猪胰岛素诱导1基因的cDNA克隆和差异表达及蛋白质序列分析
2016-10-27王京京魏麟陈斌
王京京,魏麟,陈斌*
猪胰岛素诱导1基因的cDNA克隆和差异表达及蛋白质序列分析
王京京1,魏麟2,陈斌1*
(1.湖南农业大学动物科学技术学院,湖南长沙 410128;2.怀化学院生物与食品工程学院,湖南怀化 418000)
克隆猪胰岛素诱导1()基因,并对其蛋白质序列进行分析。根据人基因的保守序列设计1对引物,以猪总RNA为模板,经RT–PCR获得基因序列,通过相对荧光定量PCR分析该基因在不同组织中的表达量;将基因序列连接到pMD19–T Simple载体上,用PCR检测阳性克隆后测序,并对克隆得到的基因进行生物信息学分析。结果表明,基因在肺中的表达量最高,其次是在肝脏,再次是在脾脏,在心脏中的表达量最低;通过克隆,获得了一段999 bp的序列,其中831 bp的开放阅读框编码276个氨基酸;该蛋白不含信号肽序列,与其他动物基因氨基酸序列的一致性在71%以上。
猪;胰岛素诱导1基因;基因克隆;序列分析
胰岛素诱导基因(insulin induced gene,)是近年来被发现在脂肪代谢调控方面起重要作用的基因[1],是其中的一个亚型[2]。Mohn等[3]于1991年在小鼠肝脏以及鼠H35肝癌细胞中分离到的mRNA; Peng等[4]最早从再生肝脏中克隆得到了基因;基因能使固醇调节元件结合蛋白(sterol regulatory element–binding protein, SREBPS)家族成员激活,通过与3–羟基–3–甲基戊二酰辅酶A 还原酶(3–hydroxy–3–methylglutary– CoA reductase, HMGR)的固醇敏感域(sterol sensing domain, SSD) 和SRE BPS裂解活化蛋白(SREBPS cleavage activating protein, SCAP)结合,调节胆固醇的合成,调控脂肪代谢。是反馈调节胆固醇和脂肪合成的重要因子,能有效调控脂肪细胞的分化、成熟与脂质合成[6]。基因表达产物在脂质合成的反馈调节中起重要作用。基因受多种物质的调节,在不同物种、不同组织中的表达量也不同。近年来,人类和小鼠基因的克隆、分离、表达和调控成为热点。邱欢[7]对基因进行了染色体定位和生物学特征鉴定;朱亚南等[8]研究了基因在猪前体脂肪细胞分化过程的表达。除上述报道外,其他关于猪基因的报道较少。本试验中采用RT–PCR方法克隆基因的cDNA片段,检测该基因在猪不同组织器官中的特异性表达,对其进行测序并进行生物信息学分析。现将结果报道如下。
1 材料与方法
1.1材料与试剂
采集黔邵花猪的心脏、肝脏、脾脏、肺、胃、肾脏和肌肉共7种样品,清洗干净后用乙醇擦拭,经焦酸二乙酯(DEPC)水处理后,–80 ℃冷冻保存,备用。
RNA提取试剂盒、cDNA合成试剂盒、质粒提取试剂盒、DNA回收纯化试剂盒、荧光定量试剂盒SYBR Prime ScriptTMRT–PCR Kit、酶、pMD19–T Vector载体、菌种JM109、DNA相对分子质量标记、T4DNA连接酶及电泳类试剂等,均购自TaKaRa公司。
1.2引物设计
用Oligo 6软件设计所有引物序列。根据NCBI/GenBank中登录的人基因序列(NM_005542),设计1对猪的特异性克隆引物。上游引物F,5–GCCGGATCCATGCCCAGACT GGACGACCA–3;下游引物R,5–ACCGAATTCTC AGTCGCTGTGTGGCTT–3。采用实时荧光定量PCR检测猪基因表达水平的引物(上游引物,AGCGGCCACACCAACAGCTGG;下游引物,TGGCTGTCGATGCAGGGGTA)。以甘油醛–3–磷酸脱氢酶()基因为参照,根据NCBI/ GenBank中登录的猪基因(NM_ 001206359)序列设计其特异性引物(上游引物,CACTGACACG TTGGCAGTGG;下游引物,CATGGAGAAGGCTG GGGCTC)。以上引物均由生工生物工程(上海)股份有限公司合成。
1.3方法
1.3.1猪基因的克隆与测序
按照Trizol试剂说明书提取猪心脏、肝脏、脾脏、肺、胃、肾脏和肌肉的RNA,检测RNA的纯度和浓度。根据cDNA合成试剂盒说明书进行cDNA链合成。将反转录产物分离纯化。PCR反应条件:94 ℃预变性3 min;94 ℃变性50 s,55 ℃退火50 s,72 ℃延伸50 min,35个循环;72 ℃延伸10 min。将回收的PCR产物与pMD19–T Vector载体连接,转到JM109感受态细胞中。挑取阳性菌落培养过夜,将获得的重组子用I和I双酶切鉴定。提取质粒送至生工生物工程(上海)股份有限公司测序。
1.3.2相对荧光定量PCR表达分析
采用相对荧光定量PCR方法,按照荧光定量试剂盒SYBR Prime ScriptTMRT–PCR Kit的操作说明,以基因的表达为对照,以SYBR为荧光染料,检测基因在猪心脏、肝脏、脾脏、肺、胃、肾脏和肌肉中的相对表达量。用ABI 7500实时定量PCR仪进行检测。反应体系:总RNA 2 μL,随机引物(100 μmol/L)1 μL,寡聚胸苷酸引物(50 μmol/L)1 μL,反转录酶1 μL,5×缓冲液4 μL,补ddH2O至20 μL。荧光定量PCR扩增反应体系:cDNA模板2 μL,上下游引物(10 μmol/L)均为0.375 μL,2×SYBR Premix ExTM 10 μL,50×ROX Reference DyeⅡ0.4 μL,补ddH2O至20 μL。PCR反应程序:95 ℃预变性3 min;95 ℃变性20 s,60 ℃复性30 s,35个循环。每个试验重复3次。采用Excel和ABI 7500 PCR仪的Sequence Detection software 软件[9]对数据进行分析。
1.3.3生物信息学分析
采用Expasy在线软件(http://expasy.org/tools)与DNA Star进行序列分析。编码蛋白氨基酸序列的基本理化性质用ProtParam程序[10]进行分析;信号肽、跨膜结构域、二级结构和三级结构分别用SignalP 4.1 Server[11]软件、TMHMM Server V. 2.0、SOPMA程序和SWISS–MODEL[12–14]进行预测与分析;用NCBI中的Blastp在线工具进行其他物种的同源氨基酸序列搜索,用MEGA 6.0软件进行聚类分析。
2 结果与分析
2.1基因的克隆与测序
提取的猪肌肉总RNA经琼脂糖凝胶电泳检测,28S rRNA 和18S rRNA 条带清晰(图1),提取的RNA完整性较好;经纯度检测,260 nm与280 nm比值的平均值为1.91,说明纯度较高,可用于后续试验。

泳道1和2为样品总RNA。
以猪基因组cDNA为模板进行PCR扩增,得到的特异性条带约为999 bp(图2)。经克隆测序,该片段大小与预期目标片段大小相符。
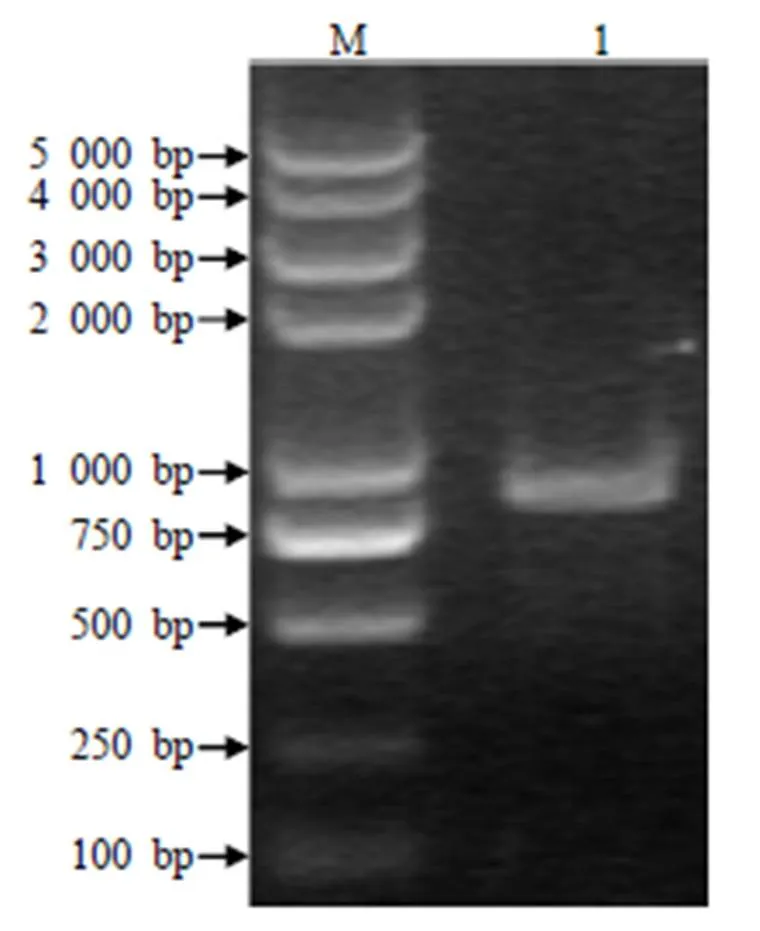
M DNA Marker 5 000;1 INSIG基因扩增产物。
用I和I内切酶双酶切鉴定重组质粒,得到2条片段,其中一条约为3 000 bp的pMD19–T线性片段,另一条约为1 000 bp的插入片段(图3),片段大小与预期的大小相符。
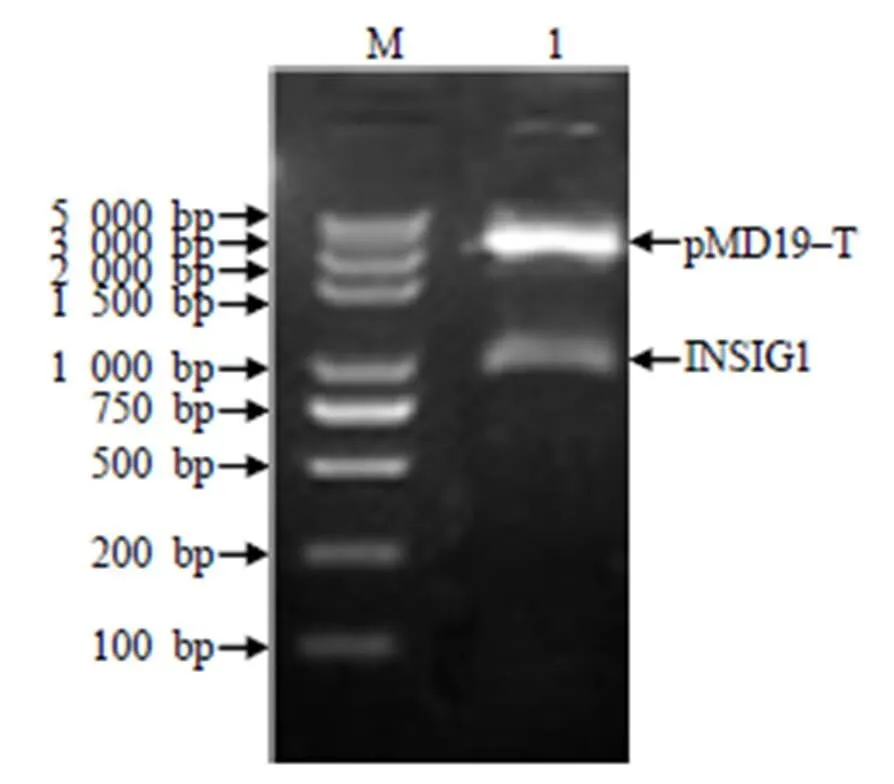
M DNA Marker 5 000;1 pMD19–T/INSIG1。
对鉴定正确的重组质粒进行测序,得到的序列长度为999 bp。该序列含有1个完整的开放阅读框架(ORF),共编码276个氨基酸(图4)。经BLAST比对,克隆得到的DNA序列为猪的序列。
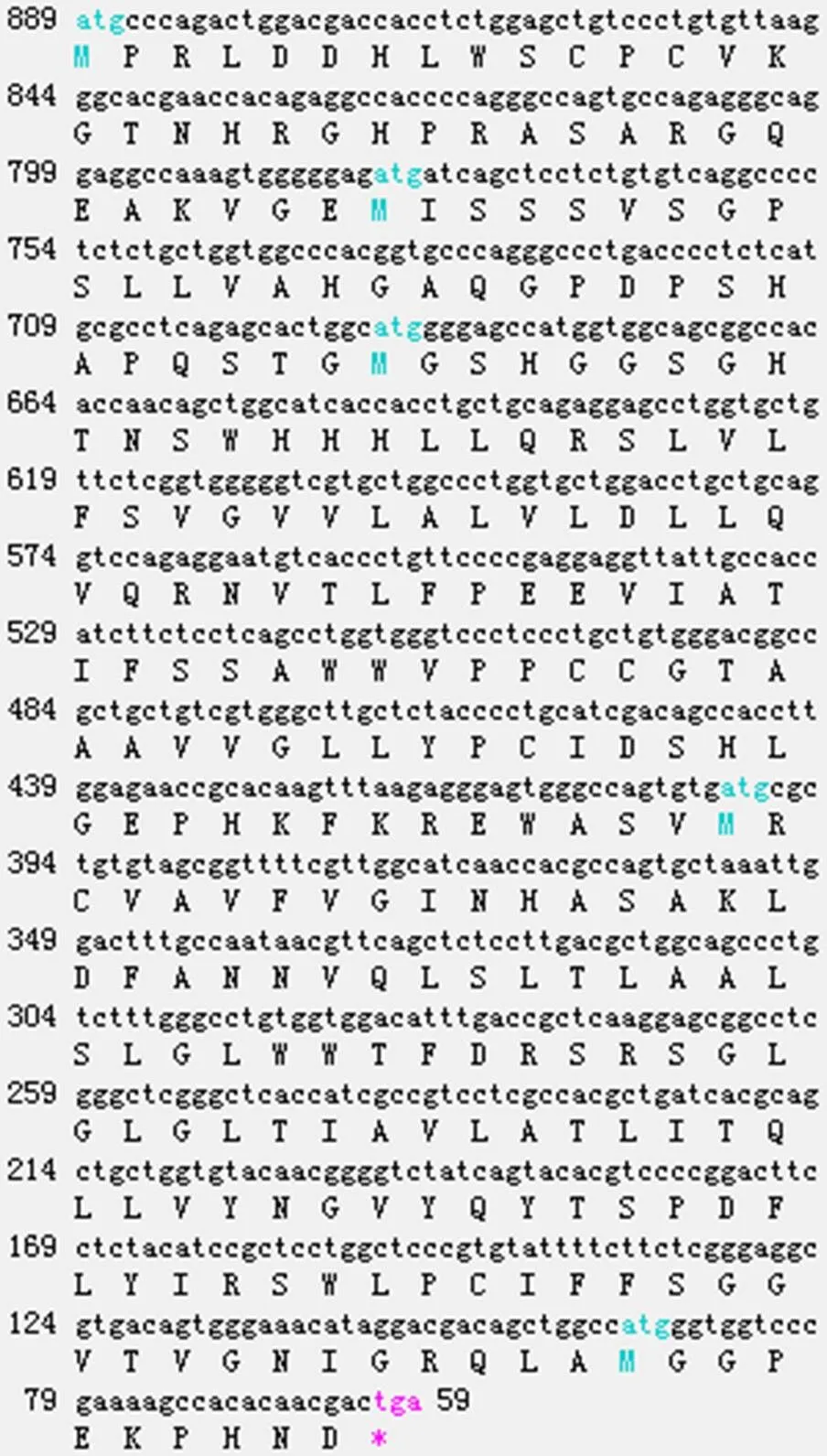
图4 猪INSIG1基因的碱基和氨基酸序列
2.2猪的差异表达
分别以基因为参照,采用荧光定量PCR检测基因在猪不同组织中的表达量,检测结果表明,该基因在肺中的表达量最高,其次为肝脏,再次为脾脏,在胃和肾脏中的表达较低,在心脏和肌肉中的表达最低(图5)。虽然在各组织中均检测到,但该基因在各组织中的表达量差异显著,表明其表达具有组织特异性。

图5 INSIG1基因在不同组织中的相对表达量
2.3猪基因的生物信息学分析
猪理化性质分析结果表明:基因编码的蛋白含有276个氨基酸残基,该蛋白的相对分子质量为29 735.2,理论等电点为8.21,其氨基酸组成中亮氨酸含量达12.3%,分子式为C1339H2075N373O372S12,半衰期为30 h,不稳定系数为32.08,属于稳定蛋白,脂肪系数为96.41,总平均亲水性为0.176。分析猪所编码的氨基酸序列,发现该序列不含信号肽,属于非分泌蛋白。对猪INSIG1进行跨膜区分析的结果表明,该蛋白有5个跨膜区,分布在82~104、124~146、186~203、208~230和240~262位氨基酸之间(图6)。二级结构预测结果显示,该蛋白由α–螺旋、β–转角、伸展链和无规卷曲构成,占总蛋白的比例分别为37.68%、6.52%、15.22%和40.58%(图7),其三级结构预测结果见图8。
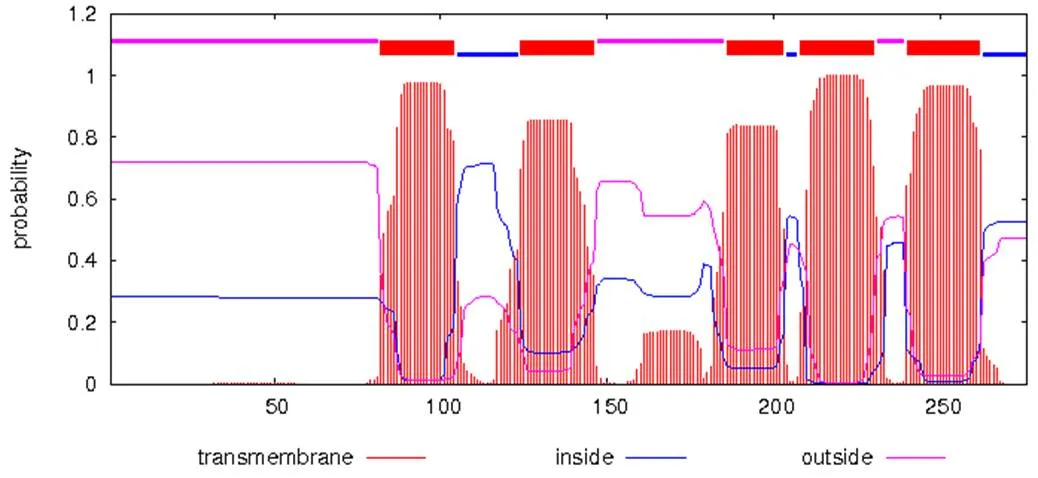
图6 TMHMM程序对猪INSIG1跨膜结构的预测结果

图7 SOPMA对猪INSIG1二级结构的预测结果
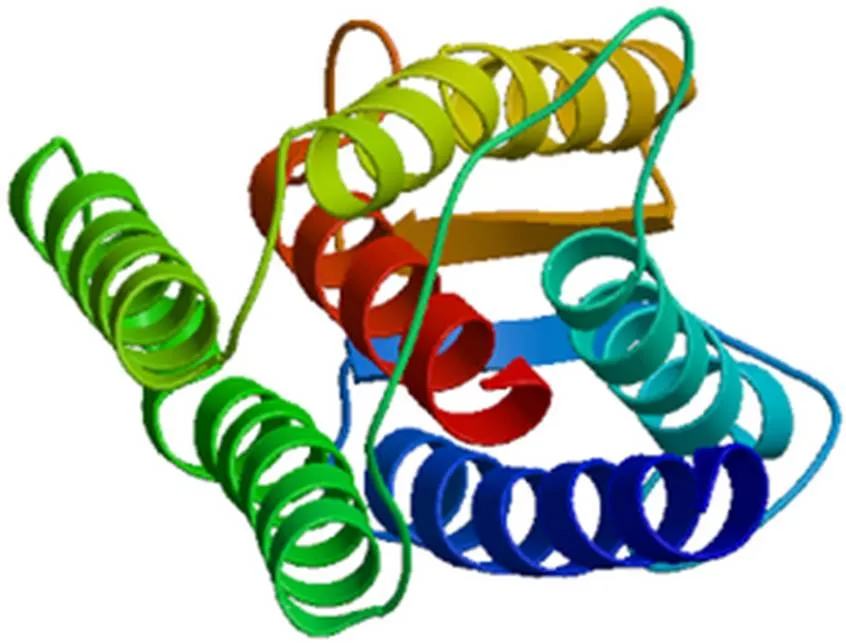
图8 猪INSIG1蛋白三维结构预测结果
2.4猪INSIG1氨基酸序列的一致性和亲缘关系分析
氨基酸序列对比结果表明,猪INSIG1与人(NP_005533)、小鼠(NP_705746)、中国仓鼠(NP_001231008)、布氏鼠耳蝠(EPQ19350)、牦牛(ELR49247)、水牛(NP_001277853)、牛(NP_001071377)、山羊(NP_001273017)、双峰驼(EPY79870)、食蟹猴(NP_001274621)、眼镜王蛇(ETE72252)、绿头鸭(EOA96705)和野鸡(NP_001026137)的INSIG1氨基酸序列一致性分别为84%、75%、75%、98%、90%、89%、89%、89%、89%、84%、76%、75%和71%,序列的一致性较高,表明基因在进化过程中具有一定的保守性。图9分析结果显示:猪与布氏鼠耳蝠、双峰驼的亲缘关系较近,与水牛、山羊、牛、牦牛的亲缘关系次之,而与眼镜王蛇、绿头鸭、野鸡等非哺乳动物的亲缘关系相对较远。
3 讨论
有研究表明,西农萨能奶山羊基因含有1个831 bp的完整阅读框,编码276个氨基酸[15]。本研究获得的猪基因编码的氨基酸数与西农萨能奶山羊的一致,比人的INSIG1蛋白氨基酸组成数少1个,与其他动物INSIG1蛋白氨基酸序列的一致性在71%以上。
据报道,基因在秦川牛组织中差异表达,且在肝脏中的表达量最高[16];在西农萨能奶山羊10个组织中均有表达,其中在肝脏中的相对表达量最高,在心脏中的相对表达量最低[15];在鼠肝脏、肾脏的表达量较高,在其他组织的表达量较低[17]。本研究中,基因在猪的7个组织中均有表达,在肺中的表达量最高,其次为肝脏,在心脏中的表达量最低。基因在各组织间的表达差异可能与组织功能差异有关,还可能受生长发育等其他因素影响,如肝脏中基因mRNA的表达量随肝脏的发育而变化[18]。
INSIG1是一种跨膜蛋白,TMHMM预测到猪INSIG1蛋白和西农萨能奶山羊INSIG1蛋白一样,只有5个跨膜区,比人INSIG1蛋白少1个。猪与人INSIG1氨基酸序列的一致性为84%,与哺乳动物的亲缘关系相对较近,猪INSIG1序列的一致性较高,表明基因在进化过程中具有一定的保守性。
参考文献:
[1] 莫朝晖,廖二元.胰岛素诱导因子与脂代谢[J].国际内分泌代谢杂志,2008,28(5):320–322.DOI: 10.3760/cma.j.issn.1673–4157.2008.05.010.
[2] Yabe D,Komuro R,Liang G,et al.Liver-specific mRNA for Insig–2 down-regulated by insulin:implications for fatty acid synthesis[J].Proc Natl Acad Sci USA,2003,100(6):3155–3160.DOI:10.1073/pnas.0130116100.
[3] Mohn K L,Laz T M,Hsu J C,et al.The immediate-early growth response in regenerating liver and insulin- stimulated H–35 cells:comparison with serum-stimulated 3T3 cells and identification of 41 novel immediate-early genes[J].Mol Cell Biol,1991,11(1):381–390.DOI: 10.1128/mcb.11.1.381.
[4] Peng Y,Schwarz E J,Lazar M A,et al.Cloning,human chromosomal assignment,and adipose and hepatic expression of the CL–6/INSIG1 gene[J].Genomics,1997,43(3):278–284.DOI:10.1006/geno.1997.4821.
[5] Yang T,Espenshade P J,Wright M E,et al.Crucial step in cholesterol homeostasis:sterols promote binding of SCAP to INSIG–1,a membrane protein that facilitates retention of SREBPs in ER[J].Cell,2002,110(4):489–500.DOI:10.3410/f.1003998.146257.
[6] Sever N,Yang T,Brown MS,et al.Accelerated degradation of HMG CoA reductase mediated by binding of insig–1 to its sterol-sensing domain[J].Mol Cell,2003,11(1):25–33.DOI:10.1016/s1097–2765(02)00822–5.
[7] 邱欢.猪SCAP和INSIG家族基因的克隆、染色体定位及生物学特征研究[D].武汉:华中农业大学,2005.10.7666/d.Y806701.
[8] 朱亚南,李青莹,任书强,等.胰岛素诱导基因1和2在猪前体脂肪细胞分化过程的表达[J].吉林农业大学学报,2014,36(2):194–198.DOI:10.13327/j.jjlau.2014. 1911.
[9] Livak K J,Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(–Delta Delta C(T)) method[J].Methods,2001,25(4):402–408.DOI:10.1006/meth.2001.1262.
[10] Gasteiger E,Hoogland C,Gattiker A,et al.Protein identification and analysis tools on the ExPASy server[J]. The Proteomics Protocols Handbook,2005:571– 607.DOI:10.1385/1–59259–890–0:571.
[11] Petersen T N,Brunak S,von Heijne G,et al.SignalP 4.0:discriminating signal peptides from transmembrane regions[J].Nat Methods,2011,8(10):785–786.DOI:10. 1038/nmeth.1701.
[12] Arnold K,Bordoli L,Kopp J,et al.The SWISS–MODEL workspace:a web-based environment for protein structure homology modelling[J].Bioinformatics,2005,22(2):195–201.DOI:10.1093/bioinformatics/bti770.
[13] Kiefer F,Arnold K,Künzli M,et al.The SWISS- MODEL Repository and associated resources[J]. Nucleic Acids Res,2009,37(Database issue):D387–D392.DOI: 10.1093/nar/gkn750.
[14] Benkert P,Biasini M,Schwede T.Toward the estimation of the absolute quality of individual protein structure models[J].Bioinformatics,2011,27(3):343–350.DOI: 10.1093/bioinformatics/btq662.
[15] 王慧,罗军,胡仕良,等.西农萨能奶山羊INSIG–1编码区的克隆、序列特征分析和组织表达[J].西北农林科技大学学报(自然科学版),2013,41(8):24–30.
[16] Liu Y,Liu X L,He H,et al.Four SNPs of insulin-induced gene 1 associated with growth and carcass traits in Qinchuan cattle in China[J].Genet Mol Res,2012,11(2):1209–1216.DOI:10.4238/2012.May.8.3.
[17] Taub R,Mohn K L,Diamond R H,et al.Molecular aspects of insulin-regulated hepatic growth [J].Molecular Biology of Diabetes,1994,46:301–320.
[18] Haber B,Naji L,Cressman D,et al.Coexpression of liver-specific and growth-induced genes in perinatal and regenerating liver:attainment and maintenance of the differentiated state during rapid proliferation[J]. Hepatology,1995,22(3):906–914.DOI:10.1002/ hep.1840220331.
Cloning and differential expression of insulin induced gene 1 in pig and the analysis on its protein sequence
Wang Jingjing1, Wei Lin2, Chen Bin1*
(1. College of Animal Science and Technology, Hunan Agricultural University, Changsha 410128, China 2. College of Biological and Food Engineering, Huaihua University, Huaihua, Hunan 418000, China)
The study aims to clone the pig insulin induced gene 1() and conducts its protein sequences analysis. According to conserved sequence genein human, a pair of primers was designed. By adopting pig total RNA as template, thegene sequences was cloned using RT–PCR, then thegene was connected to pMD 19–T Simple vector. The positive clone was sequenced after the identification of clone by PCR, and the sequence characterization was performed from biology information analysis software. The results from relative real-time PCR analysis indicated thattranscript abundance was the highest in lung, next in liver, spleen, and heart in turn. The experiment obtained a 999 bp sequence, of which included 831 bp open reading frame encoded with 276 amino acids. From the results of bioinformatics analysis showed that the protein does not contain signal peptide sequence, and from the results of amino acid sequence alignment showed that pighad high similarity score (more than 71%) with other animals in GenBank.
pig; insulin induced gene 1(); gene clone; sequence analysis
S828.2
A
1007-1032(2016)05-0505-06
2015–12–10
2016–04–06
国家现代农业产业技术体系建设专项(CARS–36);湖南农业大学青年科学基金项目(14QN27)
王京京(1981—),女,湖南常德人,博士研究生,主要从事动物遗传育种研究,304774418@qq.com;*通信作者,陈斌,博士,教授,主要从事动物遗传育种与繁殖研究,chenbin7586@126.com
投稿网址:http://xb.ijournal.cn
责任编辑:王赛群
英文编辑:王库
