四川米仓山自然保护区台湾水青冈种群生命表及动态分析
2016-11-10李金昕吴定军章世鹏何显湘史清茂
李金昕 吴定军 章世鹏 何显湘 陈 坚 史清茂 胥 晓*
(1.西华师范大学生命科学学院,南充 637009; 2.四川省旺苍县米仓山自然保护区管理局,旺苍 628200)
四川米仓山自然保护区台湾水青冈种群生命表及动态分析
李金昕1吴定军2章世鹏2何显湘2陈 坚2史清茂2胥 晓1*
(1.西华师范大学生命科学学院,南充 637009;2.四川省旺苍县米仓山自然保护区管理局,旺苍 628200)
台湾水青冈(Fagushayatae)为水青冈属植物,仅分布于中国,且是国家Ⅱ级保护植物。了解台湾水青冈的种群的龄级结构及数量对于揭示种群动态和生活史特征有重要意义。本文通过对四川米仓山自然保护区老林沟北坡台湾水青冈种群样地进行调查,根据树木径级大小划分龄级,采用匀滑技术,编制其种群生命表,并进行生存分析和谱分析。结果表明:台湾水青冈种群幼苗、幼树和小树占总数的52%,中龄级个体占总数的46%,老树较少,种群年龄结构为稳定型,种群处于成熟阶段,种群更替无障碍。死亡率在第1龄级呈最大值,在第17龄级呈峰值。生存率下降趋势总体较平滑但中间出现2次大的降幅,累计死亡率则相反。谱分析结果显示,台湾水青冈种群更新过程存在着明显的周期性。振幅Ak最大的2个值分别位于基波A1和谐波A2处,显示了台湾水青冈个体生命周期的时间长度。台湾水青冈种群数量动态除受基波A1和谐波A2影响外,还表现出特别明显的小周期波动,如A3和A7。A3处的波动与台湾水青冈的高生长有关,而A7的波动与径向生长期的激烈竞争有关。我们的研究揭示了该地台湾水青冈种群数量动态变化趋势符合Deevey Ⅲ型,种群动态具有前期锐减、中期稳定和后期衰退的特点。
生存分析;种群数量动态;台湾水青冈;谱分析
种群的数量动态变化研究一直是种群生态学的重要内容,其基本的特征往往通过编制种群生命表来予以定量化。根据种群生命表所提供的数据,能获得存活率、死亡率、内禀增长率等种群特征参数和关键信息[1~3],从而以此通过生存分析函数可以对种群生存现状和预计变化趋势进行分析[4~5]。此外,谱分析法作为分析事物周期性性变化的方法,可以用来揭示种群数量分布的周期性波动,是研究林分分布波动和周期性年龄更替过程的工具[6]。因此,种群生命表和谱分析法对研究种群数量动态和生存状况具有重要的参考价值。
台湾水青冈(Fagushayatae)为我国特有植物,属国家Ⅱ级保护植物,主要分布于台湾、四川、浙江和湖北等省,其种群分布海拔范围各不相同。如在台湾可达到2 300 m,在四川分布范围为1 200~2 000 m,在浙江四海山分布海拔仅650 m左右[7]。从查阅的文献来看,国内外对台湾水青冈的研究主要集中在药效成分、分子遗传、群落特征等方面。如在药用成分方面,Lai等[8]发现台湾水青冈的嫩枝和绿叶的提取液中的三萜类化合物对α葡萄糖苷酶有抑制作用。在分子遗传方面,Ju等[9]使用微卫星引物对台湾水青冈进行了研究并指出部分多态位点可以用于遗传多样性研究。Denk等[10]根据DNA分子片段和形态学特征调查了水青冈属植物的系统发生学关系,并确定台湾水青冈为水青冈属的基位种。宋文静等[11]对台湾水青冈的ISSR-PCR体系进行了优化。在群落特征方面,Hukusima等[12]发现台湾地区的台湾水青冈群落同邻近的常绿阔叶林拥有较少的共有种,并认为该地区的台湾水青冈林为残次林。张方钢[13]对浙江清凉峰台湾水青冈群落的群落结构和变化作了调查,认为该地群落纯度高,稳定性强,且保存完好,是该地区中山地带落叶阔叶林的重要植被类型之一。何俊等[14]研究了七姊妹山的台湾水青冈群落,发现该地群落结构呈倒金字塔形,属于衰退型群落。
综上,从种群数量特征方面对台湾水青冈的研究工作相对较少(仅见于郭瑞[15]等对浙江清凉峰台湾水青冈种群的更新研究),尤其是在种群动态和生存分析方面的研究少见报道。由于种群的数量动态直接决定着种群的存活能力,其变化规律对掌握种群的龄级结构、种群动态和生活史的特征具有重要意义。旺苍县米仓山国家级自然保护区现存大量的台湾水青冈,成片分布区域较多[16]。这地台湾水青冈林具有分布广,林龄高,成树多的特点,非常适合全面调查台湾水青冈的生活周期,是台湾水青冈种群特征研究的最佳样地。因此,本文以种群生命表及生存分析理论为基础,对米仓山自然保护区的台湾水青冈种群统计特征进行了初步分析,以期望揭示台湾水青冈的种群动态和种群生活史特征,为保护台湾水青冈提供理论支撑。
1 研究区域概况
四川米仓山自然保护区处于秦岭山系的余脉,区内大部分区域海拔1 500~2 000 m。山地层状地貌在该区相当明显。该区位于亚热带湿润季风气候,受特殊的山地地貌影响,降雨充足,光照时间久,全年无霜期较长。根据其所处于的旺苍县气象局的记录资料,该地全年平均气温16.1℃,历年最高气温39.0℃,最低-7.2℃;全年无霜期260 d,年均降雨1 200 mm,历年最多降雨量为2 092.4 mm,最少728.8 mm,年平均日照时数为1 355.3 h。区内的植物区系是中国——日本森林植物区的华中地区,以及华北地区的黄土高原亚地区与中国——喜马拉雅森林植物区的横断山脉地区交汇地带。米仓山保护区内有维管束植物195科949属2 597种,其中蕨类植物32科73属213种;裸子植物8科21属43种;被子植物155科853属2 341种。植物科占全国植物科的55.24%,属占全国植物属的29.86%,种占9.57%[17]。本次研究样地设于保护区内的老林沟北坡,该处台湾水青冈保存完好,是研究台湾水青冈种群的最佳区域。该地土层较薄,上层为腐殖质土,在部分区域能看到裸露的岩层。
2 研究方法
2.1 野外调查方法
在四川米仓山自然保护区老林沟选择台湾水青冈数量较为集中的森林群落为研究对象,在全面勘察的基础上,对老林沟北坡的台湾水青冈种群进行调查研究。由于该地属于沟谷地貌,坡度较大且地形和坡向变化大,设置大约200 m×200 m的近似方形区域为样地。在样地内采用群落常规调查方法对树高≤2 m的台湾水青冈记录树高、基径,对树高>2 m的台湾水青冈在树身1.3 m处测量其胸径并记录树高、冠幅。对于树身在1.3 m处产生分株的个体,将每一棵分株记作单独的个体,分别记录各指标。
2.2 年龄结构分级方法
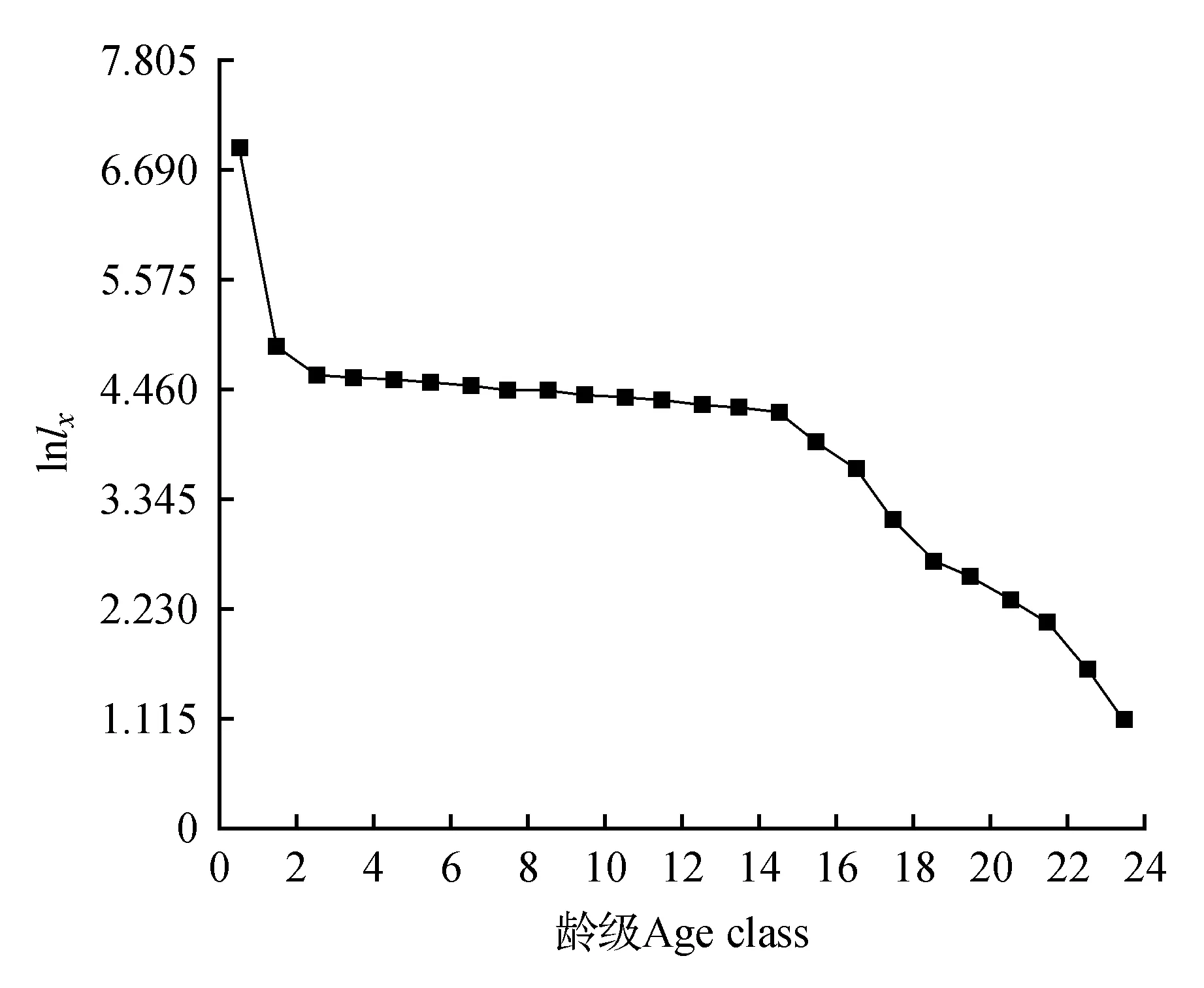
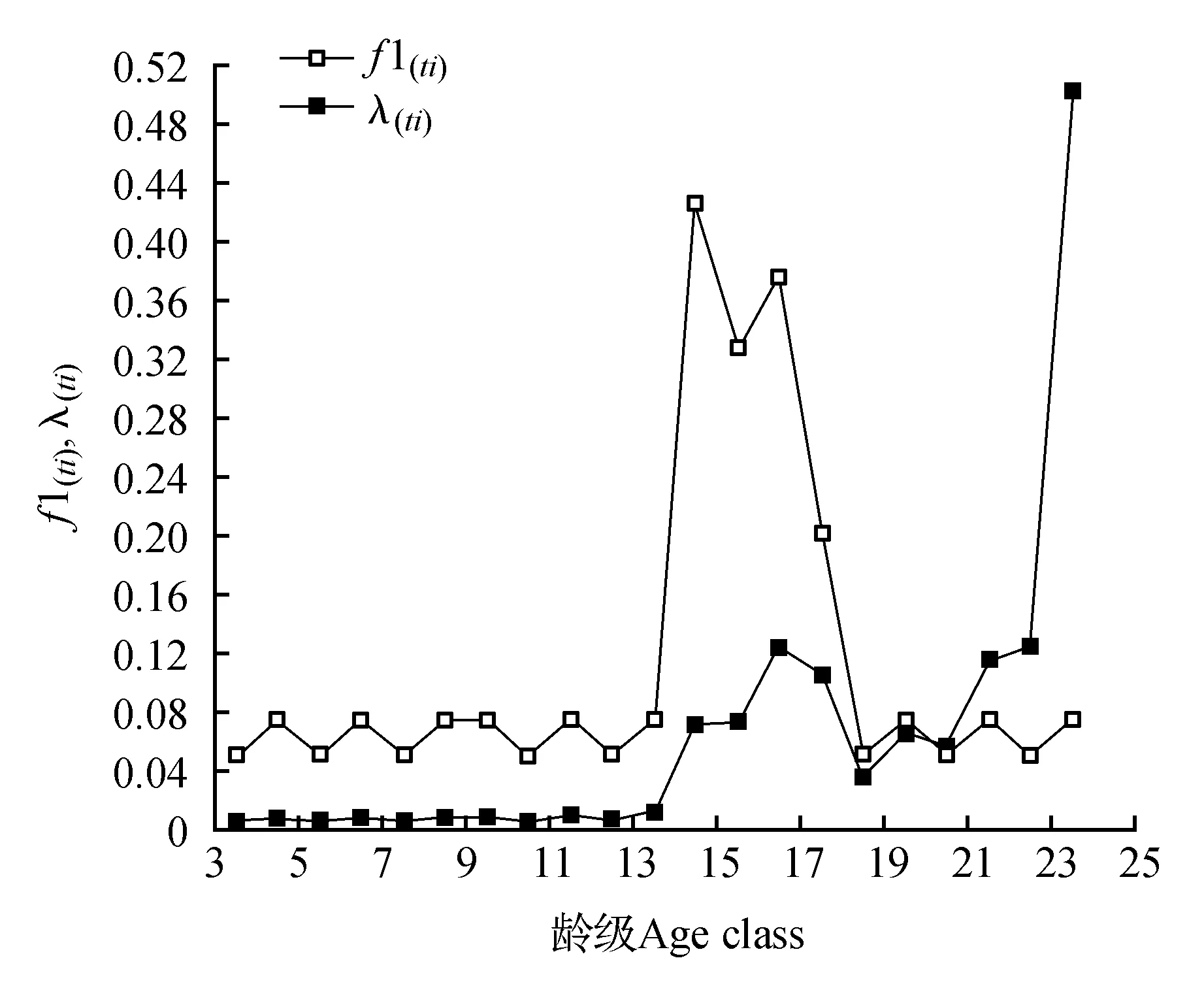
年龄结构是种群的重要要统计学参数,根据种群的年龄结构,可以知道种群过去及现在的更新情况[18]。由于台湾水青冈为国家二级重点保护植物,测定具体年龄会损伤树木,对其以年龄分级较为困难。许多学者将林木依胸径大小分级,取得了较好效果[19~22]。关于种群龄级的划分方法,许多学者提出了不同的策略[23~24]。因此,我们参照前人方法综合考虑,按树高和胸径大小将台湾水青冈种群分为24级,分别是H≤50 cm、50 cm 2.3 生命表编制与生存分析 生命表的编制方法参见孙儒泳等[25]。生存分析函数主要包括:生存率函数S(i)、积累死亡率函数F(i)、死亡密度函数f(ti)、危险率函数λ(ti)。结合静态生命表的结果,估算公式如下: S(i)=S1·S2·S3…S (1) F(i)=1-S(i) (2) fti=(Si-1-S(i))/hi (3) λ(ti)=2(1-Si)/[hi(1+Si)] (4) 其中,hi为龄级宽度。根据生存函数的估算值,绘制相应曲线。 2.4 匀滑技术 2.5 谱分析方法 谱分析是由傅立叶级数的变换得到的,具体方法参见伍业钢等的方法[6]。 3.1 台湾水青冈种群生命表 根据调查资料和龄级结构的划分标准,编制了台湾水青冈种群静态生命表(表1)。从总体上说,台湾水青冈种群结构呈金字塔型,且存在波动性。幼苗、幼树和小树占总数的52%,第4龄级到第17龄级的个体数量占总数的46%,老龄个体数量相对较少,整个种群处于成熟阶段。除第1和第2龄级阶段外,种群在第4和第15龄级出现了个体数量高峰。从第5龄级到第13龄级,个体数量相差不大。18龄级后,个体数量非常稀少。种群个体的生命期望由ex表示,计算结果表明,台湾水青冈个体在第2龄级至第11龄级阶段具有很高的期望寿命。 3.2 存活曲线分析 由存活曲线图1可以得出,台湾水青冈各龄级的存活数在幼苗阶段下降最快。从第3阶段起,台湾水青冈的死亡率降至整个种群的最低水平,该情况一直持续到第15龄级。15龄级后,种群的死亡个体比例明显上升。大约75%的第15龄级台湾水青冈能存活生长至第16龄级。存活率在第16至18龄级较低,在第19龄级时出现明显升高。19龄级以后其又开始逐渐下降且下降速度加快,该情况一直持续到最大龄级。存活曲线一般分为3种类型,即凸线型(Deevey Ⅰ),直线型(Deevey Ⅱ)和凹线型(Deevey Ⅲ)。该种群在幼苗阶段,存活数远多于其他阶段且生存率最低。因此,台湾水青冈种群的存活曲线应为Deevey Ⅲ型。 表1 台湾水青冈种群静态生命表 注:ax.存活数;lx.存活量;dx.死亡量;qx.死亡率;Lx.区间寿命;Tx.总寿命;ex.期望寿命;Kx.消失率;Sx.存活率 Note:ax.Survival number;lx.Survival quantity;dx.Death number;qx.Mortality rate;Lx.Span life;Tx.Total life;ex.Life expectancy;Kx.Vanish rate;Sx.Survival rate 图1 台湾水青冈种群的存活曲线Fig.1 Survival curve of F.hayatae populaiton 由公式换算关系可知,死亡率和消失率的变化趋势基本相同,且死亡率越小二者的差也越小。从图2可知,死亡率和消失率均存在两个特征值:第一个特征值为最大值,发生在第1龄级,显示死亡率为86.6%,消失率为2.01;第2个特征值为极大值,发生在第17龄级,显示死亡率为39.5%,消失率为0.503。第24龄级的死亡率为1,这是由于第25龄级个体数的观测值为0。根据实地调查情况,这可能是由于调查的树木数量少,林龄短导致的。因此,1是对第24龄级死亡率的有偏估计,不能反映台湾水青冈种群的真实的状况,但仍能提供有用的信息。 图2 台湾水青冈种群死亡率(qx)和消失率曲线(kx)Fig.2 Mortality rate(qx) and vanish rate(kx) value curve of F.hayatae population 图3 台湾水青冈种群生存率(S(i))和累计死亡率(F(i))曲线Fig.3 Survival rate(S(i)) and cumulative mortality rate(F(i)) functional curve of F.hayatae population 图4 台湾水青冈种群相对死亡密度(f1(ti))和危险率(λ(ti))曲线Fig.4 Relative mortality density(f1(ti)) and hazard rate(λ(ti)) functional curve of F.hayatae population 3.3 生存分析 根据种群的演替特性,种群生存率总是单调下降,而累计死亡率则是单调上升;根据公式变换可知,生存率和累计死亡率的和为1,且两者变化趋势的大小互为相反数。由图3可知,种群生存率曲线和累计死亡率曲线都在第2龄级、第14龄级出现2次明显的转折,分别使曲线变化速度减慢和加快。到第18龄级以后,种群个体数量明显大幅减少,累计死亡率超过98%。 为了更方便的表达死亡密度曲线,我们对死亡密度函数乘以100,得到相对死亡密度函数f1(ti)。f1(ti)与f(ti)有相同的变化速度。由图4可知,台湾水青冈种群的死亡密度和危险率变化趋势基本一致,都在第17龄级出现极大值。4个生存函数表明,台湾水青冈具有前期锐减、中期稳定、后期衰退的特点。 表2 生存分析函数估计值 注:S(i).生存率函数;F(i).累计死亡率函数;f(ti).死亡密度函数;λ(ti).危险率函数 Note:S(i).Survival rate function;F(i).Cumulative mortality rate function;f(ti).Mortality density function;λ(ti).Hazard rate function 3.4 谱分析 由表3可知,在振幅Ak值中基波A1=1.30和谐波A2=1.11为最大的2个值。这表明台湾水青冈种群不仅表现出明显的基本周期,而且基本周期还不稳定。基本周期反映了台湾水青冈种群的生物学特征,显示了台湾水青冈个体生命周期的时间长度。从表3还可看出,台湾水青冈种群动态也表现出较短的周期波动,如A3、A7。A3处的波动与台湾水青冈的高生长有关,此处波动约在空间序列径级小于10 cm处,对应于第5龄级;A7的波动约在空间序列径级40~44 cm处,对应于第13龄级,A7的波动与径向生长期的激烈竞争有关。台湾水青冈种群数量动态的谱分析表明,台湾水青冈种群数量动态的波动性不是由单一周期表现出来的,而是多个周期的共同作用的结果。种群周期性的波动也和种群所处生境有关,同时也符合稳定植物群落中优势种群自我更新的特点。 表3 台湾水青冈种群的周期性波动 了解台湾水青冈生长和天然更新过程所受到的各种作用,对于保护台湾水青冈有重大的意义。分析结果表明,台湾水青冈种群的存活曲线为凹线型。台湾水青冈种群在第1龄级、第2龄级出现了死亡高峰。植物有机体作为复杂群落中的一部分,其生长受到生物因素和非生物因素的双重影响。样地调查结果显示:台湾水青冈在幼苗和幼树阶段呈明显的聚集分布。这可能与其生物生态学特性有关[27~28]。台湾水青冈依靠种子繁殖,无萌生苗。其果实为坚果,种子无翅,散布距离一般不远。另外,该处的沟谷地形也加重了种子的散布困难。随着台湾水青冈群落的演替发展,幼苗对营养和光照的需求不断增加,与邻近个体和上层乔木以及下层灌木间的生态位重叠不断加大,生存环境变得困难的个体开始增多。当林内的养分、光照无法满足其生长所需的临界点时,死亡率提高,自疏作用出现并逐渐加强。从第3龄级到17龄级,台湾水青冈种群个体的死亡率明显较低,这表明个体在该区间竞争压力小,自疏作用较弱。这可能是由于台湾水青冈种群种群密度较低,种群规模未达到环境容纳量上限。17龄级后,台湾水青冈死亡率升高。这可能是由于潮湿陡峭的山地,强烈的季节风等增加了老树毁坏的范围[29]。调查结果也发现,掘根倒木和主干折断是导致大部分台湾水青冈成体死亡的直接原因,其中倒木所占的比例明显较大。 动物行为对处于某些生活史阶段的台湾水青冈有重要作用。影响地面种子库中种子命运的主要因素为动物尤其是小型啮齿动物的捕食和搬运作用[30]。壳斗科植物的果实包含丰富的蛋白质、碳水化合物和脂肪,是鸟类和小型兽类优良的食物来源[31]。刘映良等[30]在鼠类对水青冈种子命运影响的研究中发现,鼠类转运和就地消耗是影响水青冈种子命运的主要因素。因此,我们也认为啮齿类动物的采食行为对台湾水青冈种子的散布有潜在的影响。人为活动对于台湾水青冈的保护和种群的发展都有影响。在调查区域,我们发现了居民采药而对水青冈幼苗造成践踏的痕迹。这对台湾水青冈种群的更新产生了消极影响。米仓山台湾水青冈早年遭受砍伐较严重,现有面积较小,且多为零星分布,像本样地中的台湾水青冈纯林已经相当少[16]。保持一定规模的种群可以有效消除种群随机灭绝的风险。但台湾水青冈群落周围的林分植被覆盖良好且郁闭度高,通过天然更替的方式扩大台湾水青冈种群规模较为困难。 生存分析函数是研究种群动态的有力工具。分析结果表明,随着龄级的增大,生存率逐渐接近0,累计死亡率逐渐接近1,它们是同一种变化过程的两种描述形式。死亡密度曲线与危险率曲线的变化形式基本一致,但在某些阶段差别较大。这是因为死亡密度曲线以龄级内死亡个体数量为参照,而危险率曲线以龄级内死亡个体比例为参照。由观察可知,从19到24龄级,每龄级内个体死亡数为1,此时种群死亡密度基本不变,而危险率则显著升高。该差异表明,种群个体数较少时,种群的随机性会增加,此时生命表数据有可能无法准确的反映种群个体的真实生存能力,个体的危险率有可能被高估。谱分析能应用于种群动态的周期性波动研究中。谱分析结果表明,台湾水青冈种群存在明显的周期性,且基波A1的波幅最大。这表明台湾水青冈种群的数量动态明显受台湾水青冈的生命周期特性控制。但可能由于所调查的台湾水青冈种群密度低,年龄小,最大径级84~88 cm,时间系列还不够长,不能完整表现出基本周期,林分还未进入成熟阶段,不能表现出稳定的固有周期。 1.李晓笑,王清春,崔国发,等.濒危植物梵净山冷杉野生种群结构及动态特征[J].西北植物学报,2011,31(7):1479-1486. 2.韩路,王海珍,彭杰,等.塔里木河上游灰胡杨种群生存分析[J].林业科学,2010,46(1):131-135. 3.李清河,高婷婷,刘建锋,等.荒漠珍稀灌木半日花种群的年龄结构与生命表分析[J].植物研究,2009,29(2):176-181. 4.郑维列,田大伦,卢杰,等.高寒植物长鞭红景天种群结构及数量特征[J].植物研究,2009,29(4):402-410. 5.刘贵峰,臧润国,成克武,等.天山云杉的种群动态[J].应用与环境生物学报,2011,17(5):632-637. 6.伍业钢,薛进轩.阔叶红松林红松种群动态的谱分析[J].生态学杂志,1988,7(1):19-23. 7.李明华.清凉峰自然保护区志[M].长春:吉林人民出版社,2006:1-67. 8.Lai Y C,Chen C K,Tsai S F,et al.Triterpenes as α-glucosidase inhibitors fromFagushayatae[J].Phytochemistry,2012,74:206-211. 9.Ju L P,Shih H C,Chiang Y C.Microsatellite primers for the endangered beech tree,Fagushayatae(Fagaceae)[J].American Journal of Botany,2012:e000-e000. 10.Denk T,Grimm G W,Hemleben V.Patterns of molecular and morphological differentiation inFagus(Fagaceae):phylogenetic implications[J].American Journal of Botany,2005,92(6):1006-1016. 11.宋文静,金则新,李钧敏,等.台湾水青冈ISSR-PCR体系的优化[J].福建林业科技,2009,36(2):18-22. 12.Hukusima T,Lu S Y,Matsui T,et al.Phytosociology ofFagushayataeforests in Taiwan[J].Rendiconti Lincei,2005,16(3):171-189. 13.张方钢.浙江清凉峰台湾水青冈林的群落学特征[J].浙江大学学报:农业与生命科学版,2001,27(4):403-406. 14.何俊,汪正祥,雷耘,等.七姊妹山自然保护区台湾水青冈林群落学特征研究[J].华中师范大学学报:自然科学版,2008,42(2):272-277. 15.郭瑞,翁东明,金毅,等.浙江清凉峰台湾水青冈种群2006~2011年更新动态及其与生境的关系[J].广西植物,2014,34(4):478-483. 16.陈坚.米仓山自然保护区水青冈属(Fagus)资源调查报告[J].中国野生植物资源,2014,33(2):47-52. 17.吴征镒.中国植被[M].北京:科学出版社,1980. 18.Agren J,Zackrisson O.Age and size structure ofPinussylvestrispopulations on mires in central and northern Sweden[J].The Journal of Ecology,1990,1049-1062. 19.Harper J L.Population biology of plants[M].London:Academic Press,1977. 20.Rebertus A J,Veblen T T.Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego,Argentina[J].Journal of Vegetation Science,1993,4(5):641-654. 21.闫淑君,洪伟,吴承祯,等.丝栗栲种群生命过程及谱分析[J].应用与环境生物学报,2002,8(4):351-355. 22.刘任涛,毕润成,闫桂琴.山西南部翅果油树种群动态与谱分析[J].植物研究,2007,27(5):550-555. 23.江洪.云杉种群生态学[M].北京:中国林业出版社,1992. 24.何亚平,费世民,蒋俊明,等.不同龄级划分方法对种群生存分析的影响——以水灾迹地油松和华山松种群生存分析为例[J].植物生态学报,2008,32(2):448-455. 25.孙儒泳,李庆芬,牛翠娟,等.基础生态学[M].北京:高等教育出版社,2002:66-71. 26.Wratten S D,Fry G L.Field and laboratory exercises in ecology[M].London:Edward Arnold,UK,1980. 27.肖宜安,何平,李晓红,等.濒危植物长柄双花木自然种群数量动态[J].植物生态学报,2004,28(2):252-257. 28.张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156. 29.Pócs T.The epiphytic biomass and its effect on the water balance of two rain forest types in the Uluguru Mountains(Tanzania,East Africa)[AspleniumnidusL.,Microsoriumpunctatum(L.) Copel.,Corticolousmicroepiphytes,Mossyelfinforest][J].Acta Botanica Academiae Scientiarum Hungaricae,1980,26:143-167. 30.刘映良,薛建辉.鼠类及种子特征对水青冈种子命运的影响[J].南京林业大学学报:自然科学版,2010,34(5):12-16. 31.肖治术,王玉山,张知彬.都江堰地区三种壳斗科植物的种子库及其影响因素研究[J].生物多样性,2001,9(4):373-381. LifeTableandDynamicAnalysisofFagushayataePopulationinMicangshanNatureReserve,SichuanProvince,China LI Jin-Xin1WU Ding-Jun2ZHANG Shi-Peng2HE Xian-Xiang2CHEN Jian2SHI Qing-Mao2XU Xiao1* (1.College of Life Science,China West Normal University,Nanchong 637009;2.Micangshan Nature Reserve,Wangcang 628200) Fagushayatae, a rare and endangered plant in China(category II), is distributed in China. Understanding the age structure and number ofF.hayataepopulation is useful to reveal population dynamics and life-history traits. We investigatedF.hayataepopulation distributes on the north slope of Laolin Valley in Micangshan Nature Reserve, and obtained a static life table on the basis of age class structure by using the smoothing out technique. Then, we analyzed the population quantitative dynamics ofF.hayataeaccording to the results of survival and spectral analysis. The population was composed of 52% young individuals(including seedlings), 45.5% of mature individuals, and 2.5% of old individuals. Two peaks of higher mortality rate were in 1st and 17th age class periods of the lifespan. In addition to two big drops, the population survival rate decreased gradually from 1st to 24th age class. However, the opposite trend was found in cumulative mortality rate curve. By spectral analysis, the regeneration process of the population showed the marked periodic variations. The amplitudeAkhad the largest values at fundamental waveA1and harmonic waveA2reflecting the lifespan ofF.hayataeindividuals. The quantitative fluctuation of population exhibited obvious small cycles, such asA3andA7, which closely related with height growth and intense competitions during radial growth phase, respectively. Our results showed that the survival curve ofF.hayataepopulation in Micangshan Nature Reserve was agree with Deevey type Ⅲ, and the population dynamics exhibited a trend that population size decreased sharply at young stage, kept stably at mature stage, and declined at old stage. survival analysis;population quantitative dynamics;Fagushayatae;spectral analysis 四川米仓山自然保护区管理局委托项目(2014-02) 李金昕(1989—),男,硕士研究生,主要从事植物生态研究。 * 通信作者:E-mail:xuxiao_cwnu@163.com 2015-05-08 S718.5 A 10.7525/j.issn.1673-5102.2016.01.010

3 结果与分析





4 讨论
