一种新ω-3脂肪酸脱饱和酶的克隆表达和活性鉴定
2016-10-21梅甜甜陈海琴郝光飞顾震南陈卫陈永泉
梅甜甜,陈海琴,郝光飞,顾震南,陈卫,陈永泉
(江南大学 食品学院,食品科学与技术国家重点实验室,江苏 无锡,214122)
一种新ω-3脂肪酸脱饱和酶的克隆表达和活性鉴定
梅甜甜,陈海琴*,郝光飞,顾震南,陈卫,陈永泉
(江南大学 食品学院,食品科学与技术国家重点实验室,江苏 无锡,214122)
ω-3脂肪酸脱饱和酶催化ω-6多不饱和脂肪酸(PUFAs)转化为ω-3 PUFAs,对ω-3长链多不饱和脂肪酸(LC-PUFAs)的合成至关重要。为了实现在常温下发酵生产ω-3 LC-PUFAs(主要是二十碳五烯酸,EPA),根据现有常温下偏好催化20C PUFAs的ω-3脂肪酸脱饱和酶序列,从GenBank数据库筛选出与之高度相似的序列并进行生物信息学分析。为了确定序列的生物活性,进一步在酿酒酵母系统中进行重组表达,通过外源添加不同碳链长度的脂肪酸底物,测定重组酿酒酵母转化子在28 ℃和12 ℃下对不同脂肪酸的转化率。结果显示,新筛选序列编码的蛋白oAiFADS17既能催化18C PUFAs,又能催化20C PUFAs,尤其偏好催化二十碳四烯酸(AA)转化为EPA。oAiFADS17蛋白在28 ℃下对各种底物的转化率均高于12 ℃下的转化率,其中对AA的转化率达到46.3%。该研究成功测定了oAiFADS17蛋白对不同脂肪酸底物的转化率,得到了一种新的常温偏好催化20C PUFAs的ω-3脂肪酸脱饱和酶,为构建高产EPA的基因工程菌株及EPA的工业化生产奠定了理论基础。
ω-3脂肪酸脱饱和酶;转化率;二十碳五烯酸(EPA)
ω-3长链多不饱和脂肪酸(LC-PUFAs)主要包括二十碳五烯酸(Eicosapentaenoic acid, EPA, C20∶5)和二十二碳六烯酸(Docosahexaenoic acid, DHA, C22∶ 6)。EPA和DHA可以促进哺乳动物大脑发育以及神经组织的形成和修复[1],并对哮喘、癌症、抑郁、肥胖、免疫紊乱以及心血管疾病具有十分显著的预防作用[2-3]。但人体自身不能从源头合成EPA和DHA,需要从富含ω-3 LC-PUFAs的食物(如深海鱼)中摄取。然而由于大肆捕捞以及海洋环境的污染,深海鱼的供应已无法满足日益增长的市场需求[4-6]。
自然界中ω-3 LC-PUFAs的合成起始于亚油酸(linoleic acid, LA, C18∶2)和γ-亚麻酸(γ-linoleic acid, ALA, C18∶3),经过一系列脱饱和酶和延长酶的催化作用,最终转化生成EPA和DHA。ω-3脂肪酸脱饱和酶(EC 1.14.19.25)位于内质网膜上,是ω-3 LC-PUFAs合成中的关键酶[7]。研究发现,不同来源的ω-3脂肪酸脱饱和酶对不同碳链长度的脂肪酸具有不同的催化效率。早期研究发现来源于植物的ω-3脂肪酸脱饱和酶(FAD7, FAD8等)只能催化18C PUFAs[8]。而来源于秀丽隐杆线虫(Caenorhabditiselegans)的ω-3脂肪酸脱饱和酶FAT 1则能同时催化18C和20C PUFAs,但对20C底物的催化活性较低[9]。2004年PEREIRA等人发现来源于异枝水霉(Saprolegniadiclina)的ω-3脂肪酸脱饱和酶SDD17对18C PUFAs没有催化活性,却能将二十碳四烯酸(Arachidonic acid, AA, C20∶4)催化为EPA,转化率为25.9%[10]。同样的,来源于致病疫霉(Phytophthorainfestans)的ω-3脂肪酸脱饱和酶OPIN 17也不能以18C PUFAs为底物,但是对AA的转化率达到了30.94%[11]。2013年,XUE等人从瓜果腐霉(Pythiumaphanidermatum)、大豆疫霉菌(Phytophthorasojae)和栎树猝死病菌(Phytophthoraramorum)中分离出3种ω-3脂肪酸脱饱和酶——PaD 17、PsD 17和PrD 17,这3种酶都具有催化18C和20C PUFAs的能力,对20C 脂肪酸AA的转化率分别为56%、47%和36%,并且这3种酶适应在28 ℃下进行催化反应[12]。这一类在28 ℃下偏好催化20C脂肪酸的ω-3脂肪酸脱饱和酶对ω-3 LC-PUFAs尤其是EPA的合成至关重要。本研究结合基因搜索和基因工程重组表达,筛选出了一种新的偏好催化20C脂肪酸的ω-3脂肪酸脱饱和酶。新ω-3脂肪酸脱饱和酶的发现为研究这类脱饱和酶的偏好性提供了新的资源,也为ω-3脂肪酸脱饱和酶的应用提供了更多选择,同时为EPA的工业化生产奠定了理论基础。
1 材料与方法
1.1材料
1.1.1菌株和载体
大肠杆菌(Escherichiacoli,E.coli)TOP 10由本实验室保存。表达载体pYES2/NT C与尿嘧啶缺陷型酿酒酵母INVSc 1购于Invitrogen公司,现由本实验室保存。
1.1.2试剂
KOD plus高保真DNA聚合酶购自Toyobo公司;限制性内切酶EcoR I和XhoI、T4DNA连接酶、Prime Script RT reagent kit试剂盒均购自TaKaRa生物技术公司(大连);凝胶回收试剂盒购自Fermentas公司;PCR产物纯化试剂盒购自生物工程(上海)股份有限公司;TaqDNA聚合酶购自北京天根生化科技有限公司;BCA蛋白测定试剂盒购于上海碧云天公司;SYBR Green PCR Master Mix购自美国Bio-Rad公司;脂肪酸标准品LA、十八碳三烯酸(GLA)、二十碳三烯酸(DGLA)、AA购自美国Sigma公司。
1.1.3培养基及培养条件
LB培养基(g/L):胰蛋白胨 10,酵母提取物 5,氯化钠 10,pH 7.0,固体培养基添加1.5%琼脂粉,用于大肠杆菌的培养,培养温度37 ℃,摇床转速200 r/min。
YPD培养基(g/L):酵母抽提物 10,蛋白胨 20,葡萄糖 20,pH 7.0,固体培养基添加1.5%琼脂粉,用于酿酒酵母的培养,培养温度28 ℃,摇床转速200 r/min。
SC-U(g/L):酵母氮源(无氨基酸有硫酸铵) 6.7,葡萄糖 20,腺嘌呤、 精氨酸、半胱氨酸、亮氨酸、赖氨酸、苏氨酸、色氨酸、尿嘧啶各0.1,天冬氨酸、组氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、酪氨酸、缬氨酸各0.05,用于转化子扩大培养,培养温度28 ℃,转速200 r/min。诱导培养时更换碳源为10 g/L棉籽糖和20 g/L半乳糖,其余成分与SC-U培养基相同。
1.1.4实验所用引物
本研究所用引物如表1所示,根据优化得到的oAiFADS17基因序列设计引物扩增目的基因片段,同时引入EcoR I和XhoI酶切位点(表中下划线所示)。根据oAiFADS17基因序列保守区设计RT-qPCR引物,使用酿酒酵母18S rRNA通用引物作为内参[13]。

表1 本研究所用引物
注:1)表中下划线表示引入的酶切位点,括号中的字母表示限制性酶切位点。
1.2实验方法
1.2.1oAiFADS17基因的获得
利用BLAST从Genbank数据库筛选到一段来自真菌媒介丝囊菌(Aphanomycesinvadans)的基因序列(GenBank accession No. XM_008870610),与5种已知偏好催化20C PUFAs的ω-3脂肪酸脱饱和酶(SDD17, OPIN 17, PaD17, PsD17, PrD17)具有较高的同源性。将这段基因编码的蛋白与上述5种ω-3脂肪酸脱饱和酶序列在Clustal W2中进行比对,同时利用TMHMM软件预测此段序列编码蛋白的跨膜域。进一步构建含此段序列的表达载体,首先利用Genscript Optimum GeneTMsystem软件对其核酸序列进行密码子优化,人工合成优化后的基因序列,命名为oAiFADS17(GenBank accession No. KT371999),再与pUC57-simple载体连接得pUC57-oAiFADS17,保存于大肠杆菌 TOP 10中(金斯瑞,南京,中国)。依据oAiFADS17基因序列设计引物oAiFADS17 F/oAiFADS17 R来扩增目的基因片段。
1.2.2表达载体构建及测序
用限制性内切酶EcoR I和XhoI双酶切oAiFADS17基因片段和pYES2/NT C表达载体,酶切产物经切胶回收纯化后进行连接。取3 μL上述连接产物电转化大肠杆菌TOP 10。挑取阳性转化子,用目的基因引物进行PCR鉴定。PCR产物用1%琼脂糖凝胶电泳验证,条带大小正确的转化子进行测序验证。
1.2.3酿酒酵母转化及重组转化子诱导蛋白表达
采用PEG/LiAc法[14]转化酿酒酵母,从平板上挑取生长状态良好的阳性克隆接种到5 mL YPD培养基中,30 ℃ 200 r/min培养48 h。提取质粒,用通用引物T7/T7 terminator(结合于多克隆位点上下游)进行PCR验证并测序。挑取测序正确的酿酒酵母转化子单菌落接种于SC-U培养基(含空质粒的酿酒酵母转化子为阴性对照,每组3个平行),28 ℃培养48 h。测量OD600值,转接入SC-U诱导培养基,调整OD600值达到0.4。同时根据需要,向培养基中添加不同碳链长度PUFAs(见2.3.3),28 ℃/12 ℃培养48 h后收集菌体。
1.2.4酿酒酵母转化子转录水平测定
用Trizol提取酿酒酵母菌体总RNA。以1 μg总RNA为模板,根据Prime Script RT reagent kit(TaKaRa, Otsu, Shiga, Japan)试剂盒说明进行操作,获得cDNA。使用Bio-Rad CFX ConnectTMsystem 按照 iTaq Universal SYBR Green Supermix说明进行RT-qPCR反应。反应体系为:8 μL无酶水,10 μL iTaq Universal SYBR Green Supermix,0.5 μL q-oAiFADS17 F,0.5 μL q-oAiFADS17 R,1 μL cDNA模板,总体积20 μL。以酿酒酵母的18S rRNA作为内参基因,根据2-△△Ct法计算基因的相对转录水平,所有实验进行3次重复。(注明:①ΔCt=待测样品目的基因Ct值-待测样品内参Ct值;②ΔCt=实验组ΔCt-对照组ΔCt。)
1.2.5蛋白表达水平测
收集诱导后的菌体2 mL,10 000 r/min离心弃去上清。加入适量体积的裂解液(50 mmol/L Na2HPO4-NaH2PO4, 1 mmol/L EDTA, 5% 1,2,3-丙三醇, 1 mmol/L PMSF, pH 7.4)使OD600值达到100,然后加入等体积的0.5 mm酸洗玻璃珠,置于漩涡振荡器振荡30 s后置于冰上30 s,如此重复4次后于13 000 r/min离心10 min,得到的上层悬液转移至新的1.5 mL离心管中,用BCA试剂盒测定蛋白浓度后,以100 μg总蛋白的上样量进行SDS-PAGE以及Western Blot[15]。
1.2.6底物特异性测定
收集诱导后的酿酒酵母菌体,真空冷冻干燥后置于研钵中充分研磨粉碎。称取20 mg菌体粉末采用脂肪酸甲酯化法[16]进行提取,加入C15∶0(用于标定链长≤18C的脂肪酸)和C21∶0(用于标定链长>18C的脂肪酸)各100 μL作为内标。得到的脂肪酸甲酯使用GC-MS(Shimadzu Co., Japan)检测[17]。色谱柱为Rtx-Wax(30 m×0.25 mm, 0.25 μm),汽化室和检测器温度分别为240 ℃和250 ℃,分流方式进样1 μL,分流比10∶1,载气为氦气。程序升温:初始温度120 ℃保持3 min,以5 ℃/min升到190 ℃,再以4 ℃/min升到220 ℃,保持20 min。
2 结果与讨论
2.1oAiFADS17序列的生物信息学分析
oAiFADS17基因来自真菌媒介私囊菌,全长1095 bp,编码364个氨基酸,蛋白理论大小约41 kDa。为了考证oAiFADS17编码的蛋白oAiFADS17是否与ω-3脂肪酸脱饱和酶同源,将其与5种已知的ω-3脂肪酸脱饱和酶氨基酸序列进行了比对(图1)。
利用Clustal W2工具的比对结果表明,oAiFADS17与已知ω-3脂肪酸脱饱和酶的同源性高于59%,且具有与其他ω-3脂肪酸脱饱和酶一样的3个高度保守的组氨酸保守区HX3HH,HX2HH,HX2HH。进一步通过脂肪酸拓扑结构模型分析发现(图2),除位于亲水区的3个组氨酸保守区外,oAiFADS17还具有位于疏水区的4个跨膜螺旋和2个膜内结构域,共同构成了ω-3脂肪酸脱饱和酶的催化活性中心,理论上符合ω-3脂肪酸脱饱和酶的基本特征。
2.2含oAiFADS17基因重组酿酒酵母菌的构建
利用oAiFADS17 F/oAiFADS17 R引物通过PCR扩增得到oAiFADS17基因序列,将其连接至pYES2/NT C载体,命名为pYES2-oAiFADS17。将重组质粒以及空质粒pYES2/NT C转化入酿酒酵母INVSc 1,挑取阳性转化子,利用通用引物T7/T7 terminator进行PCR验证,验证结果如图3所示,2、3、4号转化子扩增得到的片段在1 463 bp(1 095 bp+368 bp)左右,而转入空质粒pYES2/NT C的1号转化子在368 bp左右出现条带,与理论大小一致。进一步通过测序验证表明1号为转入空质粒pYES2/NT C的转化子,2、3、4号为转入重组质粒pYES2-oAiFADS17的转化子。将得到1-4号转化子分别命名为INVSc 1(pYES2/NT C),INVSc 1 (pYES2-oAiFADS17)-1,INVSc 1 (pYES2-oAiFADS17)-2,INVSc 1 (pYES2-oAiFADS17)-3。

图1 真菌媒介私囊菌的ω-3脂肪酸脱饱和酶(oAiFADS17)与偏好催化20C PUFAs的ω-3脂肪酸脱饱和酶氨基酸序列比对Fig.1 Multiple equence alignment of amino acids for ω-3 desaturases from Aphanomyces invadans (oAiFADS17) and which prefer to convert 20C PUFAs. PrD17:ω-3 desaturases from Phytophthora ramorum(GenBank accession No. FW362214.1); PsD17:ω-3 desaturases from Phytophthora sojae(GenBank accession No. FW362213.1); OPIN17:ω-3 desaturases from Phytophthora infestans(GenBank accession No. XM 002902553.1); PaD17:ω-3 desaturases from Pythium aphanidermatum(GenBank accession No. FW362186.1); SDD17:ω-3 desaturases from Saprolegnia diclina (GenBank accession No. AY373823.1). Black bars show the identical amino acid residues. Deletions are indicated by dashes. Boxes represent three typical histidine motifs

图2 ω-3脂肪酸脱饱和酶oAiFADS17的拓扑结构模型Fig.2 Topology model of ω-3 desaturase oAiFADS17 Cylinders A-F indicate transmembrane helices, and the black solid circles indicate three conserved histidine-rich motifs. Prediction of transmembrane helices and topology was performed with TMHMM, HMMTOP and TOP-PRED . The predicted topology model was in qualitative agreement with other membrane-bound enzymes

M-Marker; 1-对照, INVSc 1 (pYES2/NT C); 2-INVSc 1 (pYES2-oAiFADS17)-1; 3-INVSc 1 (pYES2-oAiFADS17)-2; 4-INVSc 1 (pYES2-oAiFADS17)-3图3 酿酒酵母转化子PCR验证Fig.3 PCR verification of yeast transformation
2.3 oAiFADS17脱饱和酶重组酿酒酵母菌的活性验证
2.3.1目的基因oAiFADS17转录水平测定
将筛选获得的3株转化子于SC培养基中28 ℃培养2 d后收集菌体,通过RT-qPCR方法测定目的基因在转化子中的转录水平。转录水平分析结果表明在转入空质粒的酿酒酵母转化子INVSc 1(pYES2/NT C)中未检测到目的基因转录,而在转入重组质粒的3个酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)中,目的基因均有显著表达(图4)。

1-Control, INVSc 1 (pYES2/NT C); 2-INVSc 1 (pYES2-oAiFADS17)-1; 3-INVSc 1 (pYES2-oAiFADS17)-2; 4-INVSc 1 (pYES2-oAiFADS17)-3图4 oAiFADS17基因相对表达量Fig.4 The relative expression level of oAiFADS17 ( **:P<0.001. All data are presented as mean ± SD.)
2.3.2目标蛋白oAiFADS17表达水平测定
利用Western blot对重组菌株中目的蛋白的表达水平进行分析,具体方法参见1.2.5。转入重组质粒的3个酿酒酵母转化子均在45 kDa附近(表达载体上带有3.4 kDa His标签,故检测蛋白理论分子量大小约45 kDa)检测到目标蛋白的杂交条带(图5),该蛋白的分子量大小与其理论值相符,说明在酿酒酵母3个转化子INVSc 1 (pYES2-oAiFADS17)中目的蛋白正确表达。

1-对照, INVSc 1 (pYES2/NT C); 2-INVSc 1 (pYES2-oAiFADS17)-1; 3-INVSc 1 (pYES2-oAiFADS17)-2; 4-INVSc 1 (pYES2-oAiFADS17)-3图5 酿酒酵母转化子的蛋白表达水平Fig.5 The protein expression level of yeast transformants
2.3.3酿酒酵母重组转化子催化特性分析
为了检测重组酿酒酵母菌株对AA的转化率,将3个酿酒酵母转化子在分别添加了0.05 ,0.1,0.2 mmol/L AA的诱导培养基中28 ℃培养2 d,收集菌体后检测其脂肪酸组成。气相色谱图显示,转入目的基因的酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)在37.5 min处出现了信号峰(图6c),与EPA标准品出峰时间一致(图6a),而转入空质粒的对照组酿酒酵母INVSc 1 (pYES2/NT C)在相应的保留时间无信号(图6b)。该信号峰与EPA标准品的出峰时间完全一致。进一步将该峰对应物质的质谱图与EPA标准品质谱图(图7)比对,两者一致,且均在m/z79、m/z91、m/z119处出现强的离子峰,确认此物质为EPA。以上GC-MS分析结果表明,酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)能将ω-6系的AA转化为ω-3系的EPA,oAiFADS17具有ω-3脂肪酸脱饱和酶活性。当外源添加不同浓度的AA时(表2),各转化子对AA的转化率非常接近。在AA浓度为0.05 mmol/L时,各转化子的转化率为36%~46%不等,而随着AA浓度增高,各转化子对AA的转化率有所降低,原因可能是由于AA量达饱和导致。为了进一步探究oAiFADS17对不同链长脂肪酸的催化特性,选取酿酒酵母INVSc 1 (pYES2-oAiFADS17)-3以及对照组INVSc 1 (pYES2/NT C)转化子在添加不同碳链长度脂肪酸底物的培养基中培养,添加方式包括单一底物(0.2 mmol/L LA、GLA、DGLA、AA)及双底物(0.1 mmol/L LA+0.1 mmol/L AA)形式,培养温度分别为28 ℃和12 ℃。在两种培养温度下,酿酒酵母INVSc 1 (pYES2-oAiFADS17)-3对20CPUFAs的转化率明显高于18CPUFAs,且对AA转化率最高,而在添加双底物(LA+AA)条件下,转化子对20C底物AA的偏好性更强,由此可知oAiFADS17偏好催化20CPUFAs,而对照组却不能转化任何一种底物。同时,酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)在28 ℃下对各种脂肪酸的转化率均高于12 ℃下的转化率(表3),说明oAiFADS17在28 ℃下活性较高。综上所述,oAiFADS17是一种在28 ℃对AA有较高催化活性的ω-3脂肪酸脱饱和酶。

a-EPA standard; b-Control, INVSc 1 (pYES2/NT C); c-INVSc 1 (pYES2-oAiFADS17)图6 酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)脂肪酸色谱图Fig.6 Gas chromatogram of fatty acid methyl esters from lipid fraction of INVSc 1 (pYES2-oAiFADS17)
3 结论
本研究从GenBank数据库筛选到一个新ω-3脂肪酸脱饱和酶基因oAiFADS17,并将此基因成功地在酿酒酵母系统中进行了表达。通过外源添加底物,对该基因的转录水平、蛋白表达水平及底物特异性进行了分析。结果表明这种ω-3脂肪酸脱饱和酶(oAiFADS17)在28 ℃下可以催化18C和20C链长的ω-6 PUFAs,尤其偏好催化20C链长底物AA生成具更高附加值的EPA,对AA的转化率达到了46.3%。本研究为ω-3脂肪酸脱饱和酶偏好性的研究提供了新的资源,也为EPA的工业化生产奠定了一定的理论基础。
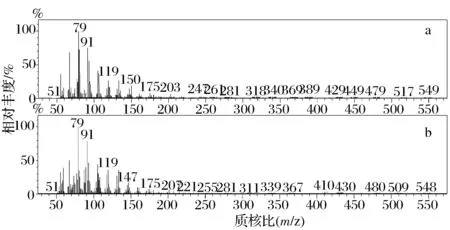
a-Mass spectrum of EPA in INVSc 1 (pYES2-oAiFADS17); b-Mass spectrum of EPA standard图7 EPA质谱图Fig.7 Mass spectrum of the EPA

表2 酿酒酵母转化子INVSc 1 (pYES2-oAiFADS17)对AA的转化率
注:1)Conversion rate is calculated as 100% ×Product/(Product+Substrate).

表3 不同温度下酿酒酵母转化子对不同底物转化率
注:1)Conversion rae is calculated as 100%×Product/(Product+Substrate).
[1]SAKURADANI E,SHIMIZU S.Single cell oil production byMortierellaalpina[J].Journal of Biotechnology, 2009,144(1):31-36.
[2]KHAWAJA O A,GAZIANO J M,DJOUSSE L.N-3 fatty acids for prevention of cardiovascular disease[J]. Current Atherosclerosis Reports,2014,16(11):450.
[3]LORENTE-CEBRIAN S,COSTA A G,NAVAS-CARRETERO S,et al.Role of omega-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases:a review of the evidence [J].Journal of Physiology and Biochemistry,2013, 69(3):633-651.
[4]RACINE R A,DECKELBAUM R J.Sources of the very-long-chain unsaturated omega-3 fatty acids:eicosapentaenoic acid and docosahexaenoic acid [J].Current Opinion in Clinical Nutrition and Metabolic Care, 2007,10(2):123-128.
[5]杜冰,潘力,林炜铁,等.多不饱和脂肪酸发酵的代谢调控育种[J].食品与发酵工业,2000,26(5):52-55.
[6]徐天宇. 利用生物技术生产廿碳五烯酸和廿二碳六烯酸[J].食品与发酵工业,1995,21(1):56-65.
[7]WANG M X,CHEN H Q,GU Z N,et al.ω 3 fatty acid desaturases from microorganisms:structure, function, evolution, and biotechnological use[J].Applied Microbiology and Biotechnology,2013,97(24):10 255-10 262.
[8]TOCHER D R,LEAVER M J,HODGSON P A.Recent advances in the biochemistry and molecular biology of fatty acyl desaturases[J].Progress in Lipid Research,1998,37(2/3):73-117.
[9]MEESAPYODSUK D,REED D W,SAVILE C K,et al.Characterization of the regiochemistry and cryptoregiochemistry of aCaenorhabditiselegansfatty acid desaturase (FAT-1) expressed inSaccharomycescerevisiae[J].Biochemistry-Us,2000,39(39):11 948-11 954.
[10]PEREIRA S L,HUANG Y S,BOBIK E G,et al.A novel omega 3-fatty acid desaturase involved in the biosynthesis of eicosapentaenoic acid[J].Biochemical Journal,2004,378:665-671.
[11]FU Y,FAN X Z,LI X Z,et al.The desaturase OPIN17 fromPhytophthorainfestansconverts arachidonic acid to eicosapentaenoic acid in CHO cells[J].Applied Biochemistry and Biotechnology,2013,171(4):975-988.
[12]XUE Z X,HE H X,Hollerbach D,et al.Identification and characterization of new delta-17 fatty acid desaturases [J].Applied Microbiology and Biotechnology,2013,97(5):1 973-1 985.
[13]CANKORUR-CETINKAYA A,DERELI E, Eraslan S,et al.A novel strategy for selection and validation of reference genes in dynamic multidimensional experimental design in yeast[J].Plos One,2012,7(6):e38 351.
[14]胡又佳,高枫,朱春宝,等.PEG/LiAc转化酵母细胞方法的改进[J].生物技术,1998(5):22-26;49.
[15]CHEN H Q,GU Z N,ZHANG H,et al.Expression and purification of integral membrane fatty acid desaturases [J].Plos One, 2013,8(3). http://dx.doi.org/10.1371/journal pone.0058139.
[16]WANG L,CHEN W, FENG Y,et al.Genome characterization of the oleaginous fungusMortierellaalpina[J].Plos One, 2011,6(12).http://dx.doi.org/10.1371/journal.pone.0028319.
[17]施忠芬,王桂瑛,陈红艳,等.GC-MS法测定宣威火腿中脂肪酸组成[J].食品与发酵工业,2012,38(9):141-145.
Cloning, expression and characterization of a new ω-3 fatty acid desaturase
MEI Tian-tian, CHEN Hai-qin, HAO Guang-fei, GU Zhen-nan,CHEN Wei, CHEN Yong-quan
(School of Food Science and Technology, State Key Laboratory of Food Science and Technology, Jiangnan University,Wuxi 214122,China)
ω-3 fatty acid desaturase can convert ω-6 PUFAs to ω-3 PUFAs, which is significant for the synthesis of ω-3 LC-PUFAs. In order to realize the production of EPA (eicosapentaenoic acid; 20∶5) at 28 ℃, a new gene which shares high sequence similarity with reported ω-3 fatty acid desaturases was screened in GenBank. After bioinformatics analysis, the new sequence was expressedinSaccharomycescerevisiaein codon optimized form. Its activity and substrate specificity was investigated through exogenous feeding different fatty acid substrates at different temperature. As a result, the new sequence-encoded protein named as oAiFADS17 could desaturate not only 18C PUFAs but also 20C PUFAs, with a strong preference for AA (arachidonic acid; 20∶4). The new desaturase was able to convert the ω-6 AA to the ω-3 EPA with a substrate conversion rate of 46.3% at 28 ℃. The enzymatic conversion rate for substrates with different carbon length at 28 ℃ was higher than that at 12 ℃. In this research, a new ω-3 desatutase oAiFADS17 with specific catalyzing function for 20C PUFAs, especially AA at 28 ℃, was successfully identified. The new enzyme provide theoretical basis for constructing EPA cell factory and its industrial production.
ω-3 desaturases; conversion rate; eicosapentaenoic acid (EPA)
10.13995/j.cnki.11-1802/ts.201608006
硕士研究生(陈海琴教授为通讯作者,E-mail:haiqinchen@jiangnan.edu.cn)。
国家自然科学基金(21276108)
2015-1-18,改回日期:2016-2-19
