重金属铅胁迫对刺槐幼苗生长及铅离子转运特性的影响
2016-10-26江泽平
周 建,江泽平,魏 远
(1.中国林业科学研究院 林业研究所,北京 100091;2.河南科技学院 园艺园林学院,河南 新乡453003)
重金属铅胁迫对刺槐幼苗生长及铅离子转运特性的影响
周建1,2,江泽平1,魏远1
(1.中国林业科学研究院 林业研究所,北京 100091;2.河南科技学院 园艺园林学院,河南 新乡453003)
为了解刺槐Robinia pseudoacacia对铅胁迫的耐受程度与耐受机制,以刺槐幼苗为研究对象,用硝酸铅溶液浇灌处理,研究幼苗的生长特性、组织铅离子质量分数(μg·g-1)及离子转运特征。结果表明:250 mg·L-1铅处理对刺槐幼苗的茎高、地径、干质量、鲜质量等有一定的促进作用;在中高铅离子质量浓度(≥500 mg·L-1)下,刺槐幼苗的生长受到抑制。随着铅处理质量浓度的增加,幼苗根系铅离子质量分数显著上升(P<0.05);茎部铅质量分数先升后降;叶中铅质量分数大体呈现降低趋势;叶/茎离子转运率急剧下降(P<0.05);(叶+茎)/根、茎/根离子转运率先升后降。通过多元回归分析,发现根系铅质量分数、(叶+茎)/根离子转运对幼苗干物质积累影响最大;茎部铅离子质量分数、根茎间铅离子转运对幼苗地径、苗高生长影响最大。图3表2参23
植物学;铅胁迫;刺槐;幼苗;生长;离子转运
土壤重金属污染已成为当前亟须解决的重大环境问题[1-3]。中国大部分区域土壤目前正在面临严重的污染问题,近1/5的耕地受到影响,其中铅锌铜镉等4种重金属污染占全部污染土地面积的70%以上。土壤重金属污染不仅导致土地生产力下降,也使得重金属元素进入食物链,严重危害人类健康。铅为最重要的土壤重金属污染元素之一[4]。针对土壤铅污染,植物修复是一种非常重要的技术手段,而修复植物选择则是其中的关键。近年来,有关铅污染条件下植物修复的研究报道颇多[5-12],但主要集中于草本,如蜈蚣草Nephrolepis auriculata,堇菜Viola verecunda,狗牙根Cynodon dactylon,早熟禾Poa annua等[13-16]。草本的生物量较小,根系分布较浅,对重金属吸收效果存在一定的局限性。因此,筛选对重金属污染耐性强、富集能力与固定能力强、生物量大的树种,实现对重金属污染物的快速吸收与转移成了植物修复重金属污染的新方向与新亮点。本研究以刺槐Robinia pseudoacacia的幼苗生长及铅离子转运特性为对象,研究刺槐对铅胁迫的耐性程度,通过离子吸收转运,了解刺槐对铅胁迫的耐受机制,为铅污染耐受性树种选择奠定理论基础。
1 材料和方法
1.1试验材料与处理方法
用40℃温水浸泡刺槐种子24 h,6 h更换1次温水。将种子埋于含水量60%的湿砂中放在向阳处催芽。待刺槐种子露白时,选取均匀饱满的刺槐种子360粒,播于事先准备好的栽培钵中。每只杯子中放入未处理的土壤,播种5颗露白刺槐种子,在种子上盖上1.00~1.50 cm土壤,浇透水。
待出苗4周后,配制质量浓度为0,250,500,1 000和1 500 mg·L-1的硝酸铅溶液,分别浇灌栽培钵中的刺槐幼苗,直至钵底托盘出现处理液,保证土壤浇透。连续浇灌3次,1周1次,以蒸馏水作对照。栽培钵4盆·处理-1,3次重复,共12盆。
1.2生长指标测定
待铅胁迫处理6周后,各个处理水平中选出具有代表性的幼苗,用游标卡尺测出幼苗的茎高和地径。用1/1 000电子天平测量幼苗鲜质量。将幼苗放至烘箱内,105℃杀青5 min,70℃恒温处理12 h,烘至恒量后称干质量。重复3次。
1.3铅质量分数测定
取烘干幼苗的叶,碾碎、研磨后倒入烧杯,加入7 mL硝酸和2 mL过氧化氢混合,放入微波中消解(150℃,20 min,5 W)后取出,加入体积分数为0.20%的稀硝酸转移至聚四氯乙酸烧杯中,放置电热板上170℃赶酸至近干,用体积分数为0.20%的稀硝酸转移至25 mL容量瓶中定容。按上述步骤对茎和根进行硝化处理。最后用Optima 2100 DV电感耦合等离子体发射光谱仪(美国PE公司,PerkinElmer)进行铅离子的全量分析。
干物质铅离子质量分数(μg·g-1)=测定液铅质量浓度(mg·L-1)×25 mL/各部分物质的干质量(g)。
铅离子转运率=(上部位置铅质量分数×相应干物质质量)/(下部位置铅质量分数×相应干物质质量)。
1.4数据处理
利用SPSS 21.0进行方差分析、多元线性回归和相关分析。
2 结果与分析
2.1不同铅胁迫质量浓度下刺槐幼苗的生长性状
由图1A可知:在250 mg·L-1铅处理下,刺槐幼苗的地径比对照植株地径增加了2.7%,但差异不显著(P>0.05)。随着铅质量浓度的增加,刺槐幼苗的地径逐步降低,在1 500 mg·L-1处理下,其幼苗地径最小。
由图1B可知:250 mg·L-1铅处理的刺槐幼苗的茎高比对照植株增加了2.2%,但差异不显著(P>0.05)。其他处理下刺槐幼苗的茎高均低于对照,但差异不显著(P>0.05),其中在500 mg·L-1处理的幼苗茎高最小,分别比对照与250 mg·L-1处理低14.0%和15.8%。

图1 铅胁迫对刺槐幼苗生长性状地径(A),苗高(B),鲜质量(C)和干质量(D)的影响Figure 1 Effect of Pb stress on growth characteristics including diameter(A),seedling height(B),fresh weight(C),dry weight(D)of Robinia pseudoacacia seedlings
250 mg·L-1铅处理的刺槐幼苗鲜质量值最高,比对照植株增加了2.6%,但差异不显著(P>0.05,图1C)。随着处理质量浓度的增加,刺槐幼苗鲜质量大体呈降低趋势,但各处理间差异不显著(P>0.05),其中在500 mg·L-1处理的幼苗鲜质量值最小,分别比对照与250 mg·L-1处理低17.6%和19.7%。
随着铅处理质量浓度的增加,幼苗干物质质量呈现递减趋势,各处理之间差异不显著(P>0.05),其中1 000 mg·L-1处理的干物质质量最小,低于对照值23.60%(图1D)。
2.2不同铅胁迫质量浓度下幼苗各部位铅离子质量分数
由图2可知:随铅质量浓度的上升,铅在根中的质量分数逐渐增加,在中低质量浓度(≤500 mg·L-1)时,根部铅离子质量分数增加幅度较小,处理间差异不显著(P>0.05);当铅质量浓度大于500 mg·L-1时,铅在根部的质量分数显著增加(P<0.05),尤其在1 000和1 500 mg·L-1时,幼苗根系中铅质量分数分别达到70.81和93.28 μg·g-1。随着铅处理质量浓度上升,刺槐幼苗茎部铅质量分数呈现单峰曲线,先上升后下降;各相邻处理间差异较大,变化显著(P<0.05),且在500 mg·L-1处理中幼苗茎部的铅质量分数最高(39.42 μg·g-1)。各处理幼苗叶片的铅质量分数均低于对照植株,但差异均不显著(P>0.05)。
2.3不同铅胁迫质量浓度下幼苗各部位铅离子转运率
根据图3A可知:随铅胁迫质量浓度的升高,各处理植株的叶/茎离子转运率急剧下降,均显著低于对照值(P<0.05);在各试验处理中,500 mg·L-1处理植株的叶/茎离子转运率显著低于其他处理(P<0.05),其他处理之间差异不显著(P>0.05)。

图2 铅胁迫对刺槐幼苗根(A),茎(B),叶(C)铅离子质量分数的影响Figure 2 Effect of Pb stress on Pb ion content of root(A),stem(B)and leaf(C)of Robinia pseudoacacia seedlings
在中低质量浓度(≤500 mg·L-1)时,随胁迫质量浓度的升高,幼苗的茎/根离子转运率显著上升(P<0.05);当铅质量浓度大于500 mg·L-1时,随铅质量浓度的增大,离子转运率逐渐降低,其中1 000,1 500 mg·L-1处理的植株茎/根离子转运率显著低于500 mg·L-1处理(P<0.05),但两处理之间差异不显著(P>0.05)(图3B)。
与茎/根离子转运相似,随着铅处理质量浓度增加,幼苗的(叶+茎)/根离子转运率在中低质量浓度(≤500 mg·L-1)阶段急剧升高,250,500 mg·L-1两处理的离子转运率均显著高于对照值(P<0.05);随铅处理质量浓度进一步增大,幼苗(叶+茎)/根离子转运率急剧降低,显著低于500 mg·L-1处理值与对照值(P<0.05),但高质量浓度处理之间差异不显著(P>0.05)(图3C)。
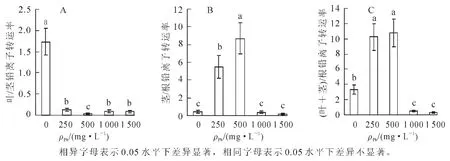
图3 铅胁迫对刺槐幼苗根、茎、叶间铅离子转运率的影响Figure 3 Effect of Pb stress on Pb ion transfer ratio between root,stem and leaf of Robinia pseudoacacia seedlings
2.4铅胁迫条件下幼苗生长特性与铅离子质量分数、转运率的相关性
在刺槐幼苗干质量、地径及苗高与幼苗根铅离子质量分数、茎铅离子质量分数、叶铅离子质量分数、根茎转运率、茎叶转运率、地上地下转运率之间进行相关分析。从表1可以看出:铅胁迫幼苗生长与植物组织铅质量分数、铅离子转运特性存在相关性。幼苗干物质质量与根系铅离子质量分数、叶铅离子质量分数及茎叶转运呈显著性相关(P<0.05)或极显著性相关(P<0.01);幼苗地径生长与根系铅离子质量分数、叶铅离子质量分数、茎叶转运及(叶+茎)/根离子转运(地上地下转运)呈显著性相关(P<0.05);苗高生长仅与叶铅离子质量分数呈显著性相关(P<0.05),可能与叶片铅离子影响生长素形成,进而影响细胞延长,限制苗高生长有关。
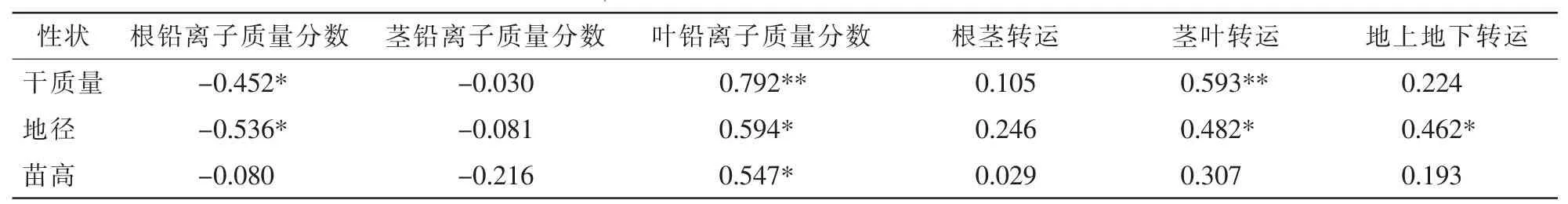
表1 铅胁迫条件下刺槐幼苗干质量、地径及苗高与幼苗根铅离子质量分数、茎铅离子质量分数、叶铅离子质量分数、根茎转运率、茎叶转运率、地上地下转运率之间的皮尔逊指数Table 1 Pearson correlation of dry weight,stem diameter and height,respectively,to Pb ion contens of root,stem and leaf and Pb ion transfer ratios of root to stem,stem to leave,and root to both stem and leaf in Robinia pseudoacacia seedlings under Pb stress
2.5铅胁迫条件下铅离子质量分数及转运特性对刺槐幼苗生长的影响
幼苗干物质是植株生物量积累最直接的指标,也是植物生理功能的最佳体现;幼苗高生长与地径生长直接反应植株纵向与横向发育的状况,体现出植物细胞延长生长与分裂的能力。同时,刺槐幼苗上述3个生长指标分别受幼苗根铅离子质量分数、茎铅离子质量分数、叶铅离子质量分数、根茎转运率、茎叶转运率、地上地下转运率等6个指标影响。为了评估各因素对铅胁迫条件下刺槐幼苗生长的影响程度,就对幼苗干质量、苗高、地径与上述6个指标分别进行多元回归分析。然后,根据各指标标准回归系数的绝对值来比较其对铅胁迫植株的影响程度。
由表2可发现:对刺槐幼苗的生物量积累影响最大的因素为根系铅质量分数,次之为叶片铅质量分数,茎部铅离子质量分数影响最小。同时,从组织的铅离子转运、分配来说,地上地下转运对植物干物质积累影响最大,茎叶转运次之,而根茎间的铅离子转运影响最小。
影响刺槐幼苗地径生长的最大因素是茎部铅离子质量分数,次之为叶片铅质量分数,影响最小的根系铅质量分数。此外,根茎间铅离子转运对幼苗横向生长影响最大,地上地下转运次之,而茎叶转运影响最小。
对刺槐幼苗高生长产生影响的最大因素是茎部铅离子质量分数,根系铅质量分数为次要因素,叶片铅质量分数影响最小。此外,根茎间铅离子转运对幼苗高生长影响最大,次要因素为地上地下转运,而茎叶转运对苗高生长影响最小。
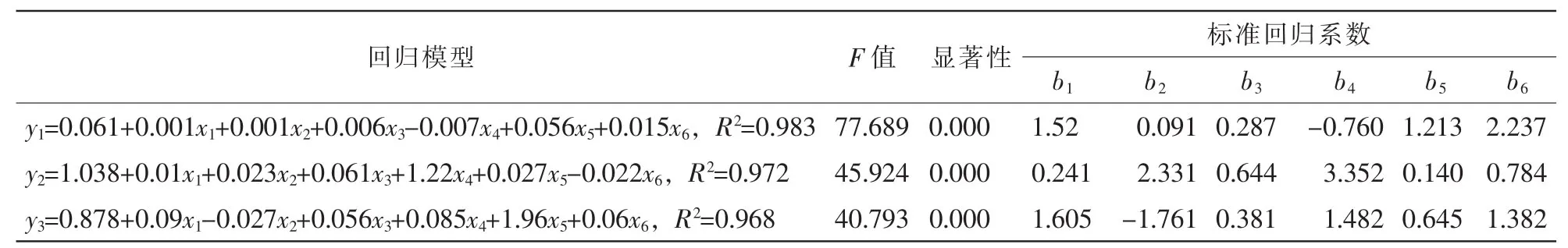
表2 铅胁迫下刺槐幼苗干质量、苗高与地径和幼苗根铅离子质量分数等6个特征指标之间的多元回归分析Table 2 Multiple regression analysis of dry weight,height and stem diameter in relation to six parameters of contents and tansfer ratios of lead ion in Robinia pseudoacacia seedlings under Pb stress
3 讨论
3.1不同铅离子质量浓度对刺槐幼苗生长的影响
在本研究中,250 mg·L-1的铅离子质量浓度处理对刺槐幼苗的苗高、地径、鲜质量有一定促进作用。这与铅胁迫下小麦Triticum aestivum[17],巨桉Eucalyptus grandis[18]幼苗的变化规律相似。有研究者认为[19],在低质量浓度的铅胁迫下,植物的叶绿素质量分数、过氧化物酶(POD)和超氧化物歧化酶(SOD)等均有所增加,增加了植物对逆境的适应能力,促进了植物的生长发育。
在中高质量浓度(≥500 mg·L-1)下,随铅离子质量浓度的增加,刺槐幼苗的茎高、地径以及植株的鲜质量、干质量大体呈渐下降趋势。可能是随着土壤中铅离子质量浓度急剧增加,植物体内富集的铅离子质量浓度超过了植物本身的耐受程度,促使叶绿体瓦解,叶绿素质量分数降低[20],抗氧化性酶体系活性降低[10],使得植物受到重金属毒害,抑制了刺槐幼苗的生长。
3.2刺槐铅离子转运特性对其耐受铅胁迫的影响
植物通过抑制重金属离子跨膜吸收等机制,将大量重金属离子阻止在根部,限制重金属向地上部位运输,降低体内的重金属离子质量分数,从而减轻植物本身受到的伤害[21]。分析刺槐各组织间铅离子转运特性,发现叶与茎的离子转运率逐步降低,而地上部与地下部、茎与根的转运率先升后降,500 mg· L-1时离子转运率达到峰值。刺槐铅离子主要富集在根部,部分聚集在茎,向叶部转移较少,其富集模式与黑麦草Lolium perenne,高羊茅Festuca elata,鹅观草Roegneria kamoji等比较相似[22-23],这样可减轻叶片光合系统遭到损伤,维持受胁迫植株的光合生产能力,提高植株对铅胁迫的耐受能力与存活率。500 mg·L-1是刺槐幼苗的一个关键阈值,当铅胁迫质量浓度大于阈值时,铅离子在组织内部的存储、转运模式发生变化,起到了主动防御的效果。在高质量浓度条件下,铅离子在刺槐根部富集,同时减少向地上部分的转运,这是刺槐幼苗对铅胁迫伤害的一种主动防御机制,也是对铅胁迫的主要耐受机制。
3.3铅胁迫条件下刺槐幼苗生长的综合影响机制
通过多元回归发现,根系铅质量分数与地上/地下部分离子转运率对刺槐幼苗的生物量积累影响最大。这种模式应该是与根系积累铅离子,然后转运到叶片,破坏叶绿体,损伤光合结构与功能,最终影响有机物质积累相关。影响刺槐幼苗地径与苗高生长的最大因素均是茎部铅离子质量分数与根/茎间铅离子转运率。这种模式与根部吸收铅离子,转运到茎部积累,损伤形成层的细胞分生功能与茎部细胞的延长功能,从而限制了刺槐幼苗地径与苗高生长有关。
[1]AMEH E G,AKPAH F A.Heavy metal pollution indexing and multivariate statistical evaluation of hydrogeochemis-try of River PovPov in Itakpe Iron-Ore mining area,Kogi State,Nigeria[J].Adv Appl Sci Res,2011,2(1):33-46.
[2]CAPUANA M.Heavy metals and woody plants-biotechnologies for phytoremediation[J].IFor-Biogeosci For,2011,4 (1):7-15.
[3]李瑞琴,于安芬,白滨,等.甘肃中部高原露地菜田土壤重金属污染及潜在生态风险分析[J].农业环境科学学报,2013,32(1):103-110. LI Ruiqin,YU Anfen,BAI Bin,et al.Analysis on current situation and potential ecological risk and the characteristic of heaven metals pollution of soil in the central plateau of Gansu Province,China[J].J Agro-Environ Sci,2013,32 (1):103-110.
[4]邱清华,邓绍云,黄娟,等.铅胁迫对十字花科种子萌发及幼苗生长的影响[J].中国农学通报,2010,26 (18):175-179. QIU Qinghua,DENG Shaoyun,HUANG Juan,et al.Study on the influence of lead stress on the mustard family’s seed sprouting and seedling growing[J].Chin Agric Sci Bull,2010,26(18):175-179.
[5]韩豫,曹莹,王绍斌,等.铅胁迫对花生生理生化特性的影响[J].花生学报,2007,36(1):24-27. HAN Yu,CAO Ying,WANG Shaobin,et al.Effects of lead stress on the physiological and biochemical characteristics of peanut[J].J Peanut Sci,2007,36(1):24-27.
[6]刘法彬,孙永林.铅污染对黄瓜种子萌发及叶片生理指标的影响[J].化学与生物工程,2008,25(10):60-63. LIU Fabin,SUN Yonglin.Effects of lead pollution on seed germination and physiological indexes in leaves of cucumber[J].Chem Bioeng,2008,25(10):60-63.
[7]张晓雯,陈世华.铅胁迫对绿豆种子萌发及幼苗生长的影响[J].安徽农业科学,2008,36(34):14859-14882. ZHANG Xiaowen,CHEN Shihua.Effects of lead stress on seed germination and seedling growth of mung bean[J].J Anhui Agric Sci,2008,36(34):14859-14882.
[8]刘明久,李淦,肖万红.Pb2+胁迫对水稻种子萌发及幼苗生长影响的研究[J].种子,2006,25(12):44-46. LIU Mingjiu,LI Gan,XIAO Wanhong.Study on the effect of seed germination and seedling growth of rice under Pb2+stress[J].Seed,2006,25(12):44-46.
[9]张义贤,付亚萍,肖志华,等.铅胁迫对不同基因型谷子幼苗生理特性及基因组DNA多态性的影响 [J].农业环境科学学报,2013,32(3):478-484. ZHANG Yixian,FU Yaping,XIAO Zhihua,et al.Effects of Pb2+stress on physiological characteristics and DNA polymorphism of genome in difference genotypes foxtail millet[J].J Agro-Environ Sci,2013,32(3):478-484.
[10]周芙蓉,王进鑫,杨楠,等.干旱和铅交互作用对侧柏幼苗生长及抗氧化酶活性的影响[J].林业科学,2013,49(6):172-177. ZHOU Furong,WANG Jinxin,YANG Nan,et al.Interaction of drought and Pb on growth and antioxidant enzyme activities of Platycladus orientalis seedlings[J].Sci Silv Sin,2013,49(6):172-177.
[11]袁菊红,胡绵好.铅胁迫下硒处理的彩叶草根系和叶片的SEM/XRD光谱学分析[J].园艺学报,2013,40 (3):562-570. YUAN Juhong,HU Mianhao.SEM and XRD analyses of the roots and leaves of coleus with selenium supplements under lead stress[J].Acta Hortic Sin,2013,40(3):562-570.
[12]WU Xiao,LIU Chao,QU Chunxiang,et al.Effects of lead on activities of photochemical reaction and key enzymes of carbon assimilation in spinach chloroplast[J].Biol Trac Elem Res,2008,126(1):269-279.
[13]王慧忠,何翠屏.重金属离子胁迫对草坪草根系生长及其活力的影响[J].中国草地,2002,24(3):55-59. WANG Huizhong,HE Cuiping.Effects of the heavy metal stress on root growth and vigour of turfgrass[J].Grassland China,2002,24(3):55-59.
[14]屈冉,孟伟,李俊生,等.土壤重金属污染的植物修复[J].生态学杂志,2008,27(4):626-631. QU Ran,MENG Wei,LI Junsheng,et al.Research progress on phytoremediation of heavy metal contaminated soil [J].Chin J Ecol,2008,27(4):626-631.
[15]马敏,龚惠红,邓泓.重金属对8种园林植物种子萌发及幼苗生长的影响[J].中国农学通报,2012,28(22):206-211. MA Min,GONG Huihong,DENG Hong.Effects of heavy metal stress on seed germination and seedling growth of eight urban plants[J].Chin Agric Sci Bull,2012,28(22):206-211.
[16]何晓良,蔡卓,邱霞琳,等.铅胁迫对狮子草可溶性蛋白含量的影响[J].广西大学学报:自然科学版,2013,38(3):550-553. HE Xiaoliang,CAI Zhuo,QIU Xialin,et al.Effects of lead stress on soluble protein content of Peristrope japonica (Thunb)Brem[J].J Guangxi Univ Nat Sci Ed,2013,38(3):550-553.
[17]康吉利,曾志军,刘玉佩.铅胁迫对小麦种子萌发及幼苗生长的影响[J].广西农业科学,2009,40(2):144 -146. KANG Jili,ZENG Zhijun,LIU Yupei.Effects of lead(Pb2+)stress on seed germination and seedlings growth of wheat [J].Guangxi Agric Sci,2009,40(2):144-146.
[18]林晓倩,张健,杨万勤,等.3种土壤类型下铅胁迫对巨桉幼苗的影响[J].林业科学,2013,49(1):1-6. LIN Xiaoqian,ZHANG Jian,YANG Wanqin,et al.Effects of Pb stress on Eucalyptus grandis seedlings in three types of soil[J].Sci Silv Sin,2013,49(1):1-6.
[19]李珊,程舟,杨晓伶,等.铅、锌胁迫对栝楼幼苗生长及抗逆生理因子的影响[J].生态学杂志,2008,27 (2):278-281. LI Shan,CHENG Zhou,YANG Xiaoling,et al.Effects of Pb and Zn stress on Trichosanthes kirilowii seedlings growth and stress-resistance physiological factors[J].Chin J Ecol,2008,27(2):278-281.
[20]明华,曹莹,胡春胜,等.铅胁迫对玉米光合特性及产量的影响[J].玉米科学,2008,16(1):74-78. MING Hua,CAO Ying,HU Chunsheng,et al.Effect of lead stress on the photosynthetic characteristics and yield of maize[J].J Maize Sci,2008,16(1):74-78.
[21]AKINCI I E,AKINCI S,YILMAZ K.Response of tomato(Solanum lycopersicum L.)to lead toxicity:growth,element uptake,chlorophyll and water content[J].Afr J Agric Res,2010,5(6):416-423.
[22]杨明琰,梁语燕,曾德榜,等.铅胁迫对黑麦草Pb富集特性及生理代谢的影响[J].西北农林科技大学学报:自然科学版,2014,42(12):97-101. YANG Mingyan,LIANG Yuyan,ZENG Debang,et al.Effects of lead stress on accumulation capacity and physiological metabolism of ryegrass[J].J Northwest A&F Univ Agric Univ Nat Sci Ed,2014,42(12):97-101.
[23]李慧芳,王瑜,袁庆华,等.铅胁迫对禾本科牧草生长、生理及Pb2+富集转运的影响[J].草业学报,2015,24 (9):163-172. LI Huifang,WANG Yu,YUAN Qinghua,et al.Effects of lead stress on growth,physiology,and lead ion accumulation and transportation in gramineous forages[J].Acta Pratac Sin,2015,24(9):163-172.
Growth and ion transport with Pb stress in Robinia pseudoacacia seedlings
ZHOU Jian1,2,JIANG Zeping1,WEI Yuan1
(1.Research Institute of Forestry,Chinese Academy of Forestry,Beijing 100091,China;2.School of Horticulture and Landscape Architecture,Institute of Science and Technology of Henan,Xinxiang 453003,Henan,China)
To explore Robinia pseudoacacia(Black Locust)tolerance and mechanism to lead(Pb)stress,R. pseudoacacia seedlings were first watered with solutions of Pb(NO3)2dissolved in distilled water.Then,properties of growth,lead concentration(μg·g-1),and ion transfer in stressed seedlings were analyzed using multivariate linear regression analysis,Ducan multiple comparison and single factor test design with treatments of 250,500,1 000 and 1 500 mg·L-1and 3 replications.Experimental results showed that 250 mg·L-1of Pb(NO3)2solution slightly promoted height,stem diameter,and seedling dry and fresh weights,but these decreased when lead concentration was subsequently increased.Lead content significantly increased(P<0.05)with accumulation found in roots;seedling stems showed a lead increase(P<0.05)and a subsequent decrease(P<0.05).In stressed seedlings leaf concentration declined in these treatments of 250 to 1 500 mg·L-1.In addition,the lead ion transfer-ratio for stem to leaf significantly declined(P<0.05).However,as lead concentration increased,the root to stem and root to both stem and leaf concentration increased(P<0.05)and then decreased(P<0.05).The multivariate linear regression analysis,showed that the most important factors affecting accumulation of dry biomass were 1)lead concentration in roots and 2)ion transfer ratio of root to both stem and leaf.Also in stressed seedlings lead concentration in stems and ion transfer ratio of root to stem the most significantly affected(P<0.05)growth of height and of stem diameter.It was drawn that lead accumulation in root and inhibition of its transfer could prevent leaves from damage deriving from lead stress,and they would be the tolerance mechanism to lead stress for Robinia pseudoacacia seedlings.[Ch,3 fig.2 tab.23 ref.]
botany;lead(Pb)stress;Robinia pseudoacacia;seedling;growth;ion transport
S718.43
A
2095-0756(2016)05-0742-07
10.11833/j.issn.2095-0756.2016.05.003
2015-11-02;
2016-01-06
“十二五”国家科技支撑计划项目(2012BAC09B03);河南省科技攻关项目(162102110133)
周建,副教授,博士,从事园林植物逆境生理生态研究。E-mail:zj200102@163.com。通信作者:魏远,助理研究员,博士,从事木本植物逆境研究。E-mail:weiyuan_caf@126.com
浙 江 农 林 大 学 学 报,2016,33(5):742-748
Journal of Zhejiang A&F University
