洱海三种生活型水生植物叶片中碳、氮与总酚含量的比较研究
2016-11-12袁昌波周存宇倪乐意张霄林
袁昌波 曹 特 周存宇 倪乐意 张霄林
(1. 长江大学,荆州 434025; 2. 中国科学院水生生物研究所,武汉 430072)
洱海三种生活型水生植物叶片中碳、氮与总酚含量的比较研究
袁昌波1,2曹特2周存宇1,2倪乐意2张霄林2
(1. 长江大学,荆州 434025; 2. 中国科学院水生生物研究所,武汉 430072)
植物体内的C、N和总酚是影响其被牧食和凋落物降解的重要因素,从而影响C和N在食物链的传递和生物地化循环。本研究测定了云南洱海3种生活型(挺水、沉水和浮叶)26种水生植物叶片中C、N和总酚含量以及C/N比率,并分析了它们与总酚的关系。结果表明:挺水植物叶片的C、N和总酚平均含量为448.08、39.30和24.70 mg/g,浮叶植物叶片的C、N和总酚平均含量为433.32、35.16和28.05 mg/g,沉水植物叶片的C、N和总酚平均含量为378.36、27.31和10.28 mg/g; 总体上看,植物叶片C和N含量:挺水植物> 浮叶植物> 沉水植物; 挺水和浮叶植物叶片的总酚含量差异不显著,且均远高于沉水植物叶片的总酚含量; 26种植物叶片中C和N与总酚含量均呈显著正相关。在富营养化条件下,3种生活型植物所处生境的光照和CO2供给均差异显著,会对这些植物叶片的C、N和总酚含量有一定影响,此外由于应对生境中的胁迫(草食性昆虫、病原体及紫外线辐射等)压力以及自身的生长策略的不同也可能导致C、N和酚在三者中形成差异; 通过对13种沉水植物叶片总酚含量比较,推测光叶眼子菜和微齿眼子菜较为适合作为洱海耐牧食恢复先锋物种; N与总酚正相关的关系可能在富营养化进程中加速湖泊沼泽化。
洱海;大型水生植物;C;N;总酚
水陆交错带蕴含繁多的动植物群落和旺盛的初级生产力,对维持湖泊生态系统的物质与能量循环并发挥健康的生态服务功能具有重要意义[1]。生长于其中的大型水生植物作为湖泊生态系统主要的初级生产者(Sculthorpe,1967)[2],在维持湖泊清水稳态方面发挥着重要作用。例如,水生植物具有产氧、吸收氮磷、吸附水体悬浮物和抑制浮游藻类生长等功能,可以减轻水体富营养化和提高水体自净能力,同时还能为水生动物提供食物来源和附着微生物提供基质,从而维持水体物种多样性[3]。Wetzel[4]按这些植物在水中生存方式的不同,将其分为挺水植物(Emergent macrophyte)、漂浮植物(Freely-floating macrophyte)、浮叶植物(Floatingleaved macrophyte)和沉水植物(Submerged macrophyte)。它们因对淹水的依赖性不同而分布在水陆交错带不同的区域,挺水植物、浮叶植物和漂浮植物分布在相对浅水的区域,它们是鸟类、哺乳动物和昆虫的重要食物;沉水植物则完全生活在水面以下,其分布的水深最深,具有相较于前者更低的光饱和点和光补偿点[5],生理上符合阴生植物特点,为浮游生物及食草性鱼类等提供食物来源。四者在介导陆域与水域之间的物质交换和能量中有着重要作用。
C、N是生物体的大量元素,其含量的多少和比率可以影响到植物的生长、竞争、营养价值、草食者的偏好以及凋落物的降解速率[6]等多个方面,并且还影响到植物体内总酚的含量。同时,已知的研究表明,酚类可以影响牧食者的口感和偏好[7,8],抗氧化[9—11],抗病菌[12],植物碎屑的降解速率[13]从而影响生物地球化学循环。除种间差异外,3种生活型水生植物体内C、N和总酚的含量也受到生长环境中的各种因子(例如光照、富营养化、空气中CO2浓度、食草性动物的捕食等)的影响。因此,富营养化将会影响水陆交错带植物的群落组成和演替进程,从而影响水陆交错带这些植物类群的C、N和总酚含量,改变湖泊的元素循环和水陆间的物质交换。
C、N与总酚对水体生态系统的物质循环有着重要意义,但目前关于C、N与酚类之间的关系及其对生态系统物质循环影响的研究却很少。Haukioja等[14]在研究毛枝桦叶片中总酚和N含量与生长,虫害以及落叶后恢复能力的关系的过程中指出,毛枝桦叶片中N含量与总酚含量呈负相关。Lima等[15]也指出叶片寿命越长,总酚含量越高,而N含量越低。上述研究中总酚与植物生长大量元素之间的关系基于陆生植物,而目前关于水生植物的相关研究却未有报道。
本研究通过对云南洱海26种不同生活型(挺水、沉水和浮叶)水生植物叶片中C、N及总酚含量测定,并分析了各生活型之间C、N及其比率与总酚的关系,试图了解富营养化水体中3种生活型水生植物利用C、N营养元素及总酚含量变化的特点和其形成的环境原因,水生植物叶片中C、N营养元素与总酚的关系与陆生植物相比有何不同以及其在湖泊生态系统物质循环中的作用。这在洱海由中营养向富营养转型关键时期[16,17]和当前水生植物与湖泊生态系统之间关系的基础研究缺乏的情况下具有重要意义。
1 材料与方法
1.1现场调查、样品采集及测定方法
在本研究中,所有植物样品均在2013年9月中旬采集于洱海北部的红山湾和沙坪湾区域(图1),该区域地势平缓,挺水植物、浮叶植物和沉水植物发育充分,植物种类丰富。因此,在该区开展研究可以部分避免因气候差异(如温度、降雨、光照等)和牧食者组成(昆虫、草食动物等)的不同对研究结果的影响。此外,由于该区域周边人口密集且农业和养殖业发达,自20世纪90年代以来该区域的水体经历快速富营养化,且藻类水华在某些年份偶发[18]。在采集样品期间,水生植物已处于生长的后期,生物量大,叶已发育成熟。
水生植物采自水深3 m至最高水位线所覆盖的区域,相邻采样点间距离约30 m,用水下镰刀(采集面积约0.2 m2)和剪刀等采集不同生活型(挺水,浮叶和沉水)水生植物。每个采样点的植物按种类分开,洗去泥沙和附着物,沥干浮水,取植物成熟完整的叶片若干,存放于保温箱带至实验室。当天采集的样品用清水洗净,吸水纸擦干后,置于80℃烘箱中烘烤至恒重。烘干的植物叶片用研钵研磨至粉末状,用元素分析仪(Flash EA1112 series,CE Instruments,Italy)测定C、N含量,用Folin-Denis法测定总酚含量[19]。
图1 本研究中植物样品采集的区域Fig. 1 Sampling area and location for aquatic plants in the study
1.2统计方法
采用Microsoft Excel 2013和SPSS 17.0 Software Package进行数据图表制作和统计分析,各物种取平均值后进行生活型之间差异比较及所有植物的相关性分析,C、N、C/N及总酚含量的比较采用One-way ANOVA分析,总酚与C、N含量和C/N比值的关系显著性采用Pearson相关分析。
2 结果
在本研究中,共采集植物种类26种,样本总数226(表1)。
2.1三种生活型水生植物叶片的C、N和总酚含量及C/N比率
挺水植物叶片的C、N和总酚的平均含量分别为(448.08±2.84)、(39.30±3.42)和(24.70±7.24) mg/g,浮叶植物的C、N和总酚平均含量分别为(433.32± 9.16)、(35.16±2.52)和(28.05±12.42) mg/g,沉水植物的C、N和总酚平均含量分别为(378.36±14.86)、(27.31±1.40)和(10.28±1.81) mg/g。总体上看,C、N平均含量:挺水植物> 浮叶植物> 沉水植物(P< 0.05);挺水和浮叶植物叶片的总酚平均含量差异不显著,但均远高于沉水植物(P< 0.05)。挺水和浮叶植物叶片的C/N比率平均值分别为12.11和12.57,二者间差异不显著,但均低于沉水植物(P< 0.05)(图2)。
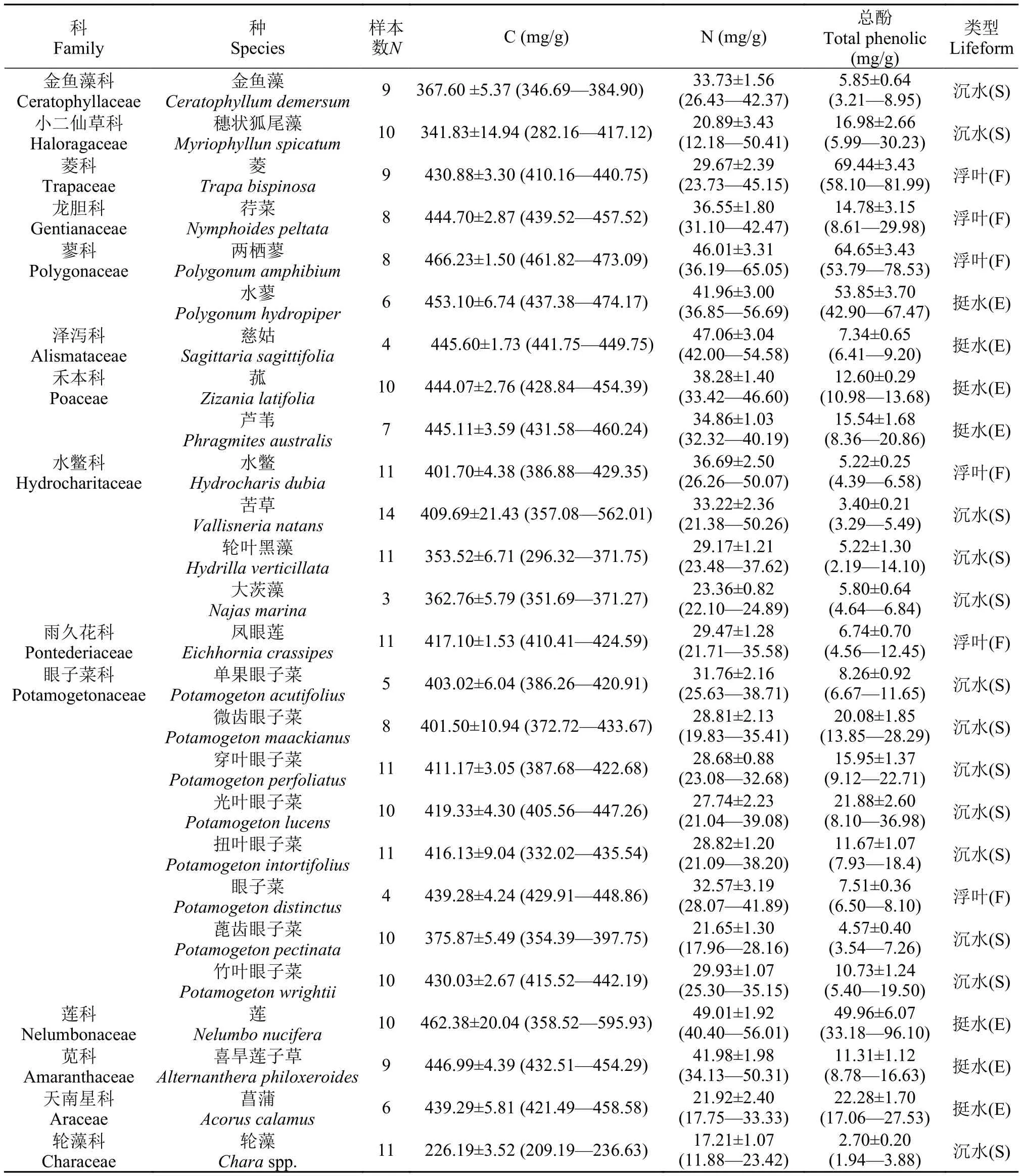
表1 洱海不同生活型水生植物及其叶片的C、N和总酚含量(平均值±标准误)Tab. 1 Taxonomy,growth form,the contents of C and N,and total phenols in leaves of 26 aquatic plants collected in Lake Erhai(means±SE; N for number of replicate samples; E for emergent,F for floating-leaved and S for submerged)
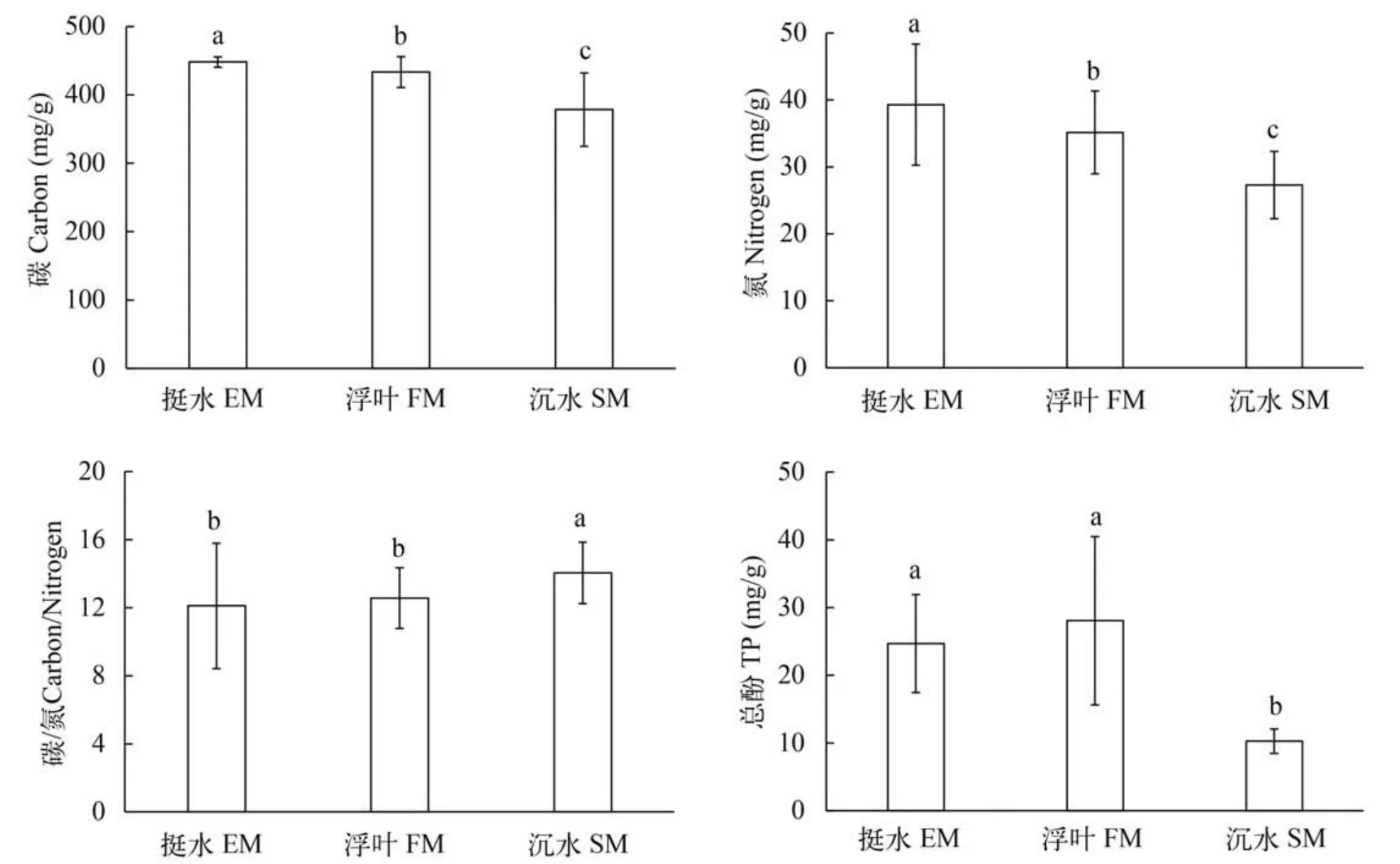
图2 三种生活型水生植物叶片中C、N和总酚含量及C/N比率Fig. 2 The C and N Content,total phenols as well as C/N ratio in leaves of emergent,floating-leaved and submerged macrophytes in Lake Erhai(TP for total phenolic; EM for emergent macrophytes,FM for floating-leaved macrophytes and SM for submerged macrophytes)
2.2沉水植物种间比较
沉水植物种间的比较中,从单一物种来看,C、N、C/N及总酚平均含量最高和最低的分别是竹叶眼子菜和轮藻、金鱼藻和轮藻、蓖齿眼子菜和金鱼藻以及光叶眼子菜和轮藻,其范围分别为226.19—430.03、17.21—33.73、10.90—17.36以及2.70—21.88 mg/g(图3)。
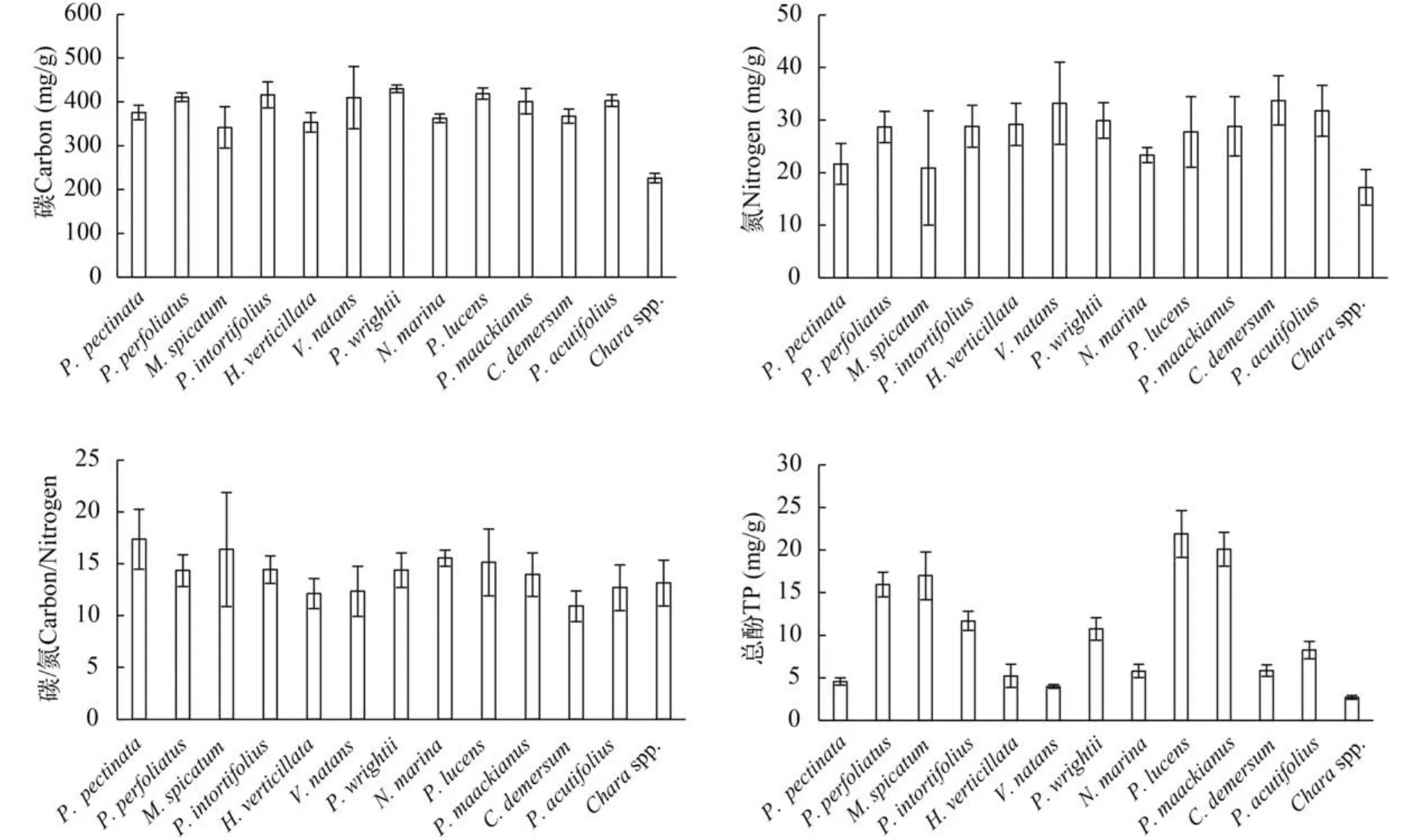
图3 沉水植物物种间C、N、C/N及总酚含量比较Fig. 3 The C and N content,C/N ratio and total phenols in leaves of submersed macrophytes in Lake Erhai(TP for total phenolic)
2.3生活型之间C、N及C/N与总酚相关性分析
Pearson相关性分析表明:所有植物叶片中C和N含量分别与总酚含量之间具有显著的正相关性(P< 0.05),C/N比率与总酚之间无显著相关性(P>0.05)(图4)。
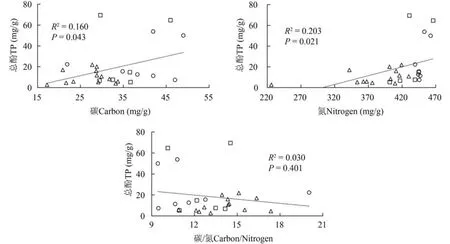
图4 三种生活型水生植物叶片中总酚含量与C和N含量及C/N比率的相关性(〇挺水植物;□ 浮叶植物;△ 沉水植物)Fig. 4 The relationship of total phenols a,C and N contents as well as C/N ratios in leaves of emergent,floating-leaved and submersed macrophytes in Lake Erhai(〇emergent;□ floating-leaved;△ submerged)
3 讨论
3.1不同生活型水生植物叶片C、N和总酚含量差异的原因
根据2013年9月洱海水质月常规监测所得数据,洱海北部湖湾红山湾和沙坪湾TN含量分别为0.87—1.05和0.83—0.94 mg/L,TP含量分别为0.03—0.06 mg/L和0.02—0.04 mg/L,叶绿素含量分别为14.50 —33.48 μg/L和18.41—32.36 μg/L(未发表),两采样地水体的N、P营养浓度,叶绿素含量以及周围环境条件相似,实验所得结果具有一定可比性。洱海水生植物叶片中C和N含量在3种生活型水生植物排序为:挺水>浮叶>沉水。该结果与黄亮等[20]研究结果一致,但N含量在3种生活型水生植物的排序与吴爱平等[21]相反。吴爱平等[21]研究对象包括中营养型到重富营养型的19个湖泊,而沉水植物在水体营养增加的过程中会过量吸收养分,因此本研究中N在3种生活型植物的分布特点与吴爱平等人研究的结果不同的原因之一可能是所研究湖泊的数量和营养类型不同。9月为洱海水生植物生长的后期,洱海水体中浮游植物和附着藻类随气温和水体营养含量的升高而达到一年中较高的生物量,水体透明度不足1.5 m(未发表),加上浮游藻类对CO2的竞争,这严重影响了沉水植物对光和CO2的利用,而挺水植物和浮叶植物叶片直接暴露于空气中,其生长不受光照条件和CO2的限制。因此,3种生活型水生植物所处生境的光照和CO2供给或部分地影响了它们的C含量,在洱海水体中可供植物利用的氮磷营养充足时,挺水植物和浮叶植物的生长和生物量均远大于沉水植物。植物的快速生长有赖于叶片的光合作用效率,而光合效率又与叶片的N含量密切正相关,因而3种生活型植物在生长和生物量上的差异可能造成它们叶片N含量的差异:挺水植物> 浮叶植物> 沉水植物。
在所有植物种类中,总酚2.70—69.44 mg/g,平均含量为18.27 mg/g,菱最高,轮藻最低;从生活型上看浮叶植物最高,挺水植物居中,沉水植物最低。不同生活型水生植物间总酚含量的差异与Smolders等[7]和Cao等[8]的报道一致。一般认为,植物体内酚类含量受到光照和营养等因素的影响。浮叶植物相比沉水植物有着更高的N含量,而挺水植物则有比沉水植物更高浓度的热量物质,因而二者比之沉水植物更容易吸引草食者。而高浓度的酚可以有效降低草食性动物对植物的牧食行为,如Lodge[22]研究发现植物组织中总酚含量与草食者的牧食行为具有显著负相关性。同时,Cronin和Lodge[23]以及Dudt和Shure[24]发现高光环境显著增加水生植物叶片的总酚含量。因此,本研究中3种生活型植物所处生境的草食者的牧食压力以及光照环境差异或是造成其叶片中总酚含量差异的原因之一;此外,云南高原地区高紫外线辐射也可能增加植物叶片的总酚含量。
3.2沉水植物总酚含量的种间差异及意义
沉水植物是湖泊生态系统中与水体关系最为密切的水生植物,对系统物质循环和能量流动有着重要的影响。长期以来,沉水植物因其营养价值低和口感不佳而不受草食者的欢迎,许多学者甚至认为牧食对沉水植物影响微乎其微[1]。Lodge等[25]在对前人的研究做了总结和比较后,指出牧食作用对水生植物的影响与对陆生植物的影响同等重要,都会减少植物现存生物量。前人研究所关注的主要是小型的牧食者,然而大型牧食者(如:水鸟/禽、鱼、虾和蟹等)对水生植物的每日牧食量却很可观[26]。目前已有研究显示,大型牧食者对沉水植物群落的生长和演替都有着重要影响[25—27],而酚含量高的沉水植物其被牧食的可能性相对会小。因此,酚类化合物含量的高低可以在一定程度上反映沉水植物环境适应能力强弱的有效指标。通过对洱海13种沉水植物叶片总酚含量比较后发现,光叶眼子菜和微齿眼子菜总酚的平均含量最高,且二者均为冠层型沉水植物,对光和水体中N、P营养物质的利用效率高于其他类型沉水植物(未发表数据),因而推测二者为洱海耐牧食恢复先锋物种。
3.3C、N与总酚相关性及其生态学意义
C和N分别与总酚含量显著正相关,说明水生植物营养生长对总酚的合成有着一定的促进作用,而C/N比率与总酚无显著相关性。N与总酚正相关的结果与Haukioja等[14]和Lima等[15]研究陆生植物得出的结果呈现相反的趋势。相比于水体环境,陆地环境中草食动物的种类(大型食草动物、鸟类和昆虫等)和数量更多,因此陆生植物面临的捕食压力更大。相关研究表明高浓度的酚类化合物可以在贫瘠的陆地生态系统中降低植物养分的流失[28,29],另外高酚以及低浓度的营养可以减少食草性动物对陆生植物的取食[30—33],此外,Lincoln等[34]指出酚类物质对某些食草性动物是有害的,Haukioja等[14]也研究得出同一区域内,毛枝桦叶片中总酚含量越高,食草性动物越少。此外,在森林生态系统中,酚类会降低土壤N循环速率,从而影响植物对N的利用[35],这些可能是陆生植物表现出N和总酚负相关的原因。
Carbon:Nutrient平衡(CNB)假说[36]认为,酚类物质的产生与植物内部生长所需要以及净光合产量之间的C供给成正比关系,本研究结果也从侧面印证了这一假说。由于植物快速生长(吸收大量含N营养)与光合作用效率(合成含C化合物)密切相关,因而在Carbon:Nutrients平衡假说的前提下,C和N分别与总酚表现出正相关性。
洱海属于高原封闭湖泊,年光照充足,加上样品采集区域水体营养盐含量较高,这些因素都会对不同生活型水生植物体内总酚含量产生不同程度的影响,考虑到N与总酚正相关,在富营养化进程中植物将因增加N供给而使酚类含量增加,此外富营养化使沉水植物衰退,挺水植物和浮叶植物的比例相应增加,这两者结合将使动物的牧食减少,凋落物难降解而累积,从而可能促进湖泊的沼泽化进程。
4 小结
不同生活型水生植物处于不同的环境中,迫使其朝着不同的方向进化发展,同时这种差异也体现在元素和次生代谢产物组成上,而富营养化的条件对其在不同生活型水生植物中的分布可能产生一定影响。对不同环境的适应也使得水生植物生理上产生了不同于陆生植物的规律,如总酚和N之间关系等。C、N和酚在水体生态系统的富营养化和沼泽化进程中的意义以及酚类物质对湖泊生态系统中物质循环的影响尚需进一步的研究。
致谢:
感谢中国科学院水生生物研究所的朱天顺、易春龙、李威、何亮、汤鑫、吴耀等师兄以及摆晓虎和王欢等同学在工作中给予的帮助,感谢洱海研究基地的刘杰研究员和龙德火工程师在样品采集中给予的帮助。
[1]Cao T. Study on the impacts ofNH+4enrichment on biochemistry,growth and competition of aquatic macrophytes [D]. Thesis for Doctor of Science. Institute of Hydrobiology,Chinese Academy of Sciences,Wuhan. 2006[曹特. 铵态氮富集对水生高等植物生化、生长和竞争胁迫研究. 博士学位论文,中国科学院水生生物研究所,武汉. 2006]
[2]Sculthorpe C D. The Biology of Aquatic Vascular Plants[M]. London:Edward Arnold. 1967,610—637
[3]Li D L,Wang L,Ding J J,et al. Ecological functions and resource utilization of aquatic plants [J]. Wetland Science,2011,9(3):290—296 [李冬林,王磊,丁晶晶,等. 水生植物的生态功能和资源应用. 湿地科学,2011,9(3):290—296]
[4]Wetzel R G. Structure and Productivity of Aquatic Ecosystems [M]. In:Wetzel R G(Eds.),Limnology(2ed). New York:CBS College Publishing. 1983,134—156,519—590
[5]Bowes G. Pathways of CO2Fixation by Aquatic Organisms [M]. In:Lucas W J,Berry J A(Eds.),InorganicCarbon Uptake by Aquatic Photosynthetic Organisms. Rockville,Maryland:American Society of Plant Physiologists. 1985,187—210
[6]Shi R H,Niu Z,Zhuang D F. Feasibility of estimating leaf C/N ratio with hyperspectral remote sensing Data [J]. Remote Sensing Technology and Application,2003,18(2):76—80 [施润和,牛铮,庄大方. 利用高光谱数据估测植物叶片碳氮比的可行性研究. 遥感技术与应用. 2003,18(2):76—80]
[7]Smolders A J P,Vergeer L H T,van der Velde G,et al. Phenolic contents of submerged,emergent and floating leaves of aquatic and semi-aquatic macrophyte species:why do they differ [J]? Oikos,2000,91(2):307—310
[8]Cao T,Xie P,Ni L Y,et al. Relationships among the contents of total phenolics,soluble carbohyd rate,and free amino acids of 15 aquatic macrophytes [J]. Journal of Freshwater Ecology,2008,23(2):291—296
[9]Lu M,Yuan B,Zeng M M,et al. Antioxidant capacity and major phenolic compounds of spices commonly consumed in China [J]. Food Research International,2011,44(2):530—536
[10]Mussatto S,Ballesteros L,Martins S,et al. Extraction of antioxidant phenolic compounds from spent coffee grounds [J]. Separation & Purification Technology,2011,83(22):173—179
[11]Pajak P,Socha R,Galkowska D,et al. Phenolic profile and antioxidant activity in selected seeds and sprouts [J]. Food Chemistry,2014,143(1):300—306
[12]Han S Q. Mechanism of phenolics on resisting fusarium wilt of banana [D]. Thesis for Master of Science. The Huazhong Agricultural University,Wuhan. 2014 [韩树全. 酚类物质抗香蕉枯萎病的作用机理. 硕士学位论文,华中农业大学,武汉. 2014]
[13]Cheng C L,Li J Q. Research advances in ecological significance and quantification of plant Polyphenols [J]. Chinese Journal of Applied Ecology,2006,17(12):2457—2460 [程春龙,李俊清. 植物多酚的定量分析方法和生态作用研究进展. 应用生态学报,2006,17(12):2457—2460]
[14]Haukioja E,Niemelä P,Sirén S. Foliage phenols and nitrogen in relation to growth,insect damage,and ability to recover after defoliation,in the mountain birch Betula pubescens ssp tortuosa [J]. Oecologia(Berlin),1985,65(2):214—222
[15]Lima A L S,Zanella F,Schiavinato M A,et al. Nitrogenous compounds,phenolic compounds and morphological aspects of leaves:comparison of deciduous and semideciduous arboreal legumes [J]. Scientia Agricola,2006,63(1):40—45
[16]Zhang H Y,Cai Q H,Tang T,et al. Comprehensive assessment and comparison of lakes' ecosystem health in Erhai watershed [J]. China Environmental Science,2012,32(4):715—720 [张红叶,蔡庆华,唐涛,等. 洱海流域湖泊生态系统健康综合评价与比较. 中国环境科学,2012,32(4):715—720]
[17]Li Z K,Yang P P,Su S F. Eutrophication trend of Erhai lake from 2004 to 2013 [J]. Environmental Science Survey,2015,34(1):1—3 [李泽坤,杨萍萍,苏社飞. 2004—2013年洱海富营养化变化趋势. 环境科学导刊,2015,34(1):1—3]
[18]Cheng J L,Hu M M,Zhou H D,et al. Studies on population dynamics and the underlying impact factors of phytoplankton during the cyanobacteria bloom in lake Erhai [J]. Acta Hydrobiologica Sinica,2015,39(1):24—28 [陈建良,胡明明,周怀东,等. 洱海蓝藻水华爆发期浮游植物群落变化及影响因素. 水生生物学报,2015,39(1):24—28]
[19]Mole S,Waterman P G. A critical analysis of techniques for measuring tannins in ecological studies [J]. Oecologia,1987,72(1):137—147
[20]Huang L,Wu Y,Zhang J,et al. Distribution of C、N、P and δ13C in aquatic plants of some lakesin the middle Yangtze Valley [J]. Acta Geoscientia Sinica,2003,24(6):515—518 [黄亮,吴莹,张经,等. 长江中游若干湖泊水生植物体内C、N、P及δ13C分布. 地球学报,2003,24(6):515—518]
[21]Wu A P,Wu S K,Ni L Y. Study of macrophytes nitrogen and phosphorus contents of the shallow lakes in the middle reaches of the Yangtze River [J]. Acta Hydrobiologica Sinica,2005,29(4):406—412 [吴爱平,吴世凯,倪乐意. 长江中游浅水湖泊水生植物氮磷含量与水柱营养的关系. 水生生物学报,2005,29(4):406—412]
[22]Lodge D M. Herbivory on freshwater macrophytes [J]. Aquatic Botany,1991,41(1—3):195—224
[23]Cronin G,Lodge D M. Effects of light and nutrient availability on the growth,allocation,carbon/nitrogen balance,phenolic chemistry,and resistance to herbivory of two freshwater macrophytes [J]. Oecologia,2003,137(1):32—41
[24]Dudt J F,Shure D J. The influence of light and nutrients on foliar phenolics and insect herbivory [J]. Ecology,1994,75(1):86—98
[25]Lodge D M,Cronin G,Donk E V,et al. Impact of herbivory on plant standing crop:comparisons among biomes,between vascular and nonvascular plants,and among freshwater herbivore taxa [A]. In:Jeppesen E,Søndergaard M,Christoffersen K(Eds.),The Structuring Role of Submerged Macrophytes in Lakes [C]. New York:Springer. 1998,149—174
[26]Søndergaard M,Lauridsen T L,Jeppesen E,Bruun L. Macrophyte-waterfowl interactions:tracking a variable resource and impact of herbivory on plant growth [A]. In:Jeppesen E,Søndergaard M,Christoffersen K(Eds.),The Structuring Role of Submerged Macrophytes in Lakes[C]. New York:Springer. 1998,298—306
[27]Ni L Y. Effects of apex cutting on growth of Elodeacanadensis St.John [J]. Acta Hydrobiologica Sinica,1999,23(4):297—303 [倪乐意. 切除顶枝对加拿大伊乐藻生长的影响. 水生生物学报,1999,23(4):297—303]
[28]Hättenschwiler S,Vitousek P M. The role of polyphenols in terrestrial ecosystem nutrient cycling [J]. Trends in Ecology & Evolution,2000,15(6):238—243
[29]Yu Z,Dahlgren R A,Northup R R. Evolution of soil properties sand plant communities along an extreme edaphic gradient [J]. European Journal of Soil Biology,1999,35(1):31—38
[30]Rhoades D F,Cates R G. Toward a general theory of plant antiherbivore chemistry [A]. In:Wallace J W,Mansell R L(Eds.),Biochemical Interaction Between Plants and Insects [C]. New York:Springer US. 1976,168—213
[31]Moran N,Hamilton W D. Low nutritive quality as defence against herbivores [J]. Journal of Theoretical Biology,1980,86(2):247—254
[32]Neuvonen S,Haukioja E. Low nutritive quality as defence against herbivores:induced responses in birch [J]. Oecologia(Berlin),1984,63(1):71—74
[33]Cooper P,Smith M,Maynard H. The design and performance of a nitrifying vertical-flow reed bed treatment system [J]. Water Science and Technology,1997,35(5):215—221
[34]Lincoln D E,Newton T S,Ehrlich P R,et al. Coevolution of the checkerspot butterfly Euphydryas chalcedona and its larval food plant Diplacus aurantiacus:larval response to protein and leaf resin [J]. Oecologia(Berlin),1982,52(2):216—223
[35]Palm C A. Contribution of agroforestry trees to nutrient requirements of intercropped plants [J]. Agroforestry Systems,1995,30(1—2):105—124
[36]Bryant J P,Chapin F S,Klein D R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory[J]. Oikos,1983,40(3):357—368
CONTENTS OF C,N AND TOTAL PHENOLS IN LEAVES OF AQUATIC MACROPHYTES IN LAKE ERHAI,CHINA
YUAN Chang-Bo1,2,CAO Te2,ZHOU Cun-Yu1,2,NI Le-Yi2and ZHANG Xiao-Lin2
(1. Yangtezu University,Jingzhou 434025,China; 2. Donghu Experimental Station of Lake Ecosystem,Institute of Hydrobiology,Chinese Academy of Sciences,Wuhan 430072,China)
Carbon(C),nitrogen(N) and total phenols(TP) in plants play important roles in palatability for herbivores and litter decomposition of the plant tissues,which impact C and N cascading among food web and biogeochemical recycling in lake ecosystem. In this study,concentrations of C,N and TP were examined in leaves of 26 aquatic macrophytes(emergent,floating-leaved and submersed) collected in Lake Erhai. Results showed that the mean values of leaf C,N and TP concentrations were 448.1,39.3 and 24.7 mg/g for emergent macrophytes(EM),433.3,35.2 and 28.1 mg/g for floating-leaved macrophytes(FM),378.4,27.3 and 10.3 mg/g for submersed macrophytes(SM),respectively. Leaf C and N concentrations decreased in the order of EM,FM and SM. EM and FM have similar TP concentration,which were higher than that of SM. The correlation analysis found significant positive correlation between leaf C,N concentrations and leaf TP concentrations for all 26 aquatic macrophytes. Light intensity and CO2availability for three kinds of macrophytes were significantly different,which might affect leaf C,N and TP concentrations. In addition,external stresses(herbivorous insects,pathogens,UV radiation,etc) and different growth strategies might also explain above findings. These results suggest that Potamogeton maackianus and Potamogeton lucens can be used as pioneer species for the recovery of Lake Erhai. The positive correlation between N and TP in leaves of macrophytes may promote the acceleration of lake paludification in the process of eutrophication.
Lake Erhai; Aquatic macrophytes; Carbon; Nitrogen; Total phenolic
Q178.1
A
1000-3207(2016)05-1025-08
10.7541/2016.133
2015-09-14;
2016-01-10
国家水体污染控制与治理科技重大专项项目(2012ZX07105-004); 淡水生态与生物技术国家重点实验室开放基金(2015FB01);国家自然科学基金项目(31200356)资助 [Supported by the National High Technology Research and Development Program of China(2012ZX07105-004); State Key Laboratory of Freshwater Ecology and Biotechnology(2015FB01); the National Natural Science Foundation of China(31200356)]
袁昌波(1990— ),男,湖北孝感人; 博士研究生; 主要从事水生植物生理生态学研究。E-mail:836207053@qq.com
周存宇(1968— ),男,湖北黄石人; 副教授; 主要从事水生植物生态学研究。E-mail:zhoucy@yangtzeu.edu.cn
