量油脂,目前大多数从事微藻能源研究的团体均采用此类研发思路来开发微藻生物柴油[4,5]; 另一种是利用微藻合成的烃类物质来生产生物柴油,因烃类物质与石油性质相似,故可代替石油成为新的可再生能源。迄今所发现的具有产烃能力的藻类有葡萄藻(B. braunii)[6—8]、盐藻(Dunaliena salina)[9]、小球藻(Chlorella vugaris)[10]、网翼藻(Dictyopteris aerostichoides)[11]等,但葡萄藻因其含烃量高、烃类的组成和结构与石油极为相似之故,对葡萄藻的研究受到世界的广泛关注。
葡萄藻又名丛粒藻,属绿藻门(Chlorophyta)、四胞藻纲(Trebouxiophyceae)、葡萄藻科(Botryo coccaceae)、葡萄藻属(Botryococcus),是一种世界性分布的淡水或微咸水绿藻[12]。葡萄藻烃含量为细胞干重的25%—86%,烃的性质因藻株有所区别[13]。葡萄藻的含烃量明显高于其它微生物,但葡萄藻生长缓慢,室外养殖容易污染,很难得到大规模的推广应用,因此,优良葡萄藻藻种是实现葡萄藻推广应用的关键。
本研究以作者分离自海南的绿藻ENN41为实验材料,通过显微形态观察,并结合18S rRNA序列特征,对ENN41进行了形态分类和分子鉴定。同时利用不同光反应器研究了ENN41的生长及烃积累情况,为藻株的下一步开发利用提供了依据。
1 材料与方法
1.1材料
藻株的分离本研究于2010年12月在海南博鳌海边淡水池塘采集水样,取无菌水稀释水样后,吸取一定量的样品滴在载玻片上,用40倍或100倍显微镜观察,将藻体移入视野中间,用毛细管直接吸取藻细胞,接种于48孔装有BG11培养基的培养板中,在25℃、光照强度为50 μmol/(m2·s)条件下培养2—3周,后期进行显微镜观察,选择只有单藻株的孔,铺平板及平板划线进一步分离纯化目的藻株,得到纯藻种。经显微镜形态学观察初步鉴定为葡萄藻属的种类,命名为ENN41。
1.2方法
藻种培养藻种的培养采用BG11培养基,反应器为内径50 mm、长度600 mm的柱式反应器,光照强度100 μmol/(m2·s),24h光照,培养温度25—28℃,在培养过程中通过向培养液中通入1%—2%的二氧化碳和空气的混合气体,将培养基的pH控制在7—9。
藻种分子鉴定基因组的提取:采用液氮研磨和试剂提取的方法获得,DNA溶于试剂盒所带Elution Buffer。具体如下:将ENN41藻株用350目滤网先将藻液浓缩,然后再离心收集,尽量除去细胞沉淀中的多余水分,冻存于-20℃。用液氮研磨冻存的藻细胞,研磨的同时,加入适量玻璃砂。使用Biospin Genomic DNA Extraction Kit试剂盒提取待鉴定的ENN41藻株的基因组DNA,使用试剂盒附带的Elution Buffer洗脱,1%琼脂糖凝胶电泳检测其浓度和纯度。
18S rRNA 序列的扩增:采用真核生物18S扩增通用引物(引物合成由上海生工生物工程公司合成)扩增微藻基因组18S基因片段。(上游引物18SF:5′-CCTGGTTGATCCTGCCAG-3′; 下游引物18SR:5′-TTGATCCTTCTGCAGGTTCA-3′)。取1 μL总DNA为模板,PCR反应条件为:95℃预变性5min,然后94℃变性50s,56℃退火50s,72℃延伸90s,反应30个循环,最后72℃延伸10min。1.0%的琼脂糖凝胶电泳检测其扩增产物,回收纯化目标DNA片段,并将目标片段连接到pMD18-T载体上(TAKARA,Japan) ,转入E. coli感受态细胞,使用引物对18SF/R检验出阳性克隆送至上海生工公司测序。
序列分析与系统进化树的构建用BLAST软件对编辑后的基因序列的同源性进行分析。采用Clustal W软件进行多序列比对,MEGA6.0进行Clustal W同源比对,然后采用Neighbor-Joining算法,Bootstrap值为1000,构建系统进化树,所用藻株信息如表1所示。
实验方法将对数生长期藻细胞接种到实验用的光生物反应器中(分别为内径30 mm、长度600 mm的柱式反应器和50 cm×5 cm×50 cm板式反应器),光强约为200 μmol/(m2·s),24h光照,温度28—30℃,通入含1%—2%CO2的压缩空气将pH值控制在7—9,定时取适量藻液,进行相应的指标检测。
分析方法生物量的测定:将Ф47 mm的Whatman(GF/C)滤膜于105℃烘干至恒重后称重(W1),于设定的取样时间移取一定体积(V)的藻液真空抽滤,在105℃的烘箱中烘干过夜,于干燥器中冷却后称重(W2)。生物量浓度用单位体积藻体的干重(Dry cell weight,DCW)表示。
生物量(DCW,g/L)=(W2-W1)/V
尼罗红染色:取培养16d的藻液0.5 mL于4000 r/ min离心5min,去上清,并用去离子水洗涤2次,后用20% DMSO水溶液重新悬浮至0.5 mL,加入尼罗红使其浓度为1 μg/L,35℃水浴10min,后用去离子水洗涤一次,此时将染色的藻细胞置于荧光显微镜下观察。
粗烃的测定:将培养16d的藻液于4000 r/min离心5min,藻泥经真空冷冻干燥获得冻干藻粉。取5.00 g冻干藻粉放置在圆底烧瓶中,加入200 mL氯仿,45℃水浴抽提8h,期间定时摇动烧瓶,使其充分混匀,抽提完后用定性滤纸过滤去除藻渣,用氯仿溶剂洗涤藻渣2次以上。收集氯仿相于旋转蒸发仪40℃水浴中去除氯仿,剩余物即为粗烃。
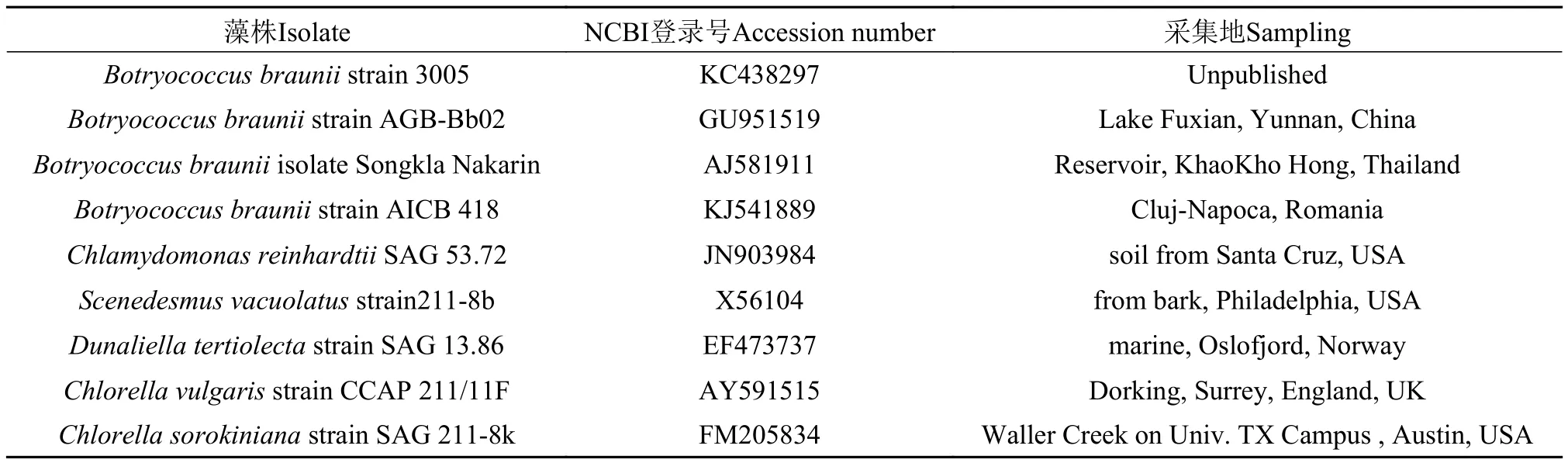
表1 构建18S系统树所用藻株信息Tab. 1 Strain information for construction of 18S phylogenetic tree
脂肪酸组成及含量的测定:(1)脂肪酸提取:取100 mg冻干藻粉放置在具Telfnon螺口瓶盖的体积为15—20 mL的小玻璃瓶中,再放置一小磁力棒,加入2—4 mL 10% DMSO-Methanol 溶液,40℃砂浴(盛砂的烧杯放置恒温加热磁力搅拌器上)5min; 然后在4℃下磁力搅拌抽提30min,3500 r/min离心,转移上清液到另一小瓶中。剩下藻渣再加入1 :1的乙醚、正己烷4—8 mL 4℃下磁力搅拌抽提1h,3500 r/min离心,转移上清液到上述一小瓶中。可重复上述过程直到藻渣变白。在上述合并抽提液中加入纯水使四者(水、DMSO-Methanol、乙醚、正己烷)体积比例为 1 :1 :1 :1,震荡分相,移取有机相转移到另一小玻璃瓶中,在通风橱中用氮气吹至成浓缩液,然后转移到事先称重过的1.5 mL塑料离心管中,再用氮气吹干至恒重。(2)脂肪酸分析:照上面方法进行提取后,用正己烷溶解,使用Agilent 6820气相色谱仪进行气相色谱分析(色谱条件为载气:氮气流量1 mL/min、氢气流量30 mL/ min、空气流量300 mL/min,进样口温度:280℃,检测器温度:280℃,检测器类型:Agilent FID,色谱柱:Agilent DB-5毛细管色谱柱(30 m × 0.25 mm,0.25 μm),分流比:4 :1。分析方法:内标法GB/T 17377-1998(气相色谱用氮气作载气,相当于液相色谱的流动相)。
2 结果
2.1ENN41的形态学特征
在显微镜下观察到ENN41的藻细胞形状为椭圆形或楔形(图1),几个或上百个细胞通过不规则且长短不同的绳索状透明胶质相连,组成大小不一的集结体; 细胞大小:长6—8 μm,宽5—7 μm,聚集群体一般在50 μm以上,大的可达数百μm至肉眼可见; 细胞颜色为深绿色或黄褐色,稳定期的细胞呈黄褐色; 细胞包被在不规则分枝或分叶的、半透明的胶群体胶被里,细胞顶部通常裸露在外; 稳定期的群体细胞经在挤压后有明显的油滴从胶被里渗出; 有时观察可见细胞分裂,以似亲孢子生殖,大的群体比较易于断裂为小群体。
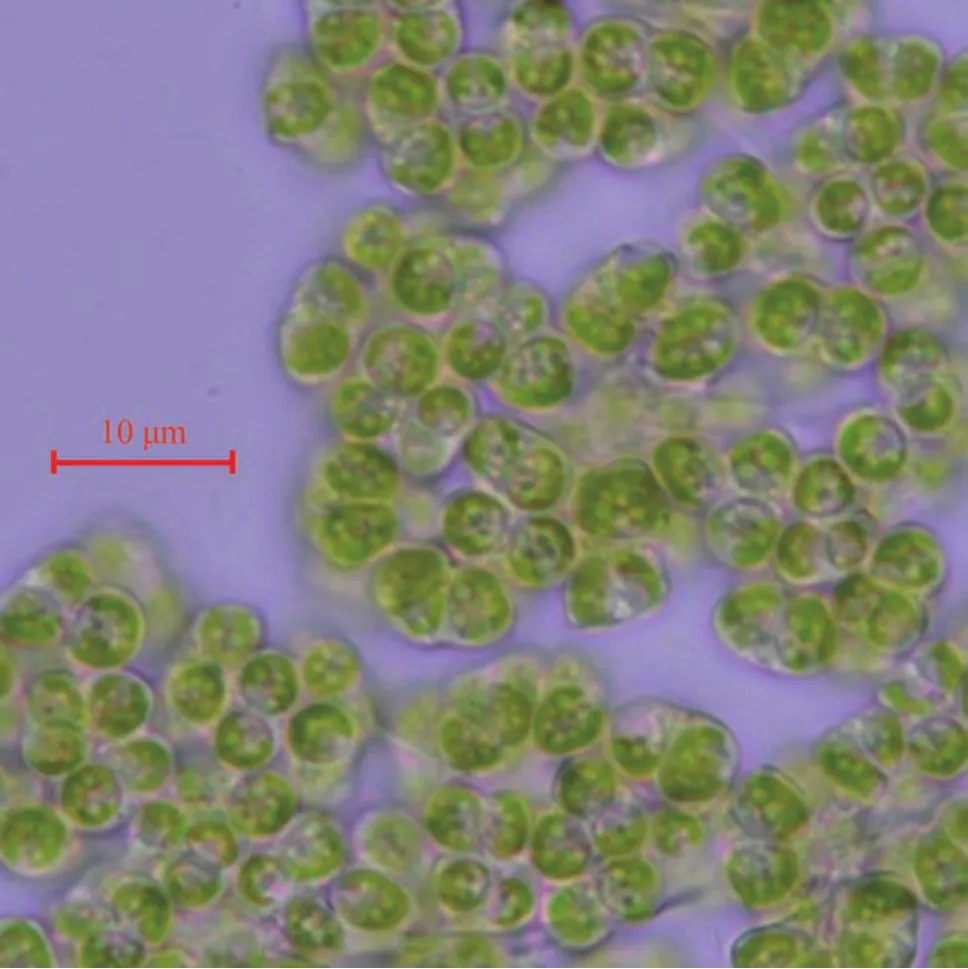
图1 ENN41在正常生长下显微照片Fig. 1 Micrograph of ENN41 under normal growth condition
2.2ENN41 18S rRNA基因序列分析
用真核生物18S rRNA扩增通用引物对18SF/R获得的ENN41的18S rRNA序列,登录GenBank数据库进行BLAST比对,结果显示与索引号为KC438297.1的布朗葡萄藻B. braunii strain 3005 的18S rRNA基因序列最为相似,匹配度为99%,覆盖率为100%。
根据上述18S rRNA序列Blast结果,选取葡萄藻和其他绿藻相关基因序列信息绘制了ENN41的18S rRNA基因系统进化树(图2)。从图2中可以看出,ENN41与布朗葡萄藻B. braunii strain 3005和B. braunii strain AGB-Bb02较近,与B. braunii isolate Songkla Nakarin和B. terribilis strain AICB 418较远。
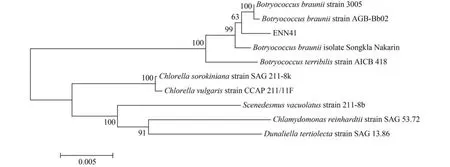
图2 根据18S Blast基因序列构建布朗葡萄藻ENN41系统进化树Fig. 2 Phylogenetic tree of Botryococcus braunii ENN41 based on the results of 18S Blast
2.3光反应器中ENN41的生长特性
柱式反应器ENN41在柱式反应器中持续培养16d,分别于4、7、10、13和16d取样测定OD值和生物量。ENN41在16d的培养中一直呈现快速生长趋势,OD750从接种时的0.300增加到2.705,平均每天增加0.150(图3a); 生物量终浓度为8.710 g/L,单位体积产率为0.483 g/(L·d)(图3b)。
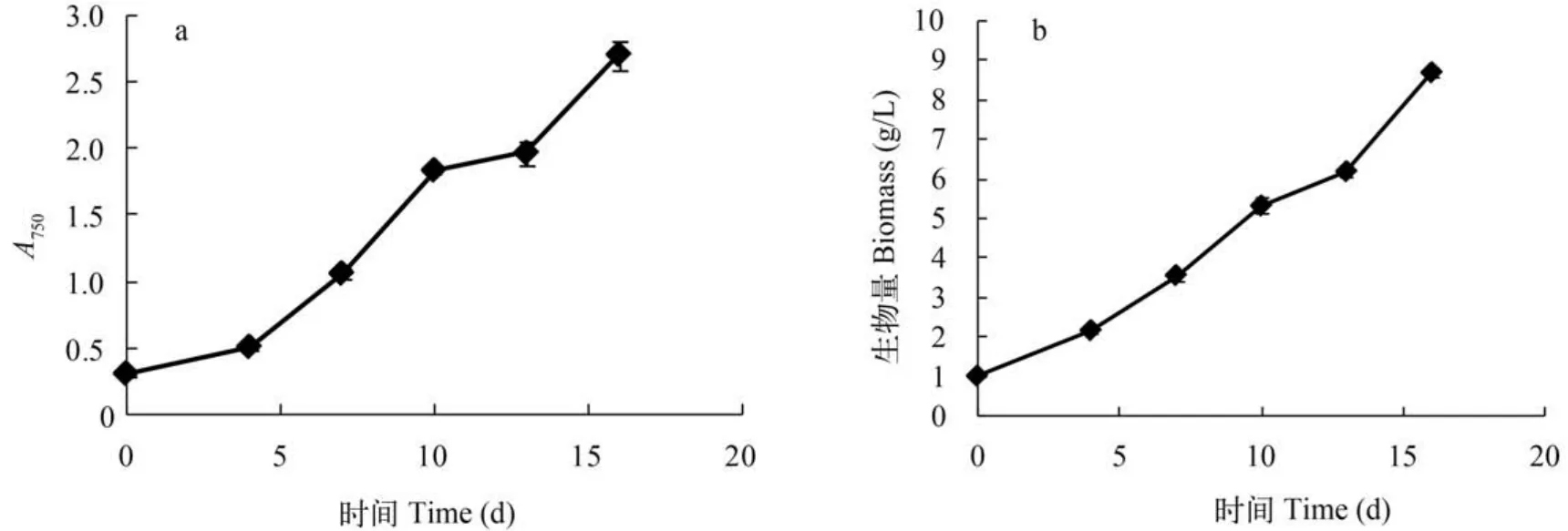
图3 ENN41在柱式反应器中OD值(a)和生物质浓度(b)的变化Fig. 3 The OD750(a) and biomass concentration(b) of ENN41 in the column reactor
如表2所示,ENN41的粗烃占干重的含量在16d时为56.6%。从表观上看,粗烃提取前藻粉呈褐绿色,提取后变为绿色,而提取出的粗烃在正己烷溶剂中呈褐色,说明在培养后期藻粉中积累了一定量的类胡萝卜素,使后期藻粉和粗烃溶液带有一定的褐色。

表2 ENN41在柱式反应器中培养16d时的粗烃含量Tab. 2 The crude hydrocarbon of ENN41 at the end of the 16d culture in column reactor
如表3所示,培养16d时ENN41的脂肪酸占干重的含量仅为5%,其中主要脂肪酸为油酸(C18:1)、十八碳四烯酸(C18:4)和棕榈酸(C16:0),三者之和占总脂肪酸组成的72.6%。

图4 粗烃提取前(a)、后(b)的ENN41藻粉及正己烷溶解的粗烃(c)Fig. 4 The extraction of crude hydrocarbon of ENN41. Algal power before(a) and after(b) extraction of ENN41,crude hydrocarbons dissolved in hexane(c)
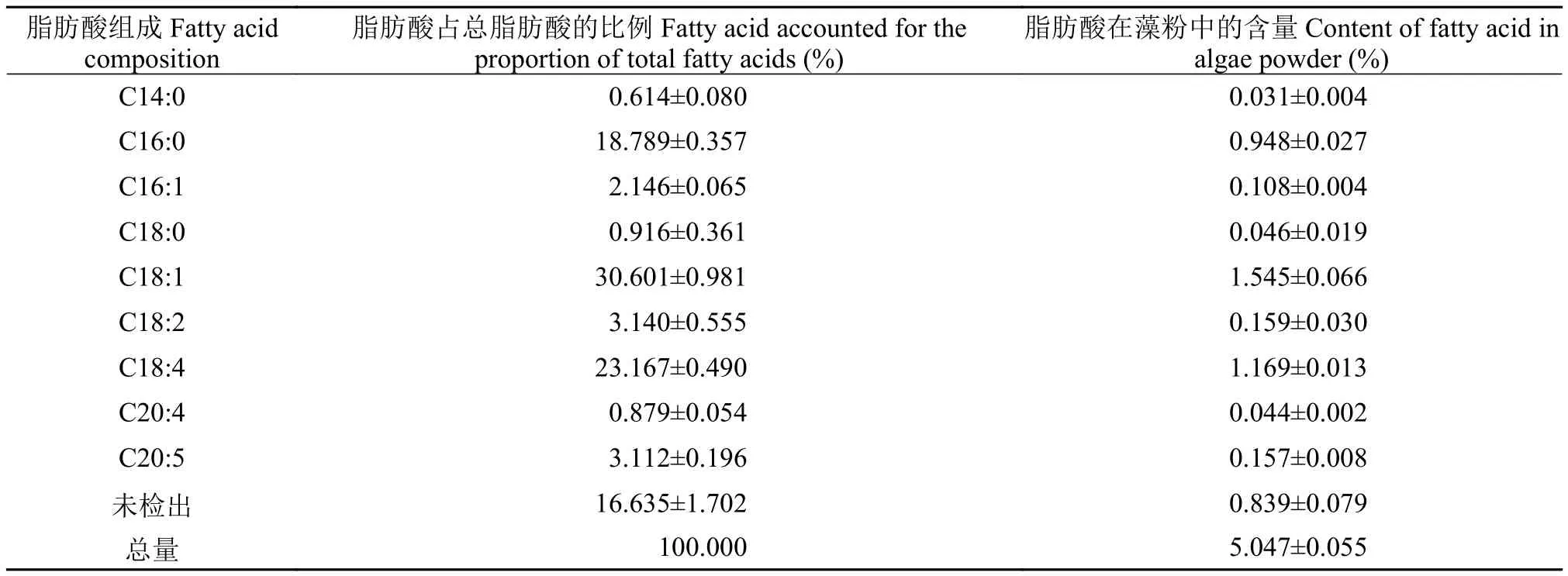
表3 ENN41在柱式反应器中培养16 d时的脂肪酸组成及含量Tab. 3 Fatty acid composition and content of ENN41 at day 16
收集在柱式反应器中培养了16d的ENN41藻体,用尼罗红染色。如图5所示,可以清楚的看见大量的油脂分布在胞外基质中,少部分分布在细胞内。
板式反应器ENN41在光径5 cm的板式反应器中持续培养16d,每隔1天取样测定生物量浓度,于培养结束时测定粗烃含量。ENN41在整个培养过程中生物量一直呈逐渐增加趋势,16d时的生物量为4.584 g/L,单位体积产率为0.234 g/(L·d),粗烃占干重的含量为20%。ENN41在放大的光生物反应器中,虽然与窄光径、小体积的柱式反应器相比,产量降低了许多,但仍然保持了相对较快的生长速度。这说明ENN41在放大的培养体系中,仍然保持了较好的生长性能,另外,本次放大实验未使用营养盐胁迫诱导条件,仍然获得了较高的烃含量。
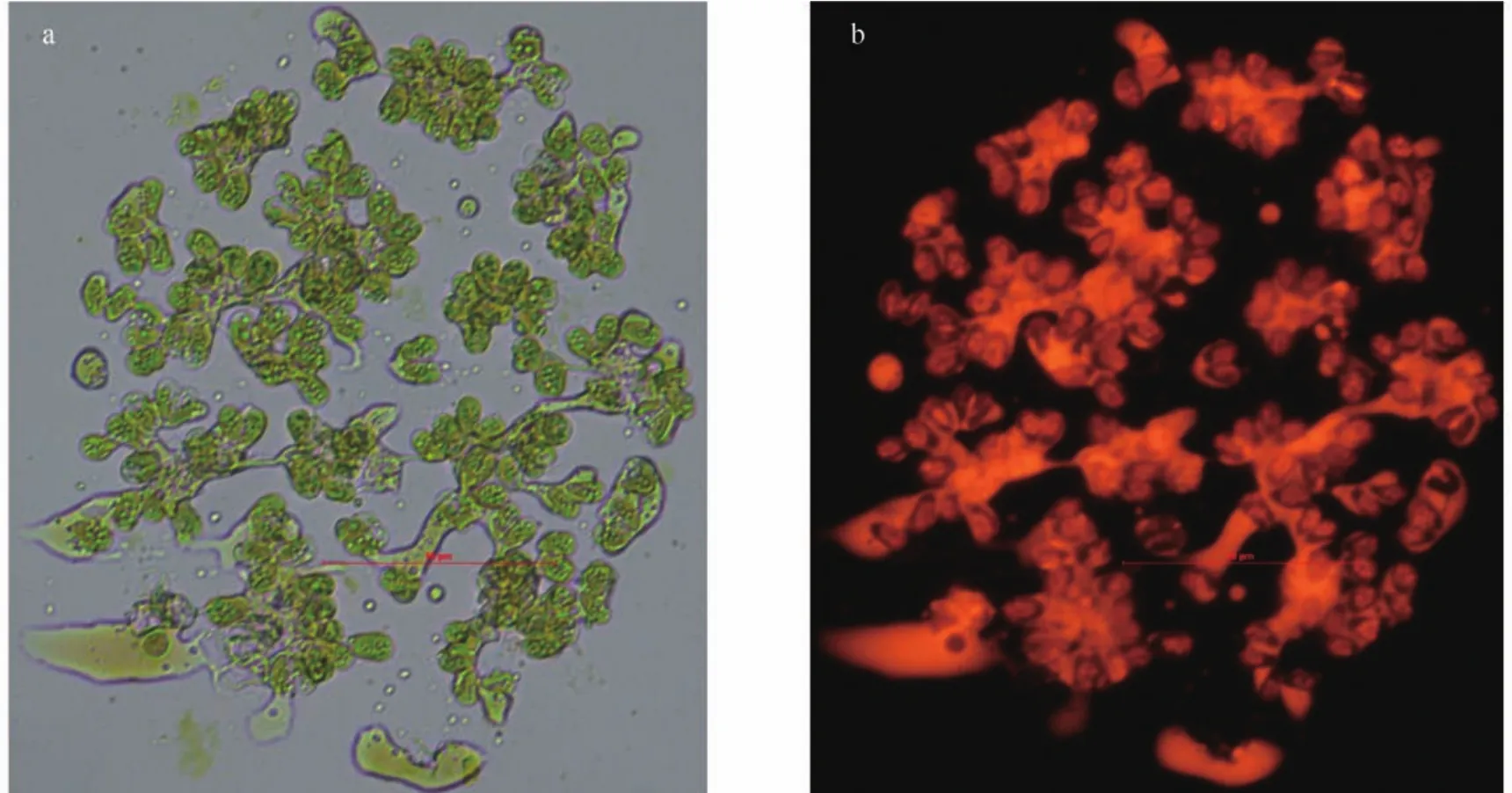
图5 布朗葡萄藻ENN41的尼罗红染色图Fig. 5 The Nile red staining of Botryococcus braunii ENN41
3 讨论
微藻形态分类需要较长周期的观察,并要求研究者有丰富的藻类学知识,部分微藻常因培养条件的改变而发生形态的变化,从而影响微藻的形态学分类。利用基因序列分析可克服在形态上变化和形态难以区别的藻株,对微藻的分类提供依据,结果的准确性也更高,同时有利于厘清同源性较高的藻株间的进化距离。本研究使用的ENN41在培养过程中也会发生细胞大小和集落大小的改变,但总体生物学形态特征与刚分离时相差不大。本研究利用18S rRNA基因序列对ENN41分类作了进一步的鉴定,系统进化树证明分离自海南的藻株ENN41与布朗葡萄藻B. braunii strain 3005和云南抚仙湖的AGB-Bb02进化距离最短,与另外几株葡萄藻藻株之间的进化距离要远一些,说明相似生境来源的藻株的同源性也会较高,而不同生境来源的藻株之间存在遗传多样性。这与王朋云等[14]报道的不同地理来源的葡萄藻藻株的形态和遗传多样性结果一致。
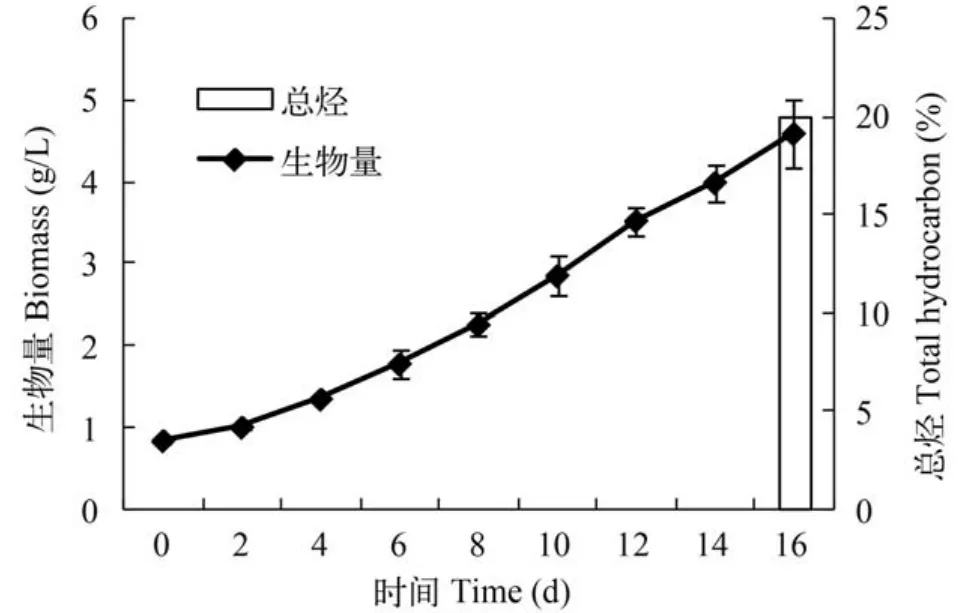
图6 ENN41在板式反应器中的生长曲线和16d时的粗烃含量Fig. 6 The growth curve of ENN41 in plate reactor and the crude hydrocarbon content over 16 days
关于葡萄藻培养方面,各国研究者已经开展了许多工作,主要集中在培养基和营养元素及环境条件的研究上。可用于培养葡萄藻的培养基主要有:Chu10、Chu13、Prat、BBM、BG11等,这些培养基主要在大量元素含量上差别较大,本文采用了营养成分较充分且淡水藻常用的BG11培养基。另外,在葡萄藻培养中,光照、温度、盐度、pH等环境因子对其生长和产烃量有很大影响。光强大小不仅影响微藻的色素比例,而且影响了葡萄藻的生长速度和产烃量。Zhang等[15]在较高光强[约140 μmol/(m2·s)]下培养布朗葡萄藻,生物量和产烃量均高于低光强下的数值,Oyama等[16]在中等光强[140—400 μmol/(m2·s)]下更有利于布朗葡萄藻的生长和产烃。温度对布朗葡萄藻的影响具有种属特异性[17],通常最适生长温度是23—25℃。pH是对微藻生具有重要影响的因素之一,pH主要影响无机碳在水中的溶解形式,而影响藻细胞的生长。本研究培养布朗葡萄藻所用的培养条件为光强200 μmol/(m2·s),温度25℃左右,pH控制在7—9,与上述文献描述的布朗葡萄藻生长条件基本吻合,有效提高了ENN41藻株的生长和产烃量。一直以来,许多研究者认为葡萄藻生长速度较慢,是限制其产业化应用的主要原因。其中Banerjee等[18]认为,葡萄藻生长速度慢的主要原因是因为藻细胞合成高热值的碳氢化合物,而不是由于营养物的缺乏。本研究利用柱式反应器培养ENN41藻株,16d时生物量达到了8.710 g/L,单位体积产率达到0.483 g/(L·d),同时,粗烃占干重的含量也达到了56.6%,表明合成高热值碳氢化合物的ENN41同时具有较高的生长速度。布朗葡萄藻ENN41具有的高生长速度、生物量积累能力和产烃能力在国内布朗葡萄藻的研究中还是首次报道,并且,这样的结果与利用相似培养条件和柱式反应器的王元丽等[19],高保燕等[20]报道的真眼点藻、栅藻产量相似,也就是说,布朗葡萄藻ENN41的生长速度和生物量积累能力能够赶上以生长快速、生物量积累能力高著称的真眼点藻和栅藻了。日本的研究者Kojima等[21]报道利用柱式光反应器(内径为7 cm,8.5 cm高的圆锥形底部)培养布朗葡萄藻,在反应器内壁光强约为140 μmol/(m2·s),培养25d,其生物量达到了7 g/L,烃含量为干重的50%。另有文献报道在鼓泡塔反应器中用猪舍废水培养布朗葡萄藻,可以有效的去除水中的氮磷,且藻细胞的干重达到了8.5 g/L,产烃量为0.95 g/L[22]。上述国外报道和作者关于布朗葡萄藻ENN41的研究结果表明,优良的布朗葡萄藻藻株在合适的培养条件下,也具有高速生长、积累高生物量的潜力和高产烃能力。
徐玲等[23]报道不同培养基对布朗葡萄藻脂肪酸组成没有影响,其主要脂肪酸组成为棕榈酸(C18:1)、油酸(C18:1)和亚麻酸(C18:3)。而Rao等[24]报道在Chu13培养基中主要脂肪酸为硬脂酸(C18:0)和亚油酸(C18:2),在培养基中添加一定浓度的NaCl后主要脂肪酸变为棕榈酸(C16:0)和油酸(C18:1)。而本研究中布朗葡萄藻ENN41培养后期主要脂肪酸为油酸(C18:1)、十八碳四烯酸(C18:4)和棕榈酸(C16:0)。由此可见,不同株系的布朗葡萄藻其主要脂肪酸可能不同,且同一株布朗葡萄藻在不同培养条件或不同培养基下,主要脂肪酸可发生改变,但布朗葡萄藻的主要脂肪酸都是由C18和C16组成。
本研究中的布朗葡萄藻ENN41利用尼罗红染色发现,大量的油脂分布在胞外基质中,少部分分布在细胞内。有文献报道利用尼罗红对烃类进行染色观察,发现有大量油滴主要分布在胞外基质中,细胞内也清晰可见少量油滴的分布[25,26]。这与本研究染色基本结果一致,说明胞外基质是布朗葡萄藻ENN41储藏烃类物质的主要场所。Largeau C等[27]通过电子显微镜观察进一步表明,烃类在胞外基质中积累量可达到95%,此时细胞内积累的烃类仅为5%左右。
4 结论
从海南博鳌海边的淡水池塘中分离获得的ENN41是一株布朗葡萄藻藻株。显微形态观察表明,ENN41的形态特征属于葡萄藻; ENN41的18S rRNA基因序列与布朗葡萄藻的18S rRNA基因序列同源性最高。ENN41在柱式反应器中培养16d,生物量为8.710 g/L,单位体积产率为0.483 g/(L·d),粗烃占干重的含量为56.6%; 主要脂肪酸为油酸(C18:1)、十八碳四烯酸(C18:4)和棕榈酸(C16:0),三者只和占总脂肪酸组成的72.6%; 利用尼罗红染色,可以清晰看见大量的油脂分布在细胞内和胞外基质中。ENN41在放大的板式反应器中培养16d的单位体积产率、生物量和粗烃含量也达到了较高水平。本研究结果表明,ENN41是具有较高的生长速度和粗烃积累能力的布朗葡萄藻藻株,具有产业化应用潜力。
[1]Williams P,Laurens L. Microalgae as biodiesel and biomass feedstocks:review and analysis of the biochemistry and economics [J]. Energy & Environmental Science,2010,3(5):554—590
[2]Chisti Y. Biodiesel from microalgae [J]. Biotechnology Advances,2007,25(3):294—306
[3]Singh J,Gu S. Commercialization potential of microalgae for biofuels production [J]. Renewable & Sustainable Energy Reviews,2010,14(9):2596—2610
[4]Miao X L,Wu Q Y. Study on preparation of biodiesel from microalgal oil [J]. Acta Energiae Solaris Sinica,2007,28(2):219—222 [缪晓玲,吴庆余. 微藻油脂制备生物柴油的研究. 太阳能学报,2007,28(2):219—222]
[5]Li L,Zheng L,Zheng M G,et al. Effect on lipid accumulation of marine oleaginous microalgae under CO2enrichment cultivation [J]. Acta Hydrobiologica Sinica,2013,37(6):1013—1019 [李林,郑立,郑明刚,等. 富碳培养对海洋富油微藻油脂积累特性的影响. 水生生物学报,2013,37(6):1013—1019]
[6]Templier J,Largeau C,Casadevall E. Variations in external and in ternal lipids associated with inhibition of the resistant biopolymer from the a race of Botryococcus braunii [J]. Phytochemistry,1993,33(5):1079—1086
[7]Metzger P,Largeau C. Botryococcus braunii:a rich source for hydrocarbons and related ether lipids [J]. Applied Microbiology and Biotechnology,2005,66(5):486—496
[8]Motohide I,Masahiro O,Nobuyoshi N,et al. Isolation of herbicide-resistant mutants of Botryococcus braunii [J]. Bioresource Technology,2012,109:300—303
[9]Tornabene T G,Holzer G,Peterson S L. Lipid profile of the halophilic alga Dunaliella salina [J]. Biochemical and Biophysical Research Communications,1980,96:1349—1356
[10]Wu Q Y,Yin S,Sheng G Y,et al. New discovery for hererotrophic etiolation algae to study on producing hydrocarbon [J]. Science in China(Series B),1993,23(4):423—429 [吴庆余,殷实,盛国英,等. 异养黄化藻类产烃研究新发现. 中国科学B辑,1993,23(4):423—429]
[11]Wirth D,Fischer-Lui I,Boland W,et al. Absolute configuration and synthesis of dextro caudoxirene the gametereleasing and gamete-attracting pheromone of the brown alga Peritha liacaudata Phaeophyceae [J]. Helvetica Chimica Acta,1992,75:751—758
[12]Banerjee A,Sharma R,Chisti Y,et al. Botryococcus braunii:a renewable source of hydrocarbons and other chemicals [J]. Critical Reviews in Biotechnology,2002,22(3):245—279
[13]Senousy H H,Beakes G W,Hack E. Phylogenetic Placement of Botryococcus braunii(Trebouxiophyceae) and Botryococcus sudeticus isolate UTEX 2629(Chlorophyceae) [J]. Journal of Phycology,2004,40(2):412—423
[14]Wang P Y,Mao Y X,Kong F N,et al. Morphological and genetic diversity of Botryococcus braunii [J]. Periodical of Ocean University of China,2011,41(5):63—70 [王朋云,茅云翔,孔凡娜,等. 丛粒藻形态多样性与遗传多样性研究. 中国海洋大学学报,2011,41(5):63—70]
[15]Zhang K,Eiichi K. Effect of light intensity on colony size of microalga Botryococcus braunii in bubble column photobioreactors [J]. Fermentation and Bioengineering,1998,86(6):573—576
[16]Oyama H,Kim S,Park E R. Effects of light intensity and nutrients on the growth of Botryococcus sp. [J]. Misaengmul Hakhoechi,1997,25(3):339—343
[17]Wang X Y,Xie S H. Effects of several factors on Botryococcus braunii [J]. Microbiology,1996,23(5):275—277[王修垣,谢树华. 几种因子对从粒藻株A的效应. 微生物学通报,1996,23(5):275—277]
[18]Banerjee A,Sharma R,Chisti Y. Botryococcus braunii:a renewable source of hydrocarbon and other chemicals [J]. Critical Reviews in Biotechnology,2002,22(3):245—279
[19]Wang Y L,Li Q Y,Li A F,et al. Growth and photosynthetic physiological characteristics of four Eustigmatophycean species [J]. Biotechnology,2014,24(2):91—95[王元丽,李其雨,李爱芬,等. 4株真眼点藻的生长及光合生理特性. 生物技术,2014,24(2):91—95]
[20]Gao B Y,Shen D D,He S S,et al. Integrated the biomassproduction of oleaginous microalga Scenedesmus acuminatus and dairy wastewater treatment [J]. Renewable Energy Resources,2014,32(5):673—679 [高保燕,沈丹丹,何思思,等. 富油微藻——尖状栅藻生物质生产与奶牛场废水处理相结合的效果研究. 可再生能源,2014,32(5):673—679]
[21]Kojima E,Zhang K. Growth and hydrocarbon production by microalga Botryococcus braunii in bubble column photobioreactor [J]. Journal of Bioscience and Bioengineering,1999,87:811—815
[22]An J Y,Sim S J,Lee J S,et al. Hydrocarbon production from secondarily treated piggery wastewater by the green alga Botryococcus braunii [J]. Journal of Applied Phycology,2003,15(2—3):185—191
[23]Xu L,Liu C Z,Li H Z. Medium selection of Botryococcus braunii and its metabolic mechanism [J]. Chinese Journal of Bioprocess Engineering,2010,8(6):40—45[徐玲,刘春朝,李华钟. 布朗葡萄藻的培养基选择及其产物代谢规律. 生物加工过程,2010,8(6):40—45]
[24]Rao A R,Dayananda C,Sarada R,et al. Effect of salinity on growth of green alga Botryococcus braunii and its constituents [J]. Bioresource Technology,2007,98(3):560—564
[25]Tanoi T,Kawachi M,Watanabe M M.Effects of carbon source on growth and morphology of Botryococcus braunii [J]. Journal of Applied Phycology,2011,23:25—33
[26]Tanoi T,Kawachi M,Watanabe M M. Iron and glucose effects on the morphology of Botryococcus braunii with assumption on the colony formation variability [J]. Journal of Applied Phycology,2014,26:1—8
[27]Largeau C,Casadevall E,Berkaloff C,et al. Sites of accumulation and composition of hydrocarbons in Botryococcus braunii [J]. Phytochemistry,1980,19(6):1043—1051
ISOLATION,IDENTIFICATION AND EVALUATION OF HYDROCARBON PRODUCING PERFORMANCE OF ONE BOTRYOCOCCUS STRAIN
CHEN Chuan-Hong,WU Hong,YIN Shun-Ji,MA Jian-Yuan and LI Qing
(State Key Laboratory of Coal-Based Low-Carbon Energy,Xinao Scientific & Technological Developmental Co.,Ltd.,Langfang 065001,China)
ENN41 is a freshwater strain isolated from Bo'Ao coast of Hainan Province. Microscopic observation showed that ENN41 has the unique morphological character of genus Botryococcus. The 18S rRNA gene of ENN41 was cloned for further species confirmation at molecular level. The homologous analysis showed that the 18S rDNA gene of ENN41 was highly homologous to the species of Botryococcus braunii in GenBank. ENN41 achieved a biomass productivity of 0.483 g/(L·d) and crude hydrocarbon content of 56.6% for 16 days culture in column reactor. The main fatty acids were oleic acid(C18:1),octadecatetraenoic acid(C18:4) and palmitic aid(C16:0),accounting for 72.6% of the total fatty acid. Large amount of lipid stained with Nile red in the extracellular matrix. ENN41 reached a high productivity after 16 days culture in plate bioreactor [0.234 g/(L·d)] with 20% crude hydrocarbon. All the results showed that ENN41 is a natural Botryococcus braunii strain with high productivity of biomass and lipids and high potential for industrial application.
ENN41; 18S rRNA; Botryococcus braunii; Biomass productivity; Crude hydrocarbon
Q949.2
A
1000-3207(2016)05-1012-08
10.7541/2016.131
2015-08-21;
2016-01-05
国家“973”项目(2012CB723606); 国家重大科学仪器设备开发专项(2012YQ15008715)资助 [Supported by the National Basic Research Program of China(973 Program)(2012CB723606); the National Key Scientific Instrument and Equipment Development Project(2012YQ15008715)]
陈传红(1982—),女,山东人; 硕士; 研究方向为藻类生物技术及应用。E-mail:chench@enn.cn
吴洪,博士; E-mail:wuhong@enn.cn
