2000—2010年长三角地区植被生物量及其空间分布特征
2016-11-02李广宇
李广宇,陈 爽,张 慧,张 童
(1.中国科学院南京地理与湖泊研究所,江苏 南京 210008;2.环境保护部南京环境科学研究所,江苏 南京 210042)
2000—2010年长三角地区植被生物量及其空间分布特征
李广宇1,2,陈 爽1①,张 慧2,张 童1
(1.中国科学院南京地理与湖泊研究所,江苏 南京 210008;2.环境保护部南京环境科学研究所,江苏 南京 210042)
植被生物量是陆地表层生态系统服务功能的重要表征,探明区域植被生物量的空间分布规律及其变化,对于制定合理的植被保护政策具有重要意义。当前针对生态系统总体的植被生物量空间估算研究较少且空间分辨率较低,难以反映土地利用/覆盖变化状况。以长三角地区16个地市为研究区,在遥感模型、GIS空间运算和数理统计分析等方法支持下,利用遥感、气象、土地利用/覆被、森林清查及粮食产量等多源数据,研究植被生物量空间分布及其变化。结果表明:植被生物量空间估算结果既保留了统计数据的总体精度,又反映了植被生物量空间分布特征;长三角地区植被生物量分布总体呈现南高北低的态势,沿长江地区和太湖流域生物量相对低。10 a间区域植被生物量总量显著上升,其中,江苏部分地市和上海植被生物量稳中有升,浙江部分地市植被生物量显著增长。用地类型保持不变的林地和耕地是生物量增加的主要来源,土地利用/覆被变化对植被生物量有显著影响,耕地、林地向建设用地转换导致植被生物量显著下降,建设用地绿化在一定程度上弥补了植被消失或受破坏引起的生物量损失。
生物量;空间估算;分布变化;长三角地区
植被生物量是地表碳循环的重要组成部分,是评价陆地生态系统结构与功能的重要指标[1-2]。准确定量估算植被生物量的空间分布是研究植被生物量时空变化及其受土地利用/覆被变化影响的基础[3],对研究区域碳平衡和评估陆地生态系统状况有重要意义[4-6]。当前植被生物量的空间估算多针对森林、草地等单一植被类型,空间分辨率较低且对整个生态系统的估算较少[7]。如刘双娜等[8]利用1∶100万森林植被图及同期净初级生产力(net primary productivity,NPP)空间分布结果估算了1 km分辨率条件下全国森林生物量;赵明伟等[9]结合1∶400万植被分布图模拟了全国森林植被生物量;朴世龙等[4]估算了8 km分辨率条件下全国草地植被生物量及其空间分布格局;TAO等[1]估算了8 km分辨率条件下全国玉米生物量的空间分布。方精云等[7]对全国林地、耕地、草地和灌丛等植被生物量总量进行估算,但未对其空间分布进行研究。
快速城市化地区土地利用/覆被变化剧烈,不透水面取代具有生态功能的自然和半自然植被景观[10-11],导致植被面积减小及景观破碎化加剧,出现了水环境恶化、城市热岛、生物多样性减少和生态服务功能降低等生态环境问题[12-17]。面对植被生物量损失引起的日趋严重的城市生态环境问题,城市化地区植被结构格局与功能的动态演化得到关注[5-6,18-19]。植被生物量可用以表征生态系统服务功能,但现有植被生物量空间估算结果的分辨率较低,且无法及时反映区域土地利用/覆被变化。基于高空间分辨率的土地利用/覆被数据,以中国最大的都市密集区——长江三角洲地区为研究区,研究城市化地区植被生物量空间分布及其变化,对区域碳循环研究、区域生态保护及土地利用政策制定具有重要意义。
1 研究区概况
研究区为国务院发布的《长江三角洲地区区域规划》中的长三角核心区(以下简称长三角地区),面积约110 800 km2,约占全国土地面积的1.1%,包括上海市和江苏省的南京、苏州、无锡、常州、镇江、扬州、泰州、南通,浙江省的杭州、宁波、湖州、嘉兴、绍兴、舟山、台州16个地市。2010年该核心区常住人口达1.07亿,GDP总量达7.1万亿,占全国总人口和GDP总量的比例分别达8.0%和17.8%。长三角地区城市空间急剧增长,形态过程变化复杂,给植被带来显著负面效应[20]。
区内自然分异特征明显,地貌以平原和丘陵为主。气候为亚热带季风气候,年均温为14~18 ℃,最冷月均温为0~5.5 ℃,最热月气温约为27~28 ℃。多年平均降水量为1 000~1 462 mm,受季风影响,降雨的年际与年内变化大,最大降雨发生在夏季。植被主要处于北亚热带常绿-落叶阔叶混交林带和中亚热带常绿阔叶林带2个植被地带,北部小部分区域为暖温带落叶阔叶林带[21]。
2 研究数据与方法
2.1 数据
采用的数据包括遥感数据、气象数据、土地利用/覆被数据、森林清查数据及粮食产量数据等。(1)遥感数据包括覆盖研究区的2000—2010年MOD13Q1数据产品,时间分辨率为16 d,空间分辨率为250 m;对影像进行镶嵌、格式转换和重投影处理,转换为Albers投影,采用最大合成法(maximum value composite,MVC)获取研究区逐月归一化植被指数(normalized difference vegetation index,NDVI)栅格数据。(2)气象数据包括2000、2010年长三角地区19个气象站点月平均温度、月降水量和6个气象站点的太阳辐射数据,数据来源于中国气象科学数据共享服务网(http:∥www.escience.gov.cn/);根据各气象站点的经纬度信息,对降雨、气温和太阳辐射气象数据进行Kriging空间插值,获取250 m分辨率的气象栅格数据。(3)土地利用/覆被数据包括2000年基于中国科学院土地资源分类系统的 1∶10万土地利用/覆被数据、20世纪80年代末期基于陆地生态系统特点的1∶25万土地利用/覆被数据(http:∥www.resdc.cn/)及2010年1∶10万面向生态系统碳收支服务的土地利用/覆被数据,3类数据分别来自中国资源环境科学数据中心(http:∥www.resdc.cn/)、地球系统科学数据共享网(http:∥www.geodata.cn/)和湖泊-流域科学数据共享平台(http:∥lwdc.niglas.cas.cn/)。建立基于生物量估算的土地利用/覆被分类体系,将土地利用/覆被类型分为耕地、林地、草地、水域、城镇建设用地和其他用地6大类,其中林地细分为常绿阔叶林、落叶阔叶林、常绿针叶林、落叶针叶林和灌丛。2010年土地利用/覆被数据可通过对原体系下的二级分类进行调整和归并,得到2010年基于生物量估算的土地利用/覆被分类。对于2000年土地利用/覆被数据,假定2000年原始数据在区域尺度上的总体数据分类精度为100%,将2000年和20世纪80年代末期土地覆被数据进行叠加,以20世纪80年代末期数据为参照,对2000年数据中的林地进行重分类,将两期均为林地的区域进行调整和归并,对于数据中不能准确判定用地类型的地块,结合1∶100万植被图、文献资料、对应年份的遥感Landsat TM/ETM影像和Google Earth影像进行辅助分类,获得2000年基于生物量估算的土地利用/覆被分类数据[22]。将2000、2010年土地覆被重分类数据转化为250 m×250 m栅格数据。(4)森林清查数据包括江苏省、浙江省和上海市第6次一类清查森林资源统计数据和第8次一类清查森林资源统计数据,分别来自中国林业科学研究院林业科学数据中心(http:∥www.cfsdc.org/)和中国林业网(http:∥211.167.243.162:8085/8/index.html)。(5)长三角地区粮食统计数据来自长三角地区16个地市2001、2011年统计年鉴。
2.2 植被生物量空间估算
准确且具时效性的植被生物量空间分布可以为植被保护和管理提供全面的依据。由于长三角地区浙江、上海和江苏的第8次森林清查数据时间分别为2009、2009和2010年,受森林清查数据的时间限制,为保持土地利用/覆被数据、森林清查数据等多源数据在时间上的匹配和一致性,研究时段定为2000—2010年。以研究区土地利用/覆被的空间分布为基础,综合遥感、气象、森林清查和粮食产量等多源数据,针对不同植被类型采用适合的模型方法进行估算,获得区域尺度植被生物量空间分布,进而揭示研究期内长三角地区植被生物量的空间分布及变化规律。
2.2.1 NPP空间估算
基于改进的CASA模型[5,23],利用土地利用/覆被数据、气象数据和MODIS NDVI遥感数据等,模拟2000、2010年长三角地区植被NPP的空间分布。
2.2.2 基于遥感降尺度技术的林地植被生物量空间估算
降尺度统计方法早期广泛应用于气候学研究,是一种将大尺度、低分辨率信息转化为区域尺度、高分辨率信息的方法[24]。基于森林生物量与NPP在大尺度上呈线性正相关的规律,综合森林清查数据和NPP空间分布图,可采用降尺度技术对区域尺度林地生物量空间分布进行估算[8,25]。
首先,采用森林资源清查数据中的乔木林优势树种面积蓄积统计数据,运用换算因子连续函数法[26],根据各优势树种生物量与蓄积量之间的回归方程,计算得到浙江、江苏和上海各优势树种的生物量,并将优势树种划入针叶林、阔叶林和针阔混交林等林型,计算不同林型的单位面积生物量。以土地利用/覆被图及NPP空间分布为纽带,采用降尺度技术,估算林地植被生物量,公式如下:

(1)
式(1)中,p为各省份;t为各森林植被类型(针叶林、针阔混交林和阔叶林);i为某一个栅格点;Bpt为某省某森林类型的平均生物量,Mg·hm-2;Ni为某栅格点的森林NPP(以C计),g·m-2·a-1;Bi为某栅格点森林生物量,Mg·hm-2;n为某省某森林类型的栅格点数。
通过文献调研得到江苏、浙江和上海灌丛植被单位面积生物量分别为12.34、33.14和12.34 t·hm-2[27],从土地利用/覆被数据中提取灌丛空间分布,对栅格像元的生物量赋值,得到灌丛植被生物量空间分布。将乔木林植被生物量空间分布图与灌丛植被生物量空间分布图进行镶嵌处理,得到林地植被生物量空间分布。
2.2.3 耕地植被生物量空间估算
耕地生物量估算方法主要包括基于遥感的生物量估算模型[28]和利用作物产量的统计数据估算[10]。基于2.2.1节方法估算得到的NPP空间分布,结合土地利用/覆被数据,提取耕地NPP空间分布信息。因模型模拟得到的NPP(以C计)单位是g·m-2·a-1,将其除以转换系数0.45,得到生物量空间分布[7]。尽管不同区域耕地的复种指数具有空间差异,该文假定耕地全年种植农作物,并以全年累积的NPP作为农作物生物量。
2.2.4 草地植被生物量估算
研究区草地面积较小,基于文献调研获取江苏、浙江和上海草地平均生物量分别为7.17、9.26和8.59 t·hm-2[4];从栅格化的土地利用/覆被数据中提取草地空间分布,分别对江苏、浙江和上海栅格像元的生物量赋值,获取草地植被生物量空间分布。
2.2.5 植被生物量空间分布图
在ArcGIS 10.0系统的支持下,以林地植被和耕地植被生物量空间分布图及草地植被生物量空间分布图为基础,将林地、耕地和草地3幅植被生物量空间分布图进行镶嵌,获得2000和2010年长三角地区植被生物量空间分布栅格图。
3 结果与分析
3.1 植被生物量空间分布
长三角地区植被生物量空间分布总体呈现南高北低的态势,沿长江地区和太湖流域生物量相对低。采用自然断裂法将2000年植被生物量划分为5个等级,从高到低依次为高值区(>4.5 kg·m-2)、较高值区(>3.3~4.5 kg·m-2)、中值区(>2.5~3.3 kg·m-2)、较低值区(>1.6~2.5 kg·m-2)和低值区(0.1~1.6 kg·m-2)。为便于比较,将2010年植被生物量按2000年分级标准划分为5个等级(图1)。
2000年长三角地区植被生物量面积最大分区为低值区,达61 473.4 km2,其他分区由高到低依次为中值区、较低值区、较高值区和高值区,面积分别为15 832.6、8 264.4、6 025.8和724.1 km2。植被生物量高值区和较高值区主要位于江苏的西南丘陵、浙江西部和东南部丘陵区;中值区包括浙江西部丘陵和浙东南丘陵的大部分地区;较低值区主要分布在中值区周边;低值区分布在长三角核心区内长江以北的江苏平原地区、长江以南至环杭州湾的平原地区及台州东部沿椒江地区(图1)。
2000—2010年间,植被生物量高值区面积显著增加,面积达26 179.9 km2,较高值区、中值区、较低值区和低值区面积显著减小,分别减至3 184.3、505.4、1 416.8和43 879.9 km2。2010年,因单位面积林地植被生物量的增长,浙江西部和东南部的中值区为较高值区和高值区取代;因耕地粮食单产的提高,长江以北的江苏平原地区植被生物量由低值区转变为较低值区,而长江以南至环杭州湾的平原地区及台州东部沿淑江地区仍为低值区(图1)。
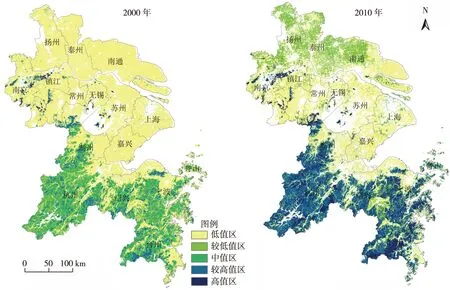
图1 2000和2010年长三角地区植被生物量密度的空间分布
3.2 10 a间植被生物量变化
10 a间,长三角地区植被生物量总量显著上升,其中,江苏部分地市和上海植被生物量稳中有升,浙江部分地市植被生物量显著增长。区域植被生物量从2000年的153.48 Tg增至2010年的202.49 Tg,增加31.93%。2000年,长三角地区江苏、浙江部分地市和上海生物量总量分别为45.93、102.74和4.81 Tg;至2010年,3者分别增至46.87、150.48和5.14 Tg,分别增长2.05%、46.47%和6.86%。
将2010和2000年植被生物量空间分布图相减,得到植被生物量变化空间分布图(图2)。采用自然断裂法分别将植被生物量增加和减少的区域划分为以下等级:高度减少区(-6.6~<-1.89 kg·m-2)、中度减少区(-1.89~<-0.65 kg·m-2)和低度减少区(-0.65~-0.026 kg·m-2),低度增长区(0.01~<1.07 kg·m-2)、中度增长区(1.07~<2.59 kg·m-2)和高度增长区(2.59~8.82 kg·m-2)。
植被生物量增长主要表现为中度增长和低度增长。中度增长区域面积为20 248.3 km2,集中分布在浙江的浙西丘陵和浙东南丘陵及宁镇扬低山丘陵。低度增长区面积达39 464.3 km2,主要包括长江以北平原、长江以南太湖以西平原、杭州湾南部和北部的沿岸平原。高度增长区面积较小,仅为6 620.9 km2,零散分布在江苏西部和浙江丘陵地区,在杭州东南部、台州南部、无锡西南部及镇江西部的丘陵地区分布相对集中。
植被生物量减少区主要分布于各地市建成区周边,以中度减少为主。中、低度减少区面积分别达18 158.3和6 460.1 km2,在沿长江、环太湖、环杭州湾及浙东南沿海等区域分布集中,呈现连片分布的态势,主要由快速城市化侵占耕地植被引起。高度减少区面积为1 583.2 km2,分布支离破碎,在江苏的环太湖、宁镇扬低山丘陵及浙江丘陵地区分散分布,主要由林地植被的损失导致。

图2 2000—2010年长三角地区植被生物量密度变化的空间分布
3.3 分地市变化特征
浙江部分地市植被生物量普遍较高,江苏部分地市和上海植被生物量偏低,不同地市生物量差距呈扩大趋势。杭州、宁波、绍兴和台州等地植被生物量远高于其他地市,2000、2010年4市植被生物量分别占总量的57.18%和64.76%。2000年生物量较小的5个地市为舟山、嘉兴、常州、无锡和镇江,5市植被生物量仅占总量的11.81%,2010年生物量较小的5个地市转变为舟山、嘉兴、常州、苏州和镇江,5市植被生物量仅占总量的9.39%(图3)。
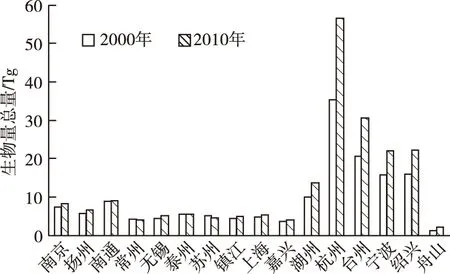
地市
除苏州、常州和泰州外,其余13个地市植被生物量均呈增长趋势。植被生物量增长最大的5个地市(杭州、台州、绍兴、宁波和湖州)占生物量增长总量的93.04%。杭州市植被生物量增长量最大,增长21.09 Tg,为2000年杭州植被生物量总量的59.54%。苏州植被生物量减少量最大,减少0.85 Tg,占2000年苏州植被生物量总量的16.13%。
16个地市植被生物量构成差异明显。计算各地市林地、耕地和草地3种植被在植被生物量中的贡献率(图4),若某市某类型植被生物量贡献率超过80%,则将该市定义为该植被类型主导型。根据2000年植被生物量构成将16个地市划分为3类:第1类是林地生物量主导型,包括杭州、台州、宁波、绍兴和舟山;第2类是耕地生物量主导型,包括上海、扬州、南通、泰州、苏州和嘉兴;第3类是林地和耕地混合主导型,包括湖州、南京、镇江、常州和无锡。
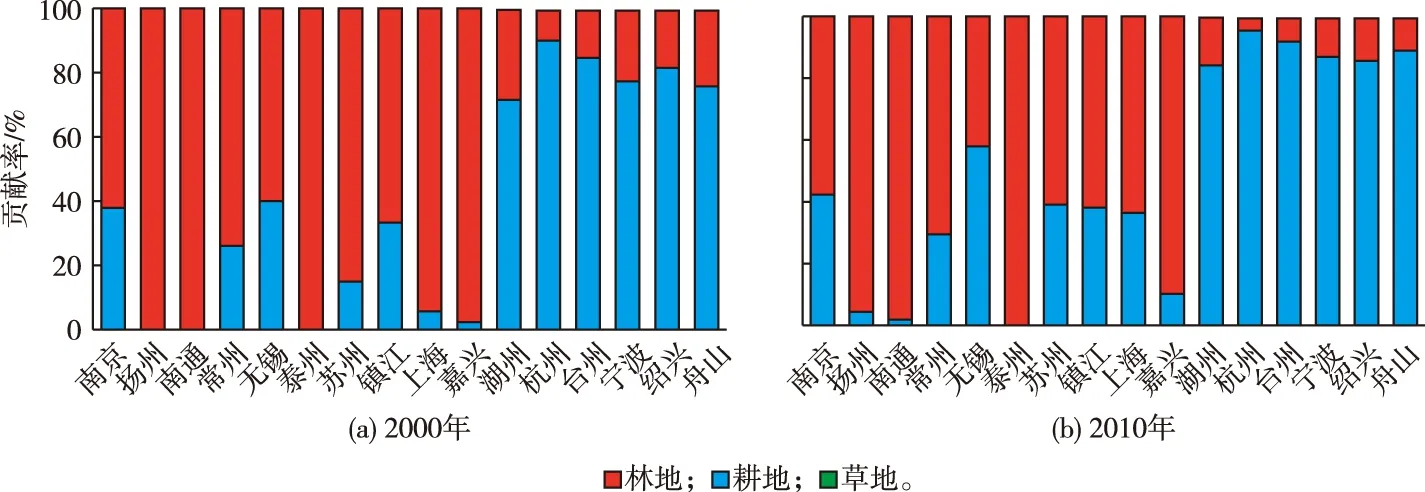
图4 2000和2010年16个地市不同类型植被生物量贡献率
分析2000—2010年3类地市植被生物量构成变化发现:林地生物量主导型的5个地市保持不变,湖州因林地生物量增长明显,由林地和耕地生物量混合主导型转变为林地生物量主导型;耕地生物量主导型地市减少,2010年仅包括扬州、南通、泰州和嘉兴;林地和耕地生物量混合主导型地市除原有南京、镇江、常州和无锡外,苏州和上海由耕地生物量主导型转变为林地和耕地生物量混合主导型。
3.4 估算结果验证
利用市(县)行政边界对长三角地区森林植被生物量空间分布图进行分区统计,与已发表文献中分市(县)的森林生物量进行对比(表1),验证森林生物量数据空间离散化精度。结果表明:多数市(县)估算精度高于80%,估算得到的森林植被生物量空间分布数据基本合理,降尺度技术对估算长三角地区植被生物量空间分布具有适用性。和估算方法有关的不同数据源和森林生物量估算方法可能导致森林生物量估算结果差异[29],笔者以土地利用/覆被数据为基础,利用森林一类清查数据估算不同森林植被类型平均生物量,以NPP为纽带进行空间估算;而已发表文献则是以森林资源一类或二类调查数据为基础的生物量估算,采用换算因子连续函数法或生物量经验(回归)模型估计法估算区域生物量密度和总量。
基于遥感模型和统计数据估算的15个地市(不含舟山)耕地植被生物量回归分析结果(图5)显示,2000和2010年两者拟合优度R2分别为0.932 7和0.913 2,认为耕地植被生物量空间估算结果保证了总体精度。
表1 笔者研究获得的森林植被生物量与相关文献中林地生物量的对比
Table 1 Forest biomass acquired by the authors and those in relevant references 万t

省份市(县)生物量总量/万t已有文献1)笔者研究2)相对误差/%江苏南京365.23284.62-22.07无锡173.97175.680.98常州131.64108.90-17.27苏州85.9378.03-9.19镇江118.99151.0626.95浙江余杭149.85112.35-25.03杭州市区303.89417.9637.54临安市1440.491416.61-1.66桐庐县643.21760.0918.17建德市931.48960.793.15淳安县2110.551762.85-16.47富阳市450.76666.6647.90
1)江苏5市生物量总量为2000年数据[30],浙江余杭为1998年数据[31],浙江除余杭以外的其他市(县)为2009年数据[32];2)江苏5市和浙江余杭生物量总量为2000年数据,浙江除余杭以外的其他市(县)为2010年数据。
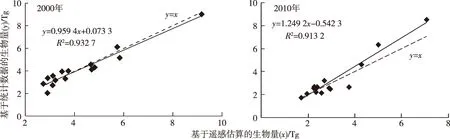
图5 基于统计数据与遥感模型的耕地植被生物量估算结果对比
3.5 不确定性分析
近10 a来长三角地区森林生态系统生物量有大幅增长,此与王磊等[30]、张茂震等[33]、聂祥永等[34]和李银等[29]对该区域的研究结果一致。对于林地植被生物量空间估算,降尺度方法建立在林地生物量与NPP呈线性相关的基础上,而实际森林生物量、NPP与林龄关系密切,受气候、人类活动等多重因素影响,这对生物量空间分布估算有一定影响。
在森林生物量的模拟和预测研究中,林龄是一个关键参数[24,35]。WANG等[36]研究表明,落叶针叶林、常绿阔叶林、热带和亚热带常绿针叶林以及落叶阔叶林单位面积NPP分别在林龄为54、40、13和122 a时达到最大值,而我国东南部分省的平均林龄普遍为20~30 a,江苏、上海和浙江中、幼龄林比例之和分别为82.63%、91.75%和76.76%[37-39]。考虑到区域尺度上NPP变化趋势与生物量相同,认为林龄对生物量空间分布格局整体影响有限[8]。
植被物候变化改变植被生长周期,对NPP和生物量产生影响[40]。研究表明随气候变暖,长三角地区森林植被生长活跃期延长,起始日期提前,终止日期延后,森林植被NDVI年均值有增加趋势[41];笔者研究得到的估算结果与之相同,即森林NPP与生物量均呈增长趋势。对于农作物,长江中下游地区降水量相对较丰富,NPP主要受制于太阳辐射[42],而长三角地区NPP与太阳辐射量的相关性高达0.739,降水与温度对NPP的直接影响较弱[43]。在土地利用/覆被变化和气候变化双重因素的驱动下,该区域土地利用/覆被变化对农田NPP总量的影响更大。
总体上,研究估算得到植被生物量空间分布,通过估算精度验证和不确定性分析,在保证植被生物量精度控制的同时,借助遥感数据在空间分布上的优势,获得了250 m×250 m的植被生物量空间分布,反映出植被生物量的空间分布及变化特征。今后可进一步结合长期固定样地的植被监测数据,研究植被生物量发展和演替规律,提高植被生物量的估算精度[44]。
4 结论
(1)集成土地利用/覆被、遥感、气象、森林资源清查和粮食产量统计等多源数据,运用遥感模型、降尺度模型和回归模型方法,可有效地对长三角地区植被生物量空间分布进行估算。估算结果既保留了统计数据的总体精度,又反映了植被生物量的空间分布特征。作为森林生物量模拟的关键参数,林龄对森林生物量估算有直接影响,但考虑到研究区森林林龄普遍较小,认为林龄对森林生物量空间分布格局整体影响有限。
(2)长三角地区植被生物量在各地市的分布具有显著的空间异质性,总体呈现南高北低的态势,沿长江地区和太湖流域生物量相对低。在区域尺度上,地域植被类型是生物量空间分布的主要影响因素,浙江的杭州、台州、宁波、绍兴和湖州森林面积和植被生物量总量均较大。10 a间植被生物量总量显著上升,从2000年的153.48 Tg增至2010年的202.49 Tg,增加31.93%。其中,江苏部分地市和上海植被生物量稳中有升,浙江部分地市植被生物量显著增长。用地类型保持不变的森林是植被生物量增长的主要来源;用地类型保持不变的耕地及绿化林地在一定程度上促进了植被生物量的增长。
(3)研究期内不同类型植被的生物量变化影响因素不同。在大尺度条件下,年均温、降水量和海拔是影响森林生物量分布的重要因素,但长三角地区气候条件较好,降水量相对较丰富,森林植被生物量增长主要受太阳辐射的影响。在气候变化背景下,植被物候变化延长了植被生长周期,有利于森林植被生物量增加。同时,研究区森林主要为中、幼龄林,随着林龄的增加,森林生物量将进一步增长。区域农作物单位面积产量呈增长态势,得益于种植水平和科技条件提高等,但增量不足以弥补耕地面积减小带来的损失。
(4)长三角地区城市化过程中的土地利用/覆被变化对植被生物量的影响显著。在沿长江、环太湖、环杭州湾及浙东南沿海等区域,建成区周边耕地向建设用地的转换呈连片分布态势,是植被生物量损失的最主要因素。在江苏的环太湖、宁镇扬低山丘陵及浙江丘陵地区,林地向建设用地的转换分散分布,导致局部植被生物量显著降低。水产养殖增加引起的耕地向水域转换在一定程度上导致植被生物量损失。尽管研究区植被生物量总量呈增加趋势,但人口和经济集聚的城市建成区及周边区域生物量的损失可能带来热岛效应、水土流失和洪涝灾害加剧等生态问题。因此,有必要通过划定生态保护红线和基本耕地红线,保护原有森林和农作物植被并进行造林绿化,使区域植被生物量总量增加且空间分布不断优化,从而缓解植被生物量损失带来的负面效应,保障区域生态安全。
[1] TAO F,YOKOZAWA M,ZHANG Z,etal.Remote Sensing of Crop Production in China by Production Efficiency Models:Models Comparisons,Estimates and Uncertainties[J].Ecological Modelling,2005,183(4):385-396.
[2] 邹长新,徐梦佳,高吉喜,等.全国重要生态功能区生态安全评价[J].生态与农村环境学报,2014,30(6):688-693.
[3] HOUGHTON R.Aboveground Forest Biomass and the Global Carbon Balance[J].Global Change Biology,2005,11(6):945-958.
[4] 朴世龙,方精云,贺金生,等.中国草地植被生物量及其空间分布格局[J].植物生态学报,2004,28(4):491-498.
[5] 陈艳梅,高吉喜,冯朝阳,等.1982—2010年呼伦贝尔植被净初级生产力时空格局[J].生态与农村环境学报,2012,28(6):647-653.
[6] 王雪,丁建伟,谭琨,等.蔚县矿区植被净初级生产力时空变化特征及影响因素[J].生态与农村环境学报,2016,32(2):187-194.[7] 方精云,郭兆迪,朴世龙,等.1981—2000年中国陆地植被碳汇的估算[J].中国科学(D辑:地球科学),2007,37(6):804-812.
[8] 刘双娜,周涛,舒阳,等.基于遥感降尺度估算中国森林生物量的空间分布[J].生态学报,2012,32(8):2320-2330.
[9] 赵明伟,岳天祥,赵娜,等.基于 HASM 的中国森林植被碳储量空间分布模拟[J].地理学报,2013,68(9):1212-1224.
[10]苏伟忠,杨桂山,甄峰.长江三角洲生态用地破碎度及其城市化关联[J].地理学报,2007,62(12):1309-1317.
[11]王雷,李丛丛,应清,等.中国1990—2010年城市扩张卫星遥感制图[J].科学通报,2012,57(16):1388-1403.
[12]杨兆平,高吉喜,沈渭寿,等.基于文献分析的中国生态服务研究[J].生态与农村环境学报,2011,27(6):1-7.
[13]GRIMM N B,FAETH S H,GOLUBIEWSKI N E,etal.Global Change and the Ecology of Cities[J].Science,2008,319(5864):756-760.
[14]GUNERALP B,SETO K C.Environmental Impacts of Urban Growth From an Integrated Dynamic Perspective:A Case Study of Shenzhen,South China[J].Global Environmental Change,2008,18(4):720-735.
[15]李双成,赵志强,王仰麟.中国城市化过程及其资源与生态环境效应机制[J].地理科学进展,2009,28(1):63-70.
[16]MORAWITZ D F,BLEWETT T M,COHEN A,etal.Using NDVI to Assess Vegetative Land Cover Change in Central Puget Sound[J].Environmental Monitoring and Assessment,2006,114(1/2/3):85-106.
[17]张祎,李玉凤,高鸿,等.基于土地转移矩阵的生态服务功能研究:以南京市仙林新市区为例[J].生态与农村环境学报,2014,30(6):800-805.
[18]李广宇,陈爽,余成,等.苏南快速城市化地区森林生物量时空变化及影响分析[J].生态环境学报,2014,23(7):1102-1107.
[19]燕守广,林乃峰,沈渭寿.江苏省生态红线区域划分与保护[J].生态与农村环境学报,2014,30(3):294-299.
[20]李广宇,陈爽,余成,等.长三角地区植被退化的空间格局及影响因素分析[J].长江流域资源与环境,2015,24(4):572-577.
[21]侯学煜.中国植被图集(1∶100万)[M].北京:科学出版社,2001.(2013-07-11)[2014-10-20].http:∥westdc.westgis.ac.cn/data/2368aa82-c1be-4f0f-b4ef-391a6f0c4e8c.
[22]李广宇.都市密集区植被生物量变化及其与城市增长的空间联系研究[D].北京:中国科学院大学,2015.
[23]朱文泉,潘耀忠,张锦水.中国陆地植被净初级生产力遥感估算[J].植物生态学报,2007,31(3):413-424.[24]戴铭,周涛,杨玲玲,等.基于森林详查与遥感数据降尺度技术估算中国林龄的空间分布[J].地理研究,2011,30(1):172-184.
[25]KINDERMANN G E,MCALLUM I,FRITZ S,etal.A Global Forest Growing Stock,Biomass and Carbon Map Based on FAO Statistics[J].Silva Fennica,2008,42(3):387-396.
[26]方精云,刘国华,徐嵩龄.我国森林植被的生物量和净生产量[J].生态学报,1996,16(5):497-508.
[27]胡会峰,王志恒,刘国华,等.中国主要灌丛植被碳储量[J].植物生态学报,2006,30(4):539-544.
[28]杜鑫,蒙继华,吴炳方.作物生物量遥感估算研究进展[J].光谱学与光谱分析,2010,30(11):3098-3102.
[29]李银,陈国科,林敦梅,等.浙江省森林生态系统碳储量及其分布特征[J].植物生态学报,2016,40(4):354-363.
[30]王磊,丁晶晶,季永华,等.江苏省森林碳储量动态变化及其经济价值评价[J].南京林业大学学报(自然科学版),2010,34(2):1-5.
[31]李惠敏,陆帆,唐仕敏,等.城市化过程中余杭市森林碳汇动态[J].复旦学报(自然科学版),2004,43(6):1044-1050.
[32]项茂林,陶吉兴,季碧勇,等.基于市县联动监测的县域森林植被生物量评估[J].浙江林业科技,2012,32(6):22-26.
[33]张茂震,王广兴.浙江省森林生物量动态[J].生态学报,2008,28(11):5665-5674.
[34]聂祥永,姚顺彬,楼毅,等.长江三角洲地区森林碳汇的初步估算与分析[J].林业资源管理,2012,41(6):51-56.
[35]BRADFORD J B,BIRDSEY R A,JOYCE L A,etal.Tree Age,Disturbance History,and Carbon Stocks and Fluxes in Subalpine Rocky Mountain Forests[J].Global Change Biology,2008,14(12):2882-2897.
[36]WANG S,ZHOU L,CHEN J,etal.Relationships Between Net Primary Productivity and Stand Age for Several Forest Types and Their Influence on China′s Carbon Balance[J].Journal of Environmental Management,2011,92(6):1651-1662.
[37]徐冰,郭兆迪,朴世龙,等.2000—2050 年中国森林生物量碳库:基于生物量密度与林龄关系的预测[J].中国科学:生命科学,2010,40(7):587-594.
[38]温小荣,蒋丽秀,刘磊,等.江苏省森林生物量与生产力估算及空间分布格局分析[J].西北林学院学报,2014,29(1):36-40.
[39]钱逸凡,伊力塔,张超,等.浙江省中部地区公益林生物量与碳储量[J].林业科学,2013,49(5):17-23.
[40]韦振锋,王德光,张罛,等.近12年陕甘宁黄土高原区植被物候时空变化特征[J].生态与农村环境学报,2014,30(4):423-429.
[41]金佳鑫,江洪,张秀英,等.利用遥感监测长江三角洲森林植被物候对气候变化的响应[J].遥感信息,2011,26(2):79-85.
[42]王轶虹,王美艳,史学正,等.2010年中国农作物净初级生产力及其空间分布格局[J].生态学报,2016,36(19):1-11.
[43]戴靓,周生路,吴绍华,等.近十年长三角地区陆地净第一性生产力时空变化[J].长江流域资源与环境,2012,21(10):1216-1222.
[44]商侃侃,郑思俊,张庆费.上海海湾国家森林公园1 hm2样地群落结构特征及其动态监测意义[J].生态与农村环境学报,2013,30(3):316-321.
(责任编辑: 李祥敏)
Variation of Spatial Estimation and Distribution of Vegetation Biomass in Yangtze River Delta During 2000-2010.
LI Guang-yu1,2, CHEN Shuang1, ZHANG Hui2, ZHANG Tong1
(1.Nanjing Institute of Geography & Limnology, Chinese Academy of Sciences, Nanjing 210008, China;2.Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection, Nanjing 210042, China)
Vegetation biomass is an important indicator of the service function of terrestrial ecosystems. It is of great significance to the formulation of rational vegetation protection policies to ascertain spatial distribution rules and temporal variation of vegetation biomass. Though a lot of works have been done on estimating vegetation biomass at regional and global scales, little is available in the literature about spatial estimation of biomass of an ecosystem as a whole and, what is more, spatial resolution of the estimation is too low to reflect changes in land use/cover. Based on multi-sourced data including remote sensing, meteorology, land use/cover, forest inventory, and grain yield, spatial distribution of vegetation biomass at 250 m resolution was studied with the aid of remote-sensing models, spatial downscaling technique, GIS spatial operation and mathematical statistical analysis. Results show that the spatial estimation of vegetation biomass not only inherits the accuracy of statistical data, but also reflects spatial distribution of vegetation biomass. The vegetation biomass in the Yangtze River Delta (YRD) displays a general distribution trend of rising from north to south and being relatively low alongside the Yangtze River and around the Tai Lake Basin. The total vegetation biomass increased significantly during the years from 2000 to 2010. It increased significantly or by 47.74 Tg in parts of Zhejiang, and remained stable with a slight rise (0.94 Tg) in parts of Jiangsu and (0.33 Tg) in Shanghai. Vegetation biomass is closely related to land use/cover (LUCC). The growth of biomass is mainly attributed to the maintenance of forest lands and farmlands in land use. In highly developed plain areas, conversion of forest lands and farmlands has led to significant decline in biomass, however, greening of construction lands helps offset the declining trend of biomass triggered by destruction of vegetation.
vegetation biomass;spatial estimation;spatial distribution;Yangtze River Delta
2016-02-29
国家自然科学基金(41371179);中央级公益性科研院所基本科研业务专项(20160304)
X24;Q948.1
A
1673-4831(2016)05-0708-08
10.11934/j.issn.1673-4831.2016.05.004
李广宇(1987—),男,河南濮阳人,助理研究员,博士,主要研究方向为城市生态学。E-mail: lgy@nies.org
① 通信作者E-mail: schens@niglas.ac.cn
