不同干扰方式对桂西北灌草植被物种多样性和生产力的影响
2016-10-21韦兰英韦启忠莫祝平童德文
韦兰英, 韦启忠, 莫祝平, 童德文
(广西林业勘测设计院, 南宁 530011)
不同干扰方式对桂西北灌草植被物种多样性和生产力的影响
韦兰英, 韦启忠, 莫祝平, 童德文
(广西林业勘测设计院, 南宁 530011)
灌草植被在生态系统功能的维持中发挥着重要作用,不同的干扰方式可能改变退化土地灌草植被的多样性及其分布格局,甚至多样性与生产力的关系。以未受干扰的植被作为对照,研究放牧和火烧两种主要干扰对桂西北灌草植被的物种多样性和生产力的影响,并分析了两者之间的关系。结果表明:放牧和火烧处理均降低了灌木的物种丰富度和Shannon-Wiener多样性指数,但放牧处理的均匀度指数与对照相比无显著差异,火烧处理反而显著增加了其均匀度指数。对草本而言,放牧不影响物种丰富度指数,但显著降低了Shannon-Wiener多样性指数和均匀度指数;火烧显著降低了物种丰富度指数,但不影响Shannon-Wiener多样性指数和均匀度指数。与对照相比,放牧和火烧均降低了灌木的地上和地下部分生物量,但两者对草本的地上和地下生物量却产生了正面影响或无影响。此外,这两种干扰方式对灌木和草本多样性与生产力间关系的影响具有不一致性。
干扰; 多样性; 生产力; 灌草植被; 退化土地; 桂西北
当前,干扰活动作为不可忽视的因素正在强烈甚至不可逆转地改变着地表的植被组成及其多样性[1-2]。作为植物群落的两个重要特征,物种组成和多样性不仅影响生态系统的稳定性,还影响生态系统的结构和功能[2-3]。生产力作为重要的生态系统功能,与物种多样性关系密切[4-6],除物种多样性外,植物群落本身的性质(特征)、外界环境条件、干扰方式均可影响群落生产力[7-9],而且各因素间具有复杂的关系。然而,由于研究时间和研究尺度的不同,多样性与生态系统功能存在不同的关系[3-4,6-7,10-11],这可能也是目前许多试验研究得不到一个统一的物种丰富度—生产力关系模式的主要原因[12]。但大多数研究认为,在相同的气候条件和植被类型下,物种组成和多样性对生产力的作用主要取决于组成群落中各物种的功能特征(物种组成)及其关系(多样性)对生产力的贡献。然而,干扰条件下这种关系将会如何改变尚不清楚。
与森林相比,灌草植被在生物多样性、环境保护和地力维持等方面发挥着重要作用。放牧和火烧作为生态系统的两种主要干扰方式,可能会对生态系统过程产生深刻影响,如Heisler等发现放牧和火烧可能会改变植物群落的组成,结构和多样性格局,进而影响生态系统结构和功能[13-14]。同时研究发现,放牧对植物群落物种组成、多样性和生产力的影响与植物群落类型及其生产力紧密相关[15-16]。随着生产力的增加,放牧导致物种组成发生更多的变化[15],对不同的植物群落而言,放牧反而能增加群落的多样性[17],而火灾有利于增加已经建立的灌木群落的盖度及其丰富度[13]。这就表明干扰对植物群落的影响还与干扰特征、植物群落特征及植物的生物学特性和受干扰地点的资源条件有关。
由于受不合理的人类活动(采伐、放牧、火烧和开垦)的影响,桂西北大面积天然植被遭受破坏,原生植被几乎不再存在。植被破坏的后果不仅导致大规模的土地退化,还导致了生物多样性的丧失和生产力的下降。目前,灌草植被是该区域退化土地的主要植被类型。研究干扰条件下灌草植被的多样性和生产力特征,有助于了解灌草植被在维持生态系统功能中的重要作用。本研究以桂西北未受干扰的灌草植被作为对照,研究放牧和火烧两种主要干扰方式对灌草植被物种多样性、生产力及多样性与生产力关系的影响。开展这一研究,有助于了解放牧和火烧这两种主要干扰方式对退化土地灌草植被的多样性及其生产力的影响,有助于预测这种影响对该区域植物多样性与分布格局的可能效应。
1 研究区自然条件和研究方法
1.1研究区自然概况
研究区位于广西西北部的田林县高龙乡者车村(23°58′—25°37′N,104°47′—106°55′E ),该区域地处珠江上游,属云贵高原的东南边缘,位于南亚热带季风气候区,年均温20℃,年均降雨量1 500 mm。该区历史上分布有大面积森林,但由于人类活动的影响(如过度采伐、开垦、薪柴、放牧以及火烧等),区域植被退化十分严重,原生植被几乎不再存在,现有植被大多数为人工种植的马尾松林、杉木林和竹林。灌草植被是退化土地的主要植被类型,遭受放牧、火烧的样地均有20 a历史,主要的植物种类有野牡丹(MelastomacandidumD.Don)、柃木(E.japonicaThunb)、粗糠柴(Mallotusphilippensis)、铁芒箕(Dicranopterislinearis(Burm.)Underw.)、五节芒(Miscanthusfloridulus(Labill.)Warb.)、蔓生莠竹(Microstegiumvagans(Nees ex Steud.) A.Camus.)和纤毛鸭嘴草(I.indicum(Houtt.) Merr.)等,其样地基本特征如表1所示。

表1 研究样地概况
1.2研究方法
于2007年5—8月对研究区域内遭受不同类型干扰的灌草植被进行样方调查,放牧样地为长期放牧样地,火烧样地每年经受一次火烧,我们于火烧后半年进行采样。每个类型分别设置4个10 m×10 m的样地,每个样地内部再随机布置4个2 m×2 m的样方进行植被调查。各样方设置后,先用GPS确定样方的位置,并记录相关地理位置指标和立地条件(经度、纬度、海拔、坡向和坡位),然后调查样方内灌木和草本的种类和数量。植被调查后,采用全部收获法分别测定样方中出现的灌木和草本的地上和地下生物量。测定时,地下部分挖至无根系为止,然后将灌木和草本分开,分别测定各自地上和地下部分生物量的鲜重,并取出相应样品,带回实验室,85℃烘干,测定其干重。
1.3数据处理
采用物种丰富度指数(Ma)、Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)进行多样性的分析,计算公式如下[19]:
(1) 丰富度指数:
物种丰富度指数(R)=出现在样方中的物种数
(2) Shannon-Wiener指数(H′):
(3) Pielou均匀度指数(J):
式中:Pi为第i种的个体数ni占所有种的个体总数n的比例,即Pi=ni/n;S为物种数;i=1,2,3。
采用SPSS统计分析软件包(SPSS 16.0 for Windows,Chicago,USA)对数据进行相关分析、One-Way ANOVA方差分析,并用LSD法进行多重比较。
2 结果与分析
2.1不同干扰方式对灌木和草本物种多样性的影响
对照、放牧和火烧灌木的物种丰富度指数、Shannon-Wiener多样性指数和均匀度指数的变化范围为5~8,1.42~1.87,0.84~0.94。草本的物种丰富度指数、Shannon-Wiener多样性指数和均匀度指数的变化范围为4~5,0.85~1.10,0.50~0.69(图1)。与对照相比,放牧和火烧均显著降低了灌木的物种丰富度和Shannon-Wiener多样性指数(p<0.001),但放牧不影响其均匀度指数(p>0.05),火烧显著增加了均匀度指数(p<0.001)。对草本而言,放牧不影响物种丰富度指数(p>0.05),但显著降低了Shannon-Wiener多样性指数和均匀度指数(p<0.01);火烧显著降低了物种丰富度指数(p<0.05),不影响Shannon-Wiener多样性指数和均匀度指数(p>0.05)。可见,放牧和火烧对灌木和草本的物种丰富度指数(S)、Shannon-Wiener多样性指数(H)和均匀度指数(J)均产生了不同程度的影响,不同处理间差异显著。

注:不同大写字母代表不同干扰方式间灌木物种数差异显著,不同小写字母代表不同处理间草本物种数差异显著。
图1不同干扰方式对灌木和草本物种多样性的影响
2.2不同干扰方式对灌木和草本地上、地下生物量的影响
放牧和火烧均显著降低了灌木地上部分生物量(AGB)和地下部分生物量(BGB)(p<0.001),放牧和火烧的地上和地下生物量无显著差异(p>0.05)(图2)。放牧显著增加了草本的地上部分生物量(AGB),对其地下部分生物量(BGB)无影响(p>0.05);火烧显著增加了草本的地上和地下生物量(p<0.001)。
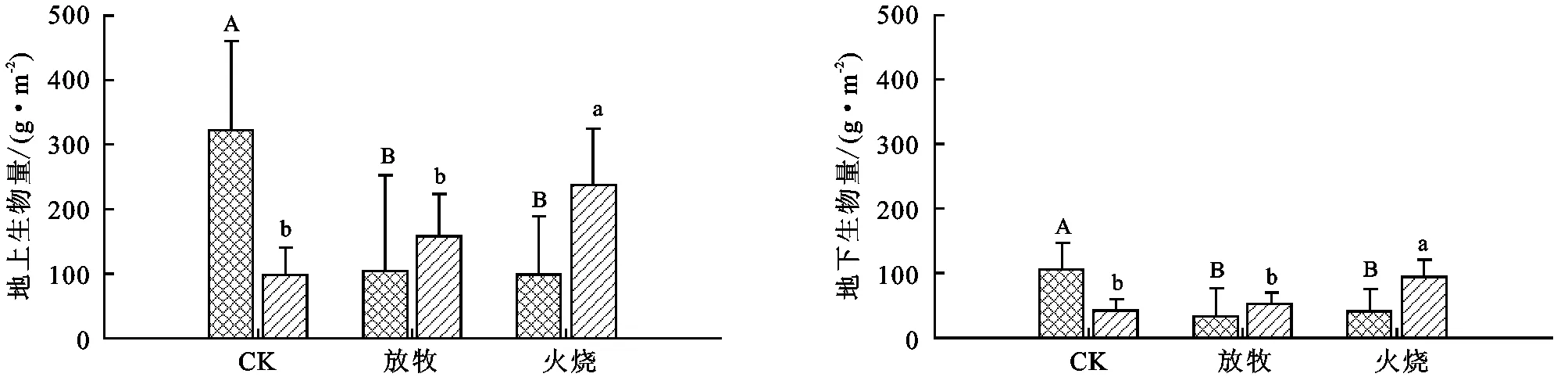
注:大写字母代表不同干扰方式间灌木物种数的差异,小写字母代表草本间的差异。
图2不同干扰方式对灌木和草本生物量的影响
2.3不同干扰方式对灌木和草本物种多样性和生物量关系的影响
2.3.1不同干扰方式对丰富度指数与生物量关系的影响物种丰富度与群落地上与地下生物量的关系在对照、放牧和火烧间存在差异(图3)。与对照相比,放牧和火烧处理没有改变灌木地上部分生物量与物种多样性间的正相关关系,但相关性减弱;放牧处理改变了草本地上部分生物量与物种多样性的关系,火烧处理两者关系与对照相同,为负相关,但相关性增强。与对照相比,火烧和放牧对物种丰富度与地下生物量间的影响与对地上部分生物量与物种多样性间的关系相似。

图3不同干扰方式对物种数与生物量关系的影响
2.3.2不同干扰方式对多样性指数与生物量的关系影响对照、放牧和火烧处理的灌木Shannon-Wiener多样性指数与地上和地下生物量均为正相关关系;草本shannon-Wiener多样性指数则与地上和地下生物量呈负相关关系(图4)。可见,与对照相比,放牧和火烧并没有改变地上、地下部分生物量与Shannon-Wiener多样性指数之间的关系,但两者相关性减弱(图4)。
2.3.3不同干扰方式对均匀度指数与生物量关系的影响对照和放牧灌木均匀度指数与地上和地下生物量均为正相关关系,但是火烧处理则为负相关关系;对照和放牧草本均匀度指数与地上和地下生物量为负相关关系,火烧处理则相反(图5)。可见,放牧没有改变灌木和草本均匀度指数与地上和地下生物量的关系,但是火烧处理则改变了这种关系。

图4不同干扰方式对多样性指数与生物量关系的影响
3 讨论与结论
放牧和火烧作为主要的干扰活动,是植被组成及其动态变化的主要驱动因素[20-21]。通常情况下,人们认为放牧和火烧一般会降低物种丰富度、多样性和生产力[14,22-23],如果能免于放牧,一般能增加物种丰富度[24]。事实上,放牧不仅能改变物种的时空分布格局[25],也可能增加物种丰富度和多样性[17,26],这取决于放牧的程度[27]。放牧主要是通过改变土壤条件进而改变植物间的竞争关系,从而改变群落组成和多样性[28]。由于火烧可能对种子库及其萌发和定居产生正面或负面影响,因此一般会改变植被的组成和结构[21,23],但对物种多样性的影响却表现为增加、降低或者无影响[13,29-31]。
本研究中,放牧和火烧均显著降低了灌木的物种丰富度和多样性指数,但对草本物种丰富度和多样性指数却产生了不同的影响,这表明外界干扰对植物群落的影响不仅因干扰方式而异,也因群落类型不同而异[21]。虽然避免放牧能增加物种数量,但是同时对物种均匀度也会产生影响,因此,合理的放牧和适宜的火烧,对维持生物多样性和关键的生态系统过程均具有非常重要的作用[27,31]。

图5不同干扰方式对均匀度指数与生物量关系的影响
刘玲玲等[27]对滇西北不同管理方式亚高山草地的物种多样性和生物量的研究发现,适量放牧有利于地上生物量的净增长,过度放牧将促使生物量向地下转移,但大多数情况下,放牧一般会增加向地下部分分配的生物量[27,31],这是由于放牧可能通过改变植物—土壤的反馈关系,进而改变植物的生物量分配[20]。然而,本研究中放牧反而减少了灌木的地下部分生物量,但对草本的地上和地下生物量却产生了正面影响或无影响,这可能是由于本研究中的放牧强度与前人研究不同,同时表明灌木和草本对放牧的响应程度不同。火烧对灌木和草本地上和地下生物量的影响与放牧对灌木和草本对地上与地下生物量的影响相同,这可能是由于火烧清除灌木后对草本反而产生有利了影响,与Nayaka等[31]研究一致。
漆良华等[32]对不同植物群落的物种多样性与生物量的研究发现,可用“S”曲线较好地描述退化土地物种丰富度与群落生物量之间的关系,双曲线能较好地描述群落多样性指数、群落均匀度指数与生物量之间的关系。本研究中,物种丰富度与生物量的关系表现为正相关、负相关和无相关三种关系。目前,关于干扰对植物群落多样性与生产力间关系的影响至今还没有一致的结论,二者的关系主要有单增、单峰、正线性相关、对数线性增加以及无显著相关等类型[4,5,12,33],其中无相关性出现的最多[34]。多样性与生产力间以多种方式相关,可能是由于各种机制间的相互作用以及环境复杂性和时空尺度分析的差异性导致[8],也可能是由于研究区域、研究尺度和多样性指标选择差异等因素导致。本研究中,与对照相比,放牧和火烧并未改变灌木物种多样性与生产力之间的关系,但相关性减弱;放牧处理改变了草本物种多样性与生产力之间的关系。放牧和火烧对物种多样性与生产力关系的影响存在不确定性,这可能与干扰方式有关,但也可能是由于退化土地灌草植被物种多样性和生产力之间不存在必然的因果联系,这与袁自强等[34]的研究一致。因为在自然生态系统中,物种的分布、数量、特征先决于环境条件,研究位点间环境因子的差异往往影响群落结构和生产力的关系。
我们研究发现均匀度指数与生产力的关系(地上和地下生物量)要高于丰富度指数与生产力的关系,这与Ebrahimi 等[24]的研究一致,可能是由于均匀度指数对生产力的贡献更大导致。一般说来,物种丰富度高的立地,生产力也较高,丰富度指数与生产力关系较弱,主要是由于研究区域的灌草植被为稀疏植被[35]。尽管由于复杂的地形、气候和传统的土地利用实践活动,山区具有较多的生物多样性,然而由于放牧和火烧,却对灌木生物多样性产生了负面的影响,但对草本多样性和生物量却无影响或产生了正面影响。可见,放牧和火烧对桂西北退化土地灌木和草本多样性和生产力的影响还存在不确定性。因此,未来研究中,我们应该考虑生物因素和非生物因素以及与之相连的环境因素和人类干扰活动(如放牧和火烧等),因为这些因素可能会通过改变生态系统条件进而改变植被组成或导致入侵植物的出现,进而导致生态系统结构和功能的改变[1,17,36],这种改变对于退化生态系统尤为关键。未来研究我们应该综合这方面考虑,以期为预测生态系统的动态变化提供科学依据。
[1]Keeley J E, Lubin D, Fotheringham C J. Fire and grazing impacts on plant diversity and alien plant invasions in the southern Sierra Nevada[J]. Ecological Applications,2003,13(5):1355-1374.
[2]Hooper D U, Chapin F S, Ewel J J, et al. Effects of biodiversity on ecosystem functioning:a consensus of current knowledge[J]. Ecological Monographs,2005,75(1):3-35.
[3]Bond E M, Chase J M. Biodiversity and ecosystem functioning at local and regional spatial scales[J]. Ecology Letters,2002,5(4):467-470.
[4]Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems[J]. Nature,1996,379(6567):718-720.
[5]Tilman D. Distinguishing between the effects of species diversity and species composition[J]. Oikos,1997,80(1):185.
[6]Tilman D, Knops J, Wedin D, et al. The influence of functional diversity and composition on ecosystem processes[J]. Science,1997,277(5330):1300-1302.
[7]Grime J P. Benefits of plant diversity to ecosystems:immediate, filter and founder effects[J]. Journal of Ecology,1998,86(6):902-910.
[8]Spehn E M, Hector A, Joshi J, et al. Ecosystem effects of biodiversity manipulations in European grasslands[J]. Ecological Monographs,2005,75(1):37-63.
[10]高本强,袁自强,王斌先等.施肥和刈割对亚高山草甸物种多样性与生产力及其关系的影响[J].植物生态学报,2014,38(5):417-424.
[11]张全国,张大勇.生物多样性与生态系统功能:最新的进展与动向[J].生物多样性,2003,11(5):351-361.
[12]杜国祯,覃光莲,李自珍等.高寒草甸植物群落中物种丰富度与生产力的关系研究[J].植物生态学报,2003,27(1):125-132.
[13]Heisler J L, Briggs J, Knapp A. Long-term patterns of shrub expansion in a C4-dominated grassland:fire frequency and the dynamics of shrub cover and abundance[J]. American Journal of Botany,2003,90(3):423-428.
[14]Bai Y F, Wu J G, Pan Q M et al. Positive linear relationship between productivity and diversity:evidence from the Eurasian Steppe[J]. Journal of Applied Ecology,2007,44(5):1023-1034.
[15]Oesterheld M, Semmartin M. Impact of grazing on species composition: Adding complexity to a generalized model[J]. Austral Ecology,2011,36(8):881-890.
[16]Lezama F, Baeza S, Altesor A, et al. Variation of grazing-induced vegetation changes across a large-scale productivity gradient[J]. Journal of Vegetation Science,2014,25(1):8-21.
[17]Chaneton E J, Perelman S B, Omacini M, et al. Grazing, environmental heterogeneity, and alien plant invasions in temperate Pampa grasslands[J]. Biological Invasions,2002,4(1/2):7-24.
[18]毛志宏,朱教君.干扰对植物群落物种组成及多样性的影响[J].生态学报,2006,26(8):2695-2701.
[19]马克平.生物群落多样性的测度方法.Ⅰ:α多样性的测度方法[J].生物多样性,1994,2(3):162-168.
[20]Veen G F, de Vries S, Bakker ES, et al Grazing-induced changes in plant-soil feedback alter plant biomass allocation[J]. Oikos,2014,123(7):800-806.
[21]Santana V M, Alday J G, Baeza M J. Effects of fire regime shift in Mediterranean Basin ecosystems:changes in soil seed bank composition among functional types[J]. Plant Ecology,2014,215(5):555-566.
[22]Schultz N L, Morgan J W, Lunt I D. Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient[J]. Journal of Vegetation Science,2011,22(1):130-142.
[23]Salazar A, Goldstein G. Effects of Fire on Seedling Diversity and Plant Reproduction (Sexual vs. Vegetative) in Neotropical Savannas Differing in Tree Density[J]. Biotropica,2014,46(2):139-147.
[24]Ebrahimi S S, Pourbabaei H, Potheir D, et al. Effect of livestock grazing and human uses on herbaceous species diversity in oriental beech (FagusorientalisLipsky) forests, Guilan, Masal, northern Iran[J]. Journal of Forestry Research,2014,25(2):455-462.
[25]Aikens M L, Ellum D, Ellum D, et al. The effects of disturbance intensity on temporal and spatial patterns of herb colonization in a southern New England mixed-oak forest[J]. Forest Ecology and Management,2007,252(1/3):144-158.
[26]Takala T, Tahvanainen T, Kouki J. Grazing promotes bryophyte species richness in semi-natural grasslands[J]. Ann. Bot. Fennici,2014,51(3):148-160.
[27]刘玲玲,吴兆录,李青,等.滇西北藏族地区不同管理方式的草地生物量的比较研究[J].生态学杂志,2005,24(12):1409-1412.
[28]Medina-Roldán E, Paz-Ferreiro J, Bardgett R D. Grazing-induced effects on soil properties modify plant competitive interactions in semi-natural mountain grasslands[J]. Oecologia,2012,170(1):159-169.
[29]Silva da D M, Batalha M A. Soil-vegetation relationships in Cerrados under differ-ent fire frequencies[J]. Plant Soil,2008,311(1/2):87-96.
[30]Stephan K, Miller M, Dickinson M B. First-order fire effects on herbsand shrubs:present knowledge and process modeling needs[J]. Fire Ecology,2010,6(1):95-114.
[31]Nayak R R, Vaidyanathan S, Krishnaswamy J. Fire and grazing modify grass community response to environmental determinants in savannas:Implications for sustainable use[J]. Agriculture, Ecosystems and Environment,2014,185:197-207.
[32]漆良华,彭镇华,张旭东,等.退化土地植被恢复群落物种多样性与生物量分配格局[J].生态学杂志,2007,26(11):1697-1702.
[33]王长庭,龙瑞军,王启基,等.高寒草甸不同草地群落物种多样性与生产力关系研究[J].生态学杂志,2005,24(5):483-487.
[34]袁自强,魏盼盼,高本强,等.取样尺度对亚高寒草甸物种多样性与生产力关系的影响[J].植物生态学报,2012,36(12):1248-1255.
[35]Komac B, Domènech M, Fanlo R. Effects of grazing on plant species diversity and pasture quality insubalpine grasslands in the eastern Pyrenees(Andorra):Implications for conservation[J]. Journal for Nature Conservation,2014,22(3):247-255.
[36]Cingolani A M, Noy-Meir I, Díaz S. Grazing effects on rangeland diversity: A synthesis of contemporary models[J]. Ecological Application,2005,15(2):757-775.
Effects of Different Disturbances on Species Diversity and Productivity of the Shrub-Grass Vegetation on the Degraded Land in Northwest Guangxi, China
WEI Lanying, WEI Qizhong, MO Zhuping, TONG Dewen
(GuangxiForestInventoryandPlanningInstitute,Nanning530011,China)
Shrub-grass vegetation plays an important role in maintaining the functions of ecosystems. On degraded lands, different disturbances might alter species diversity and distribution pattern of shrub-grass vegetation, and even the relationship between plant species diversity and productivity. However, our knowledge of the consequences of these effects on diversity and composition of ground flora remains very poor. In this study, we examined the species diversity, productivity and the relationship between diversity and productivity in undisturbed, grazing and fire vegetation of northwest Guangxi. It′s found that shrubs exhibited lower species richness and Shannon-Wiener index under both grazing and fire treatments than those of the undisturbed site. Moreover, compared with undisturbed treatment, the evenness index of shrubs differed insignificantly under grazing treatment, while it′s reduced by fire treatment. With respect to grass species, grazing had no influence on their diversity, but it did reduce the Shannon-Wiener index and evenness index distinctly. On the contrary, fire treatment decreased the grass species diversity, whereas it showed little effect on the Shannon-Wiener index and evenness index. In addition, the relationship between species diversity and productivity of shrubs and grasses was also affected inconsistently by grazing and fire treatment.
disturbance; species diversity; productivity; shrub-grass vegetation; degraded land; Northwest Guangxi
2015-10-16
2015-11-20
广西自然科学基金项目(桂财教[2015]139号;广西“新世纪十百千人才工程”专项基金(桂人社办〔2012〕第92号);广西林业科技项目(桂林科字〔2014〕第21号)
韦兰英(1980—),女,广西桂林人,博士,助理研究员,研究方向为植物生态学。E-mail:weilanyingccn@163.com
韦启忠(1964—),男,广西上林人,学士,高级工程师,研究方向为林业调查规划。E-mail:wqz1978@sina.com
Q948
A
1005-3409(2016)04-0288-06
