冰温贮藏对羊肉中蛋白质磷酸化水平的影响
2016-10-14刘永峰张德权
张 艳,李 欣,李 铮,李 蒙,刘永峰,张德权
冰温贮藏对羊肉中蛋白质磷酸化水平的影响
张 艳1,2,李 欣2,李 铮2,李 蒙2,刘永峰1,张德权2
(1陕西师范大学食品工程与营养科学学院,西安 710119;2中国农业科学院农产品加工研究所/农业部农产品加工重点实验室,北京 100193)
【目的】通过测定冰温贮藏及冷藏过程中肌肉肌浆蛋白与肌原纤维蛋白的磷酸化水平,分析冰温贮藏对蛋白质磷酸化水平的影响,为肉类冰温贮藏品质调控提供理论依据。【方法】选取大尾寒羊与小尾寒羊杂交公羊的背最长肌于冷藏和冰温条件下成熟,分别在0.5 h、12 h、24 h、3 d、5 d、9 d取样测定蛋白激酶活性、肌浆蛋白与肌原纤维蛋白的磷酸化水平。采用蛋白激酶活性测定试剂盒、运用酶联免疫的方法检测各处理组蛋白激酶的活性随贮藏时间的变化;通过SDS-PAGE电泳、荧光染色的方法得到全蛋白染色与磷酸化染色的肌浆蛋白与肌原纤维蛋白条带,运用Quantity One软件分析各处理组各处理时间的肌浆蛋白及肌原纤维蛋白的磷酸化水平。【结果】贮藏初期(0.5—12 h),冰温组蛋白激酶活性显著升高(<0.05),冷藏组蛋白激酶活性则显著降低(<0.05),贮藏12 h—9 d过程中,冰温组蛋白激酶活性均显著高于冷藏组(<0.05)。两处理组蛋白激酶活性均随贮藏时间的延长而逐渐降低,且冰温组蛋白激酶活性均显著高于冷藏组(<0.05)。对肌浆蛋白染色效果图中分布于15—250 kDa的17个蛋白条带逐一进行分析,结果表明贮藏温度极显著影响肌浆蛋白各蛋白条带的磷酸化水平(<0.001)。冷藏组肌浆蛋白整体磷酸化水平先升高后降低,贮藏第3天达到最大值,冰温组肌浆蛋白整体磷酸化水平先降低后升高,最小值出现在贮藏第24天,贮藏第12天至3天时,冷藏组肌浆蛋白整体磷酸化水平高于冰温组(<0.05),贮藏第9天时,冰温组肌浆蛋白整体磷酸化水平高于冷藏组(<0.05)。对肌原纤维蛋白染色效果图中分布于15—250 kDa的20个蛋白条带逐一进行分析,结果表明不同贮藏温度对所有肌原纤维蛋白条带的磷酸化水平均产生极显著影响作用(<0.001)。两处理组肌原纤维蛋白整体磷酸化水平均随贮藏时间的延长呈现先升高后降低的趋势,冷藏组与冰温组最大值分别出现在24 h和12 h,贮藏初期(0.5—12 h),两处理组间肌原纤维蛋白整体磷酸化水平无显著差异(<0.05),贮藏24 h至贮藏期结束,冷藏组肌原纤维蛋白整体磷酸化水平始终高于冰温组(<0.05)。【结论】冰温贮藏通过影响蛋白激酶的活性来调控蛋白质的磷酸化水平,进而通过影响糖酵解途径、肌肉收缩及骨架蛋白降解来间接调控肉品质。
冰温;蛋白激酶;肌浆蛋白;肌原纤维蛋白;蛋白质磷酸化
0 引言
【研究意义】冰温贮藏是指将食品贮藏在其冰点以上、0℃以下的温度区域,该温度区域既可使机体的生理活动降到最低程度,又能维持其正常的新陈代谢,既能抑制微生物生长,又不会对食品产生冻害,进而起到长期保鲜食品的作用[1]。磷酸化是经蛋白激酶催化的最常见、最重要的一种蛋白质的翻译后修饰方式[2]。糖酵解、肌肉收缩及蛋白质降解等与肉品质形成相关的生物学过程都受蛋白质磷酸化和去磷酸化这一可逆过程的调节[3-5]。因此,研究冰温贮藏条件下肌肉蛋白质磷酸化水平的变化,可为研究宰后蛋白质磷酸化修饰调控肉品质的形成和劣变提供理论依据。【前人研究进展】李培迪等[6]的研究表明,相较于冷藏,冰温贮藏可使丙酮酸激酶活性升高,乳酸脱氢酶活性降低,同时延长μ-钙蛋白酶的活性保持时间,从而延缓肌肉糖酵解进程,延迟肌肉成熟,说明贮藏温度能够通过调控酶的活性影响糖酵解过程。ANDERSON等[7]指出宰后肌浆蛋白中葡萄糖磷酸变位酶-1的磷酸化水平与肌肉剪切力呈现负相关,另外,SHEN等[8]的研究指出,猪宰后早期背最长肌中蛋白激酶的激活导致磷酸果糖激酶2(PFK-2)和磷酸果糖激酶1(PFK-1)的磷酸化,进而形成白肌肉(PSE肉),这表明宰后肌肉品质与肌浆蛋白的磷酸化关系密切。同时,HUANG等[9]研究发现宰后肌肉成熟及肌肉品质变化与肌原纤维蛋白磷酸化水平相关。HEELEY[10]、MAZZEI[11]、RYDER[12]等研究发现肌肉中大部分肌原纤维蛋白的功能特性均受蛋白质磷酸化和去磷酸化过程影响,可见肉品质变化同样与肌原纤维蛋白的磷酸化存在紧密联系。【本研究切入点】冰温贮藏有利于延缓宰后肌肉的成熟进程,从而调控肌肉品质,同时有研究表明宰后成熟过程中(0—24 h)肌浆蛋白的磷酸化修饰水平能够间接影响糖酵解进程,肌原纤维蛋白的磷酸化修饰可以通过影响肌肉收缩而调控肉的僵直及嫩化进程。肌浆蛋白与肌原纤维蛋白是肌肉蛋白最主要的组成成分,肌浆蛋白包括大多数与糖酵解有关的酶类,肌原纤维蛋白包含主要的收缩蛋白及收缩调控蛋白[13]。在宰后较长时间的贮藏过程中蛋白质磷酸化水平是否会发生变化,贮藏条件是否会对蛋白质磷酸化水平产生影响,均未见报道。【拟解决的关键问题】通过检测贮藏过程中蛋白激酶的活性、肌浆蛋白及肌原纤维蛋白的磷酸化水平,揭示冰温贮藏对蛋白质磷酸化水平的影响。
1 材料与方法
试验于2015年10月至2016年2月在中国农业科学院农产品加工研究所进行。
1.1 试验材料
选取饲养方式相同、体重相近的8月龄大尾寒羊×小尾寒羊杂交公羊5只。采用清真方式屠宰,宰前静养1 h以消除应激,宰后0.5 h内迅速取下两侧背最长肌。
每只羊在宰后0.5 h取其任意一侧背最长肌靠近尾端的1/7,液氮速冻,贮存于-80℃,用于测定初始指标。剩余背最长肌样品均在宰后2 h内于冰盒上运回实验室,每只羊两侧背最长肌随机分为冷藏及冰温贮藏处理组,每条背最长肌样品平均分为6份,从头至尾随机分配至各个贮藏时间(12 h、24 h、3 d、5 d、9 d,剩余一块用于监测温度),白色托盘盛放,贴上标签,食品级PE保鲜膜,氧气透过率为(10 600± 20)% cm3/(m2·24 h·atm)包裹后放入-18℃冰箱进行降温,冷藏组待肉块中心温度降至4℃时取出,冷藏贮存(2—4℃),冰温组待肉块中心温度降至-1℃时取出,冰温贮存(-1—-1.5℃)。宰后12 h、24 h、3 d、5 d、9 d取出相应肉块,去除可见脂肪及筋膜后经液氮速冻,保存于-80℃。
1.2 试验试剂与仪器
1.2.1 主要试剂 试验使用试剂均为分析纯。三羟甲基氨基甲烷(Trizma base)、二硫苏糖醇(DTT)、甲叉双丙烯酰胺(methylene diacrylamide)、丙烯酰胺(Acrylamide)、过硫酸铵(APS)、四甲基乙二胺(TEMED)、十二烷基硫酸钠(SDS)、甘氨酸、曲拉通-100(Triton-100)、EGTA、巯基乙醇(β-mercaptoethanol)、考马斯亮蓝G250、溴酚蓝、甘油,美国Sigma公司;蛋白酶抑制剂(Roche Complete)、磷酸酶抑制剂(Roche PhosStop),瑞士Roche公司;蛋白浓度测定试剂盒(BCA protein assay kit),美国Pierce公司;蛋白激酶活性测定试剂盒,加拿大Immunechem公司;Pro-Q染色液(Pro-Q Diamond)、Ruby染色液(SYPRO Ruby),美国Invitrogen公司;氯化钾(KCl)、无水乙酸钠、磷酸、乙腈、甲醇、无水乙醇、乙酸,国药集团化学试剂北京有限公司。
1.2.2 主要仪器 MIR-154-PC低温恒温培养箱,日本Panasonic公司;Ultra Turrax Disperser S25匀浆器,德国IKA公司;CR22GII高速冷冻离心机,日本日立公司;SpectraMax 190多功能酶标仪,美国Molecular Devices公司;1-14小型台式离心机,美国Sigma公司;DK-S28电热恒温水浴锅,上海精宏实验设备有限公司;电泳仪、凝胶成像仪,美国Bio-Rad公司;T6紫外可见分光光度计,北京普析通用仪器有限责任公司;超低温冰箱,美国Thermo公司。
1.3 试验方法
采用Pro-Q Diamond和SYPRO Ruby染液分别检测磷酸化蛋白和全蛋白。将某一蛋白条带经Pro-Q Diamond染色后的灰度值(P)与SYPRO Ruby染色后的灰度值(T)之比作为该条带磷酸化水平。试验共60个样品(5只羊×2个处理组×6个时间点),所有样品从蛋白质提取到进行电泳及蛋白质条带染色均进行3次以上重复性检测。
1.3.1 蛋白激酶活性测定 取-80℃冻存的肉样,用预冷的生理盐水洗涤2次,滤纸吸干,称取3 g肉样,加入12 mL预冷缓冲液(25 mmol·L-1Tris-HCl,2 mmol·L-1EGTA,50 mmol·L-1巯基乙醇),冰浴中15 000×充分匀浆(3次,每次15 s,间隔30 s)。在4℃、1 000×条件下离心30 min,上清液用两层纱布过滤,即得蛋白激酶粗提液,分装后于-80℃保存。
将蛋白激酶粗提液稀释20倍后采用考马斯亮蓝法测定样品蛋白浓度,调整原激酶粗提液至相同浓度,稀释2 000倍,采用酶联免疫方法,运用蛋白激酶活性测定试剂盒测定蛋白激酶活性,蛋白激酶相对活性以495 nm处样品吸光度值与空白(20×蛋白激酶提取缓冲液)吸光度值之差表示。
1.3.2 肌浆蛋白提取及其磷酸化水平测定 肌浆蛋白提取参考Lametsch等[14]的方法,并略作修改,取-80℃冻存的肉样1.0 g,加6 mL预冷的缓冲液(10 mmol·L-1Tris,10 mmol·L-1DTT,蛋白酶抑制剂1片/50 mL,磷酸酶抑制剂5片/50 mL),冰浴中15 000×匀浆30 s(3次,每次10 s,间隔30 s),4℃、20 000×离心30 min,上清液用两层纱布过滤即得肌浆蛋白,震荡均匀后分装,于-80℃贮存。
取-80℃贮存的肌浆蛋白,稀释8倍后用BCA法测定蛋白浓度,将肌浆蛋白浓度调整为2 μg·μL-1后与上样缓冲液(10% SDS 4 mL,甘油2 mL,2 mL 0.5 mol·L-1Tris-HCl pH6.8,2 mL 1 mol·L-1DTT,溴酚蓝0.01 g)1﹕1混合,沸水浴煮5 min,2 000×离心2 min,分装后-20℃贮存备用。
采用SDS-PAGE电泳、磷酸化染色及全蛋白染色测定肌浆蛋白磷酸化水平。分离胶浓度为14%,浓缩胶浓度为4%,上样量为5 μg。电泳结束后至成像前所有操作均需在摇床(60 r/min)上进行。固定液(50%甲醇,10%乙酸)固定(100 mL、30 min/次,2次)以确保凝胶上电泳缓冲液被清洗干净,超纯水水洗(100 mL、10 min/次,3次),Pro-Q Diamond染液避光染色80 min,Pro-Q脱色液(20%乙腈,50 mmol·L-1乙酸钠,pH 4.0)脱色(100 mL、30 min/次,2次),超纯水水洗(100 mL、5 min/次、3次),进行磷酸化蛋白染色成像;SYPRO Ruby染色液染色4 h,Ruby脱色液(10%乙醇,7%乙酸)脱色(100 mL、30 min/次,2次),超纯水水洗(100 mL、5 min/次、3次)。
使用ChemiDocTMMP凝胶成像仪对染色后的蛋白质凝胶进行扫描。Pro-Q Diamond染色后在激发波长为532 nm、发射波长为 580 nm、分辨率为 200 mm的条件下扫描。 SYPRO Ruby染色后在激发波长为532 nm、发射波长为610 nm、分辨率为200 mm的条件下扫描。
1.3.3 肌原纤维蛋白提取及其磷酸化水平测定 肌原纤维蛋白提取参考Smuder等[15]的方法并略作修改,取-80℃冻存的肉1.0 g,加6 mL预冷的缓冲液(10 mmol·L-1Tris,10 mmol·L-1DTT,蛋白酶抑制剂1片/50 mL,磷酸酶抑制剂5片/50 mL),冰浴中15 000×匀浆30 s(3次,每次10 s,间隔30 s),4℃、20 000×离心30 min,弃上清。沉淀中加入8 mL预冷的肌原纤维溶解缓冲液(100 mmol·L-1KCl,1% Triton),冰浴中匀浆15 s,4℃,2 000×离心15 min,弃上清,此操作重复2次。沉淀中加入10 mL 5% SDS(60℃),使肌原纤维蛋白充分溶解,80℃加热20 min,分装后-80℃保存。
取-80℃贮存的肌浆蛋白,稀释15倍后用BCA法测定蛋白浓度,将肌浆蛋白浓度调整为2 μg·μL-1后与上样缓冲液1﹕1混合,沸水浴煮沸5 min,2 000×离心2 min,分装后-20℃贮存备用。
肌原纤维蛋白SDS-PAGE电泳分离胶浓度为12%,浓缩胶浓度为4%,其磷酸化蛋白染色、全蛋白染色过程均同肌浆蛋白。
1.4 图像分析及统计分析
运用Quantity One(美国Bio-Rad公司)软件对经荧光染液染色的SDS-PAGE凝胶电泳蛋白条带的光密度值进行分析,得到每个条带的磷酸化蛋白染色(Pro-Q Diamond)光密度值(P)和全蛋白染色(SYPRO Ruby)光密度值(T),同一条带的磷酸化蛋白与全蛋白光密度值之比P/T即为该条带蛋白质磷酸化水平,样品整体磷酸化水平为其泳道上所有蛋白条带的P/T值之和。利用SPSS Statistics 20软件对试验结果进行分析,不同处理组间的差异显著性采用独立样本T检验进行分析;以贮藏条件、宰后时间及贮藏条件×宰后时间3个因素作为变量,动物作为随机因素,进行混合模型方差分析。多重比较方法采用Duncan法,显著性水平为<0.05。结果均以平均值±标准差表示。
2 结果
2.1 蛋白激酶活性分析
如图1所示,冰温贮藏条件下蛋白激酶活性在宰后0.5—12 h显著升高(<0.05),随后逐渐下降。冷藏过程中蛋白激酶活性随贮藏时间的延长而逐渐显著降低(<0.05),且冷藏过程中降低速率高于冰温贮藏,12 h—9 d的贮藏过程中冰温组蛋白激酶活性均显著高于冷藏组(<0.05)。在贮藏后期,冷藏组蛋白激酶接近失活状态而冰温组仍处于较高水平。
2.2 肌浆蛋白各蛋白条带磷酸化水平分析
冰温贮藏组及冷藏组在宰后0.5 h、12 h、24 h、3 d、5 d、9 d的肌浆蛋白SDS-PAGE电泳图如图2-a、2-b所示。对图2中分布于15—250 kDa的17个蛋白条带逐一进行分析,结果如表1所示。不同贮藏温度下,各个蛋白条带的磷酸化水平均差异显著(<0.001),条带2、3、5、6、10、14、15的磷酸化在不同贮藏温度和不同贮藏时间下均产生显著差异(<0.05),条带2、5、6、14、15、17的磷酸化水平在贮藏温度与贮藏时间的交互作用下差异显著(<0.05)。其中,条带4、7、9的磷酸化水平随贮藏时间的延长先升高后降低,且分别在第24 h、第5天、第12 h达到最大值。条带2、5、6、11、15的磷酸化水平在贮藏期内先降低后升高,最小值分别出现在第12 h(及24 h)、第24 h、第5天、第24 h、第12 h。条带10磷酸化水平随贮藏时间延长逐渐升高。条带8、12、13、17在整个贮藏期内磷酸化水平均处于较稳定状态。另外,冷藏组条带11、17的磷酸化水平显著高于冰温组(<0.05),而冰温组条带6、13、14、15、16的磷酸化水平显著高于冷藏组(<0.05)。
2.3 肌浆蛋白整体磷酸化水平随贮藏时间的变化
对图2中电泳图谱进行灰度值分析,不同贮藏处理组的整体磷酸化水平如图3所示。从图中可知,宰后冷藏组肌浆蛋白磷酸化水平呈现先上升后下降的趋势,冰温组在宰后0.5—24 h时磷酸化水平降低,随后上升。冷藏组肌浆蛋白最高磷酸化水平出现在贮藏第3天,而冰温组肌浆蛋白磷酸化水平最高值出现在宰后0.5 h及第5天。其中在贮藏第12、24 h及第3天时,冷藏组肌浆蛋白整体磷酸化水平均显著高于冰温组(<0.05),贮藏第9天时冰温组肌浆蛋白磷酸化水平显著高于冷藏组(<0.05)。

不同小写字母表示冷藏条件不同贮藏时间之间存在差异显著(P<0.05);不同大写字母表示冰温条件不同贮藏时间之间存在差异显著(P<0.05);*表示不同温度处理间差异显著(P<0.05)。下同

a:磷酸化肌浆蛋白;b:全肌浆蛋白 a: Image of phosphoproteins stained with Pro-Q Diamond; b:Image of total proteins stained with SYPRO Ruby

表1 不同贮藏条件对肌浆蛋白磷酸化水平的影响
表中各条带的磷酸化水平(P/T)均以平均值±标准差表示。不同小写字母表示同一条带不同处理间差异显著(<0.05)。下同
Phosphorylation level of each band is expressed as the mean±standard error. Different small letters represent significant difference (<0.05). The same as below

图3 冷藏、冰温贮藏过程中肌浆蛋白整体磷酸化水平分析
2.4 肌原纤维蛋白各蛋白条带磷酸化水平分析
冰温贮藏组及冷藏组在宰后0.5 h、12 h、24 h、3 d、5 d、9 d的肌原纤维蛋白SDS-PAGE电泳图如图4-a、4-b所示,其中图4-a为Pro-Q Diamond 染色的磷酸化蛋白,图4-b为SYPRO染色的全肌原纤维蛋白。对图4中20个蛋白条带逐个进行分析,结果如表2所示。不同贮藏温度对所有肌原纤维蛋白条带的磷酸化水平均产生极显著影响(<0.001),不同贮藏时间对所有的肌原纤维蛋白条带的磷酸化水平产生显著影响作用(<0.05),贮藏温度与贮藏时间的交互作用对条带2、3、4、5、7、8、9、10、13、14、15、17、18、20的磷酸化水平有显著影响作用(<0.05)。其中,条带10的磷酸化水平随贮藏时间的延长逐渐减低。条带8的磷酸化水平在贮藏期内先降低后升高,最小值出现在第12 h。条带9、11、16、20的磷酸化水平随贮藏时间的延长先升高后降低,最大值分别出现在第24 h、第12 h、第5天、第24 h。条带1、3、4、14的磷酸化水平均在贮藏第12 h达到最大值,条带15、17的磷酸化水平在贮藏第5天达到最大值。另外,冷藏组条带2、7、8、9、12、14、15、18的磷酸化水平显著高于(<0.05)冰温组,而条带1、4、10、11、16、19在冰温组的磷酸化水平显著高于(<0.05)冷藏组,说明在不同贮藏条件下,肌原纤维蛋白各蛋白条带的磷酸化水平变化并不一致。

a:磷酸化肌原纤维蛋白;b:全肌原纤维蛋白 a: Image of phosphoproteins stained with Pro-Q Diamond; b: Image of total proteins stained with SYPRO Ruby

表2 不同贮藏条件对肌原纤维蛋白磷酸化水平的影响
2.5 肌原纤维蛋白整体磷酸化水平随贮藏时间的变化
对图4中电泳图谱进行灰度值分析,不同贮藏处理组的整体磷酸化水平如图5所示,宰后羊肉肌原纤维蛋白整体磷酸化水平在冰温贮藏组与冷藏组中均呈现先上升后下降的趋势,其中冷藏组在宰后24 h时达到最大磷酸化水平,冰温贮藏组在宰后12 h达到最大磷酸化水平,且从贮藏24 h开始至贮藏期结束,冷藏组肌原纤维蛋白整体磷酸化水平均显著高于冰温组(<0.05)。
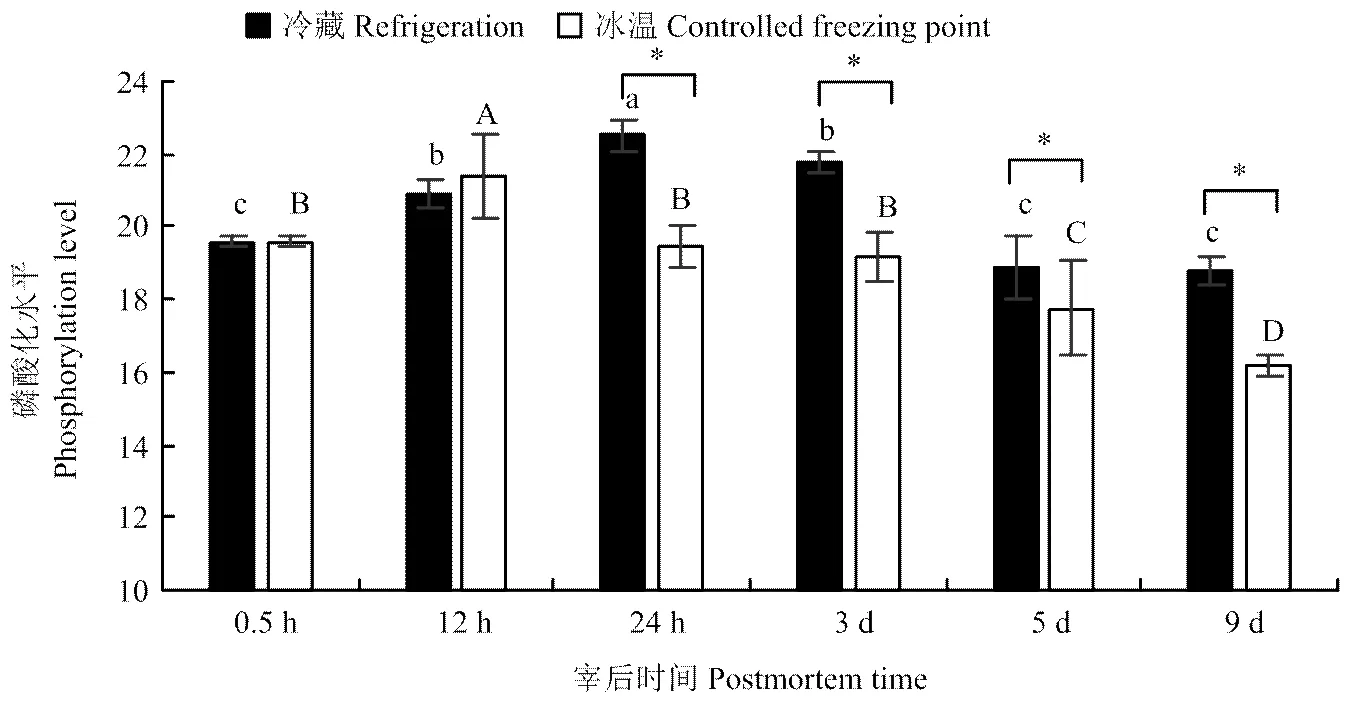
图5 冷藏、冰温贮藏过程中肌原纤维蛋白整体磷酸化水平分析
3 讨论
3.1 不同贮藏处理对蛋白激酶活性的影响
蛋白激酶是一类能够催化蛋白质磷酸化反应的酶,它们几乎在每一个细胞功能中都起着重要作用[16]。所有的蛋白激酶都处于基态,在需要时被不同的刺激因素激活,进而催化某些蛋白质的磷酸化反应并激活磷酸酶[17]。SHEN等[8]的研究表明,PSE肉中AMP激活性蛋白激酶(AMPK)在宰后早期被激活并在0.5 h酶活性最高,随后逐渐降低,而正常猪肉中AMPK酶活性在宰后1 h最高,说明AMPK活性对宰后肉品质存在影响。AMPK在宰后糖酵解进程及PSE肉的形成中起关键,宰后早期ATP的消耗与AMP的生成使AMPK被激活,导致肌肉pH快速下降,在猪肉中则表现为PSE肉的形成[18-20]。李培迪等[6]研究发现,冰温贮藏的肌肉较冷藏晚4 d达到其极限pH。本研究中,冰温条件下蛋白激酶活性的激活滞后于冷藏,结合李培迪的研究,可以从新的角度揭示冰温贮藏条件下肌肉达到极限pH所需时间较长的原因。
3.2 不同贮藏处理对肌浆蛋白磷酸化水平的影响
糖酵解作为宰后肌肉最主要的供能方式,与肌肉肉色、pH、系水力及肌肉僵直都有直接关系,肌浆蛋白中糖酵解酶占三分之二以上[21]。宰后肌肉中ATP大量消耗导致糖酵解速率加快,pH快速下降,这将会加速僵直现象的出现,促进成熟,减少僵直总时间[22-23]。另有研究表明,蛋白质磷酸化在调节糖酵解酶活性方面发挥着重要作用[3],因此,选择宰后羊肉的肌浆蛋白来检测并分析其磷酸化水平在不同贮藏条件下6个贮藏时间点的变化。
贮藏12 h至3 d时,冷藏组肌浆蛋白整体磷酸化水平高于冰温组,而第9天时冰温组高于冷藏组(图3),原因可能是贮藏初期冷藏组蛋白激酶活性高于冰温组(图1),催化肌浆蛋白发生磷酸化反应的程度较强,而随着贮藏时间的延长,冷藏组由于糖酵解速率快[6],ATP消耗较快、含量降低,导致ATP依赖性蛋白激酶活性降低[24],肌浆蛋白磷酸化反应程度降低,磷酸化水平不再升高。相反,冰温贮藏有利于维持蛋白激酶活性,进而促进肌浆蛋白整体磷酸化水平的稳定。即使冷藏有利于肌浆蛋白整体磷酸化水平的提高,但并不是所有的冷藏处理组所有的肌浆蛋白磷酸化水平均高于冰温贮藏组。肌浆蛋白整体磷酸化水平受贮藏条件、贮藏时间及贮藏条件与贮藏时间的交互作用共同影响。
HUANG等[3]的质谱鉴定结果显示,条带3主要为糖原磷酸化酶,有研究表明糖原磷酸化酶等大多数糖酵解酶都可以在体外被不同激酶催化发生磷酸化反应,反应后酶活性或其稳定性增强,进而加速糖酵解进程[25-26],本研究中,冷藏组糖原磷酸化酶磷酸化水平高于冰温组,因此推测冰温贮藏会通过下调糖原磷酸化酶的磷酸化水平而使其活性降低,从而延缓糖酵解。条带4主要为磷酸果糖激酶,磷酸果糖激酶是糖酵解作用的限速酶,其发生磷酸化反应后活性下降,减少了2,6-二磷酸果糖的产生,从而抑制糖酵解进程,减缓肌肉pH下降速率,本试验结果为冰温组磷酸果糖激酶磷酸化水平高于冷藏组,所以冰温贮藏有利于通过上调磷酸果糖激酶磷酸化水平而使其活性升高,对糖酵解进程起到限制作用。条带7主要为丙酮酸激酶,其为糖酵解途径的限速酶,可催化磷酸稀醇丙酮酸向丙酮酸转化,进一步转变为乳酸,本研究结果为冷藏组丙酮酸激酶磷酸化水平高于冰温组,此外,李培迪等[6]的研究表明,冰温贮藏可上调丙酮酸激酶活力,同时延缓肌肉达到极限pH的时间,因此,认为冰温贮藏可通过调控肌肉中糖酵解酶的活性来影响其成熟进程。但另有研究表明,AMPK会抑制L-丙酮酸激酶的活性,从而抑制糖酵解,说明丙酮酸激酶通过多个途径影响糖酵解进程。条带8可能为AMPK,AMPK的激活会引起磷酸果糖激酶2的磷酸化反应,本试验结果为冰温组AMPK磷酸化水平高于冷藏组,说明冰温条件更有利于AMPK催化磷酸果糖激酶发生磷酸化反应,这也是冰温组磷酸果糖激酶磷酸化水平高于冷藏组的原因。条带13主要为乳酸脱氢酶,乳酸脱氢酶能够可逆地催化并氧化L-乳酸生成丙酮酸,李培迪[6]的研究表明冰温贮藏下调乳酸脱氢酶活力,本研究中,冰温贮藏中乳酸脱氢酶磷酸化水平低于冷藏条件中其磷酸化水平,因此认为冰温贮藏能够通过调控乳酸脱氢酶磷酸化水平而影响其活性,进而影响糖酵解。条带14可能为热休克蛋白27,该蛋白是涉及细胞生长及细胞凋亡等功能的重要蛋白,当细胞受到胁迫,热休克蛋白27表达量将增加,且会被磷酸化从而具有活性,行使保护肌动蛋白的功能,阻碍肌动蛋白与肌球蛋白的结合,进而影响嫩度,本试验中,冰温组热休克蛋白27的磷酸化水平高于冷藏组,说明冰温条件能够上调热休克蛋白27的磷酸化水平使其对肉品嫩度的调控作用发生改变。另外,REISS[27]、SALE[28]等的研究都表明猪背最长肌中的丙酮酸激酶、磷酸果糖激酶等糖酵解酶的活性均受自身磷酸化程度的影响,它们发生磷酸化反应后更加活跃,从而加快肌肉的糖酵解反应。因此,本研究结果表明,肌肉中各种糖酵解酶的磷酸化水平受冰温贮藏调控,导致其活性受到影响,进而通过影响宰后糖酵解进程调控肉品品质。
3.3 不同贮藏处理对肌原纤维蛋白磷酸化水平的影响
肌原纤维蛋白是骨骼肌蛋白中含量最多的蛋白,其含量占蛋白总量的55%—60%,肌原纤维蛋白与肌肉收缩特性相关,并且与肌肉功能特性及原料肉烹饪特性之间均存在很大联系[29]。Heeley[10]、Mazzei[11]、Ryder[12]等学者研究发现,肌间线蛋白、肌钙蛋白、肌动蛋白、原肌球蛋白及肌球蛋白轻链等多种肌原纤维蛋白都会发生蛋白质磷酸化反应。结合笔者课题组[30]及Huang等[9]的质谱试验结果,本试验选择了分析部分重要肌原纤维蛋白条带的磷酸化水平。
条带3中主要为肌球蛋白结合蛋白C,研究表明肌球蛋白结合蛋白C能够在蛋白激酶C作用下发生磷酸化反应,磷酸化的肌球蛋白结合蛋白C与肌动蛋白的相互作用将会减弱,不利于肌肉收缩[31],有利于肌肉嫩化,本试验结果为冷藏组蛋白激酶C磷酸化水平高于冷藏组,说明冰温贮藏可通过下调肌球蛋白结合蛋白C的磷酸化水平来延缓肌肉成熟。条带8、9可能为原肌球蛋白,原肌球蛋白主要功能为调控肌球蛋白与肌动蛋白之间的相互作用,WU等[32]研究表明蛋白激酶C可使原肌球蛋白的丝氨酸位点发生磷酸化反应,从而提高其磷酸化水平,本研究结果为,冷藏组原肌球蛋白磷酸化水平高于冰温组,说明冰温贮藏可能会通过影响原肌球蛋白的磷酸化水平而调控肌肉收缩。条带10与条带11可能为肌钙蛋白T3(TNNT3)与肌钙蛋白T1(TNNT1),肌钙蛋白是肌肉中主要的调节蛋白,直接参与钙离子调控的肌肉收缩[33-34],MOIR等[33]研究发现,肌钙蛋白T激酶、磷酸化酶b激酶、酪蛋白激酶TS以及Ca2+依赖性磷脂敏感蛋白激酶均可使肌钙蛋白T发生磷酸化反应,本研究中冰温组条带10、条带11的磷酸化水平均高于冷藏组,说明相较于冷藏,冰温贮藏可上调肌钙蛋白T的磷酸化水平,对肌肉收缩的强度与速度产生重要影响。条带16可能为肌钙蛋白I,ENGLAND[35]的研究表明,肌钙蛋白I的磷酸化现象能够促进肌肉收缩,本试验中冰温组条带16的磷酸化水平高于冷藏组,说明冰温条件更有利于肌钙蛋白I磷酸化水平的升高,有利于其对肌肉收缩的促进作用。条带20为肌球蛋白轻链2,STULL等[36]发现肌球蛋白轻链的磷酸化会引起肌球蛋白的结构改变,由此增加肌球蛋白的收缩强度,本试验中冰温组肌球蛋白轻链2磷酸化水平高于冷藏组,因此冰温贮藏引起肌球蛋白轻链磷酸化水平的升高,从而增强肌球蛋白的收缩。综上所述,冰温贮藏可通过影响肌原纤维蛋白中肌肉收缩相关主要蛋白的磷酸化水平,而影响肌肉收缩的速度与强度,从而调控肉品质。
4 结论
冰温及冷藏条件下蛋白激酶活性均随贮藏时间的延长而降低,但降低的程度不同,贮藏12 h—9 d时,冷藏组蛋白激酶活性始终显著低于冰温组。冷藏组肌浆蛋白整体磷酸化水平最大值出现在贮藏第3天,冰温组出现在第5天,且在贮藏早期(12 h—3 d)冷藏组显著高于冰温组,贮藏9 d时冰温组显著高于冷藏组。冷藏组肌原纤维蛋白整体磷酸化水平在贮藏24 h时达到最大值,冰温组最大值则出现在12 h。综上所述,冰温贮藏有利于降低蛋白质磷酸化水平,从而间接调控肉品质,可为宰后贮藏条件的优化提供参考。
References
[1] 孙天利. 冰温保鲜技术对牛肉品质的影响研究[D]. 沈阳: 沈阳农业大学, 2013.
Sun T L. Influences of controlled freezing point storage on beef [D]. Shenyang: Shenyang Agricultural University, 2013. (in Chinese)
[2] 钱小红, 贺福初. 蛋白质组学: 理论与方法. 北京: 北京科学出版社, 2003.
Qian X H, He F C.. Beijing: Beijing Science Press, 2003. (in Chinese)
[3] Huang H G, Larsen M R, Karlsson A H, Pomponio L, Costa L N, Lametsch R. Gel-based phosphoproteomics analysis of sarcoplasmic proteins in postmortem porcine muscle with pH decline rate and time differences., 2011, 11(20): 4063-4076.
[4] Gannon J, Staunton L, O’ connell K, Doran P, Ohlendieck K. Phosphoproteomic analysis of aged skeletal muscle., 2008, 22(1): 33-42.
[5] DOUMIT M E, BATES R O. Regulation of pork water holding capacity, color, and tenderness by protein phosphorylation., 2000, 63(1): 17-22.
[6] 李培迪, 李欣, 李铮, 陈丽, 李仲文, 陈丽娟, 田建文, 张德权. 冰温贮藏对宰后肌肉成熟进程的影响. 中国农业科学, 2016, 49(3): 554-562.
Li P D, Li X, Li Z, Chen L, Li Z W, Chen L J, Tian J W, Zhang D Q. Effects of controlled freezing point storage on aging from muscle to meat., 2016, 49(3): 554-562. (in Chinese)
[7] ANDERSON M J, LONERGAN S M, Huff-LONERGAN E. Differences in phosphorylation of phosphoglucomutase 1 in beef steaks from the longissimus dorsi with high or low star probe values., 2014, 96(1): 379-384.
[8] SHEN Q W, MEANS W J, UNDERWOOD K R, THOMPSON S A, ZHU M J, MCCORMICK R J, FORD S P, ELLIS M, DU M. Early post-mortem AMP-activated protein kinase (AMPK) activation leads to phosphofructokinase-2 and -1 (PFK-2 and PFK-1) phosphorylation and the development of pale, soft, and exudative (PSE) conditions in porcine longissimus muscle., 2006, 54(15): 5583-5589.
[9] HUANG H G, LARSEN M R, LAMETSCH R. Changes in phosphorylation of myofibrillar proteins during postmortem development of porcine muscle., 2012, 134(4): 1999-2006.
[10] HEELEY D H, WATSEN M H, MAK A S, DUBORD P, SMILLI L B. Effect of phosphorylation on the interaction and functional properties of rabbit striated-muscle alpha-alpha-tropomyosin., 1989, 264(5): 2424-2430.
[11] MAZZEI G J, KUO J F. Phosphorylation of skeletal-muscle troponin I and troponin T by phospholipid-sensitive Ca2+-dependent protein kinase and its inhibition by troponin C and tropomyosin., 1984, 218(2): 361-369.
[12] RYDER J W, LAU K S, KAMM K E, STULL J T. Enhanced skeletal muscle contraction with myosin light chain phosphorylation by a calmodulin-sensing kinase., 2007, 282(28): 20447-20454.
[13] 尹靖东. 动物肌肉生物学与肉品科学. 北京: 中国农业大学出版社, 2011.
Yin J D.. Beijing: China Agricultural University Press, 2011. (in Chinese)
[14] LAMETSCH R, KRISTENSEN L, LARSEN M R, THERKIDSEN M, OKSBIERG N, ERTBJERG P. Changes in the muscle proteome after compensatory growth in pigs., 2006, 84: 918-924.
[15] SMUDER A J, KAVAZIS A N, HUDSON M B, NELSON W B, POWER S K. Oxidation enhances myofibrillar protein degradation via calpain and caspase-3., 2010, 49(7): 1152-1160.
[16] MANNING G, WHYTE D B, MARTINEZ R, HUNTER T, SUDARSANAM S. The protein kinase complement of the human genome., 2002, 298(5600): 1912-1934.
[17] ROBERT R J. A historical overview of protein kinases and their targeted small molecule inhibitors., 2015, 7(10): 1-23.
[18] BARON S J, LI J, RUSSELL 3rd R R, NEUMANN D, MILLER E J, TUERK R, WALLIMANN T, HURLEY R L, WITTERS L A, YOUNG L H. Dual mechanisms regulating AMPK kinase action in the ischemic heart., 2005, 96(3): 337-345.
[19] HWANG J T, HA J, PARK O J. Combination of 5-fluorouracil and genistein induces apoptosis synergistically in chemo-resistant cancer cells through the modulation of AMPK and COX-2 signaling pathways., 2005, 332(2): 433-440.
[20] HWANG J T, LEE M, JUNG S N, LEE H J, KANG I, KIM S S, HA J. AMP-activated protein kinase activity is required for vanadate- induced hypoxia-inducible factor 1alpha expression in DU145 cells., 2004, 25(12): 2497-2507.
[21] HAWLEY S A, PAN D A, MUSTARD K J, ROSS L, BAIN J, EDELMAN A M, FRENGUELLI B G, HARDIE D G. Calmodulin- dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinase., 2005, 2(1): 9-19.
[22] WOODS A, DICKERSON K, HEATH R, HONG S P, MIMCILOVIC M, JOHNSTONE SR, CARLSON M, CARLING D. Ca2+/ calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells., 2005, 2(1): 21-33.
[23] SCOPES, R K, STOTER A. Purification of all glycolytic enzymes from one muscle extract., 1982, 90: 479-490.
[24] DICKENS J A, LYON C E. The effects of electric stimulation and extended chilling times on the biochemical reactions and texture of cooked broiler breast meat., 1995, 74(12): 2035-2040.
[25] SAVELL J W, MUELLER S L, BAIRD B E. The chilling of carcasses., 2005, 70(3): 449-459.
[26] WALSH D A, PERKINS J P, KREBS E G. An adenosine 3’,5’-monophosphatedependant protein kinase from rabbit skeletal muscle., 1968, 243(13): 3763-3774.
[27] REISS N, KANETY H, SCHLESSINGER J. Five enzymes of the glycolytic pathway serve as substrates for purified epidermal- growth-factor-receptor kinase., 1986, 239(3): 691-697.
[28] SALE E M, WHITE M F, KAHN C R. Phosphorylation of glycolytic and gluconeogenic enzymes by the insulin receptor kinase., 1987, 33(1): 15-26.
[29] LEE S H, JOO S T, RYU Y C. Skeletal muscle fiber type and myofibrillar proteins in relation to meat quality., 2010, 86(1): 166-170.
[30] CHEN L J, LI X, NI N, LIU Y, CHEN L, WANG Z Y, SHEN Q W, ZHANG D Q. Phosphorylation of myofibrillar proteins in post- mortem ovine muscle with different tenderness., 2016, 96(5): 1474-1483.
[31] WANG L, SADAYAPPANAPPAN S, KAWAI M. CARDIAC. Myosin binding protein C phosphorylation affects cross-bridge cycle’s elementary steps in a site-specific manner., 2014, 9(11): e113417.
[32] WU S C, SOLARO R J. Protein kinase C zeta. A novel regulator of both phosphorylation and de-phosphorylation of cardiac sarcomeric proteins., 2007, 282(42): 30691-30698.
[33] MOIR A J, PERRY S V. The sites of phosphorylation of rabbit cardiac troponin I by adenosine 3’,5’-cyclic monophosphate-dependent protein kinase: Effect of interaction with troponin C., 1977, 167(2): 333-343.
[34] PINNA L A, MEGGIO F, DEDIUKINA M M. Phosphorylation of troponin T by casein kinase TS., 1981, 100(1): 449-454.
[35] ENGLAND P J. Correlation between contraction and phosphorylation of the inhibitory subunit of troponin in perfused rat heart., 1975, 50(1): 57-60.
[36] STULL J T, KAMM K E, VANDENBOOM R. Myosin light chain kinase and the role of myosin light chain phosphorylation in skeletal muscle., 2011, 510(2): 120-128.
(责任编辑 赵伶俐)
Effects of Controlled Freezing Point Storage on the Protein Phosphorylation Level
ZHANG Yan1,2, LI Xin2, LI Zheng2, LI Meng2, LIU Yong-feng1, ZHANG De-quan2
(1College of Food Engineering and Nutritional Science, Shaanxi Normal University, Xi’an 710119;2Institute of Food Science and Technology, Chinese Academy of Agricultural Sciences/Key Laboratory of Agro-Products Processing, Ministry of Agriculture, Beijing 100193)
【Objective】 The aim of this study was to investigate the effect of controlled freezing point on protein phosphorylation level, so as to provide a new theoretical basis for controlling meat quality by controlled freezing point. 【Method】 The longissimus doris (LD) of large tail×small tail crossed sheep were obtained and divided into two groups: refrigeration group and controlled freezing point group. Each LD was assigned to 6 storage times (0.5 h, 12 h, 24 h, 3 d, 5 d and 9 d). Kit for protein kinase activity measurement and enzyme linked immunosorbent assay were applied to detect protein kinase activity. A gel-based phosphoproteomic study was performed. And SDS-PAGE electrophoresis and fluorescence staining were applied to gain the protein bands of sarcoplasmic proteins and myofibrillar proteins stained by SYPRO Ruby and Pro-Q Diamond. A software named Quantity One was used to analyze the protein phosphorylation level by measuring the fluorescence intensity.【Result】 At 12 h, the protein kinase activity of controlled freezing point group significantly increased (<0.05) and that of refrigeration group significantly decreased. The protein kinase activity of the two groups were reduced from 12h to 9 d, which was significantly higher in controlled freezing group compared with refrigeration group (<0.05). Seventeen protein bands distributed from 15-250 kDa in each sarcoplasmic protein sample were analyzed and the results suggested that storage temperature influenced (<0.001) the protein phosphorylation level of all sarcoplasmic proteins significantly. The total phosphorylation level of sarcoplasmic proteins in refrigeration group increased at first and then decreased while the sarcoplasmic proteins in controlled freezing point group showed an opposite variation trend. The phosphorylation level of 3 d was the highest, while the lowest was 24 h. Globally, the sarcoplasmic proteins of refrigeration group had the higher total phosphorylation level than controlled freezing point group during 12 h to 3 d. However, the total phosphorylation level of the sarcoplasmic proteins of controlled freezing point was higher than refrigeration group on 9 d. Twenty protein bands distributed from 15-250 kDa in each myofibrillar protein sample were analyzed and the results suggested storage temperature influenced the protein phosphorylation level of all myofibrillar proteins very significantly (<0.001). The total phosphorylation level of myofibrillar proteins increased firstly and then decreased in two groups. And the highest phosphorylation level of 24 h in refrigeration group and 12 h in controlled freezing point group was the highest. Total phosphorylation level of myofibrillar protein had no significant difference between two groups from 0.5 h to 12 h. However, total phosphorylation level of myofibrillar protein was higher in refrigeration group compared with controlled freezing group from 24 h to the end of storage.【Conclusion】 The controlled freezing point storage regulated the phosphorylation level of protein by affecting the activity of protein kinase, which can indirectly regulate the meat quality by influencing the glycometabolism, muscle contraction and degradation of skeleton protein.
controlled freezing point; protein kinase; sarcoplasmic proteins; myofibrillar proteins; protein phosphorylation
2016-04-18;接受日期:2016-07-29
国家公益性行业(农业)科研专项(201303083)、国家农业科技创新工程、陕西省科技统筹创新计划(2015KTTSNY04-07)
张艳,E-mail:zhangwen2255@126.com。通信作者刘永峰,E-mail:yongfeng200@126.com。通信作者张德权,E-mail:dequan_zhang0118 @126.com
