亚高山草甸植物群落对气候变化的响应
2016-10-14庞晓瑜雷静品邓云鹏
庞晓瑜,雷静品*,王 奥,邓云鹏
(1 中国林业科学研究院林业研究所,国家林业局林木培育重点实验室,北京 100091;2 北京林业大学自然保护区学院,北京 100083)
亚高山草甸植物群落对气候变化的响应
庞晓瑜1,雷静品1*,王 奥2,邓云鹏1
(1 中国林业科学研究院林业研究所,国家林业局林木培育重点实验室,北京 100091;2 北京林业大学自然保护区学院,北京 100083)
为了研究气温升高、氮素增加和人为干扰对亚高山草甸植物生长和非结构性碳水化合物(NSC)的影响,该研究采用开顶式生长箱(OTC)模拟增温,同时进行施加氮肥和除草处理,对青藏高原东南缘邛崃山脉东坡巴郎山(四川盆地向青藏高原的过渡地带)的亚高山草甸植物的生长和NSC含量进行测定分析。结果显示:(1)各处理土壤全磷(P)和全钾(K)含量与对照均无显著差异,增温加施肥处理的土壤全氮(N)含量与对照无显著差异,但增温处理、施肥处理、除草处理、增温加除草处理、施肥加除草处理和增温加施肥加除草处理的土壤全氮含量较对照均显著降低。(2)增温促进禾本科和杂类草功能群生长,抑制莎草科功能群生长,提高禾本科功能群重要值,降低杂类草功能群重要值,且对莎草科功能群重要值的作用受施氮和除草的影响;施肥促进禾本科和杂类草功能群的高生长,并且促进莎草科功能群生长;除草促进莎草科功能群生长,抑制禾草科和杂类草功能群的生长;而施肥和除草的交互作用有利于禾草科功能群生长,施肥和除草都提高了莎草科功能群的重要值,降低了禾草科功能群的重要值。(3)不同物种NSC含量及分配对于各处理的响应有所不同,紫地榆的NSC含量与物种分盖度相关性显著,珠芽蓼的NSC含量与物种高度相关性显著。研究表明,气候变暖和土壤氮素增加有利于禾本科和莎草科植物的生长,并使植物改变体内非结构性碳水化合物的分配来抵御环境压力。
亚高山草甸;模拟增温;氮素添加;非结构性碳水化合物
全球气候变化毋庸置疑[1],IPCC第五次评估报告的预测显示,未来20年,全球预计增温0.3~0.7 ℃,截止本世纪末可能增温0.3~4.8 ℃[2]。温度升高能够改变土壤特性等环境因子及植物的生理过程,从而影响植物的生长发育,进而影响植物群落的组成、结构及生产力[3-5]。高纬度和高海拔地区的植被对温度的升高更为敏感[6],其植被和生态系统对温度变化的响应一直是生态学研究的重点。大量野外试验表明增温能促进植被的个体生长和群落高度,而群落组成、生产力及多样性等方面,因试验地点、群落类型、物种组成以及试验时间等因素的影响,目前尚没有一致的结论[7-9]。其次,氮素是植物生长所需要的重要元素,适宜的外源氮素能促进植物生长和生物量的积累,但过量的氮素对植物的生长有害[10],外源氮素对植物生产力的促进与否取决于生态系统的氮素是否饱和[11]。有研究表明,外源氮素的添加使喜氮植物在竞争中更具优势,从而影响群落的组成结构和多样性,并且,外源氮素的添加能降低群落丰富度和物种多样性[12-13]。另外,人为干扰不仅直接影响植物群落,还会影响气候变化和氮素添加等环境因子对植物群落的作用[14]。目前人为干扰对高寒草甸影响的研究多集中在放牧上,但其他人类活动的影响(如采药)研究较少[15-16]。在植被资源丰富的山区,药材采集和野菜挖掘时有发生[17],但这种干扰对亚高山草甸的影响尚不清楚。我们通过模拟增温、施肥和除草试验,研究亚高山草甸植物生长和非结构性碳水化合物含量的响应,探讨气候变暖、氮素增加和人为干扰对亚高山草甸的影响。
1 试验地概况与方法
1.1 试验地概况
试验区位于四川省阿坝藏族羌族自治州,汶川藏族羌族自治县境内,地处青藏高原东南缘邛崃山脉东坡巴郎山,属四川盆地向青藏高原的过渡地带。该地区雨水充沛,气温年差较小,昼夜温差较大,干湿季较为明显,垂直气候差异较大,年均温8.4 ℃,年均降水量862 mm,且降水主要集中于夏季。土壤棕褐色,土层较薄,土内砾石较多,表层富含有机质,土壤pH值偏酸性。该区植物资源丰富,样地内的主要植物有:垂穗披碱草(Elymusnutans)、黑穗画眉草(Eragrostisnigra)、珠芽蓼(Polygonumviviparum)、甘青大戟(Euphorbiamicractina)、紫地榆(Geraniumstrictipes)、驴蹄草(Calthapalustris)、草玉梅(Anemonerivularis)、野草莓(Fragariavesca)、葱状灯心草(Juncusallioides)、川陕金莲花(Trolliusbuddae)等。在该地区,当地农民除耕种农田外,还从事放牧(绵羊和牦牛)和药材采集,如川贝(Fritillariacirrhosa)。
1.2 试验设计与取样方法
试验样地设在巴郎山阳坡的亚高山草甸(30°53′N,102°58′E,海拔3 400 m)。采用开顶式生长箱(open-top chamber, OTC)模拟增温,氮素添加施用尿素(施氮量120 kg/hm2),用将植株拔起的方式模拟人为采药活动。设置增温施肥除草(TNC)、增温施肥不除草(TN)、增温不施肥除草(TC)、增温不施肥不除草(T)、不增温施肥除草(NC)、不增温施肥不除草(N)、不增温不施肥除草(C)以及不增温不施肥不除草(CK)共8个处理,重复4次,一共设置32个样方,样方间隔大于1 m,样方为直径60 cm的六边形。选取4个增温样方和4个不增温样方安装空气温湿度记录仪(离地面30 cm,HOBO Pro v2 U23-001型,Onset Computer Cor.,USA)监控温湿度变化。试验于2015年4月布置完成。于2015年7月14日至24日调查并记录每个样地内各物种的数量、高度、盖度,少于10个的物种记录所有植株的高度,大于10个的物种,随机选取10株记录其高度,计算重要值[重要值=(相对多度+相对频度+相对盖度+相对高度)/4]。并选取多数样方均有出现的常见种甘青大戟、紫地榆和珠芽蓼取样带回,在75 ℃烘箱中烘干至恒重,研磨过筛,用于测定非结构性碳水化合物。于2015年10月14日在每个样地用直径7 cm的土钻取3个土柱,去掉根系,烘干土壤,过100目筛,用于测定土壤氮、磷、钾含量。
1.3 样品测定与数据处理
植物样品用蒽酮比色法测定非结构性碳水化合物含量。土壤样品用凯氏定氮仪测定全氮含量,用钼锑抗比色法测定全磷含量,用火焰光度计法测定全钾含量。用Excel 2013及SPSS v19进行数据处理。
2 结果与分析
2.1 OTC对温度和湿度的影响
通过增温处理,OTC内平均增温0.893 ℃,日均温最高增加2.484 ℃;空气湿度平均增加了8.494%,日平均湿度最高增加32.136%,最高降低8.169%。OTC的设置使空气温度显著升高,但空气湿度在5、6月份显著低于对照,在7、8、9和10月份显著高于对照(P<0.05)。OTC的设置并未改变
温度月均温的变化趋势,对照和增温处理的月平均温度都是先升高后降低,对照样地的月平均温度最高值出现在7月,而OTC内则提前至6月(表1)。但OTC改变了空气月平均湿度的变化趋势,对照中空气湿度在生长季内整体呈现先降低后上升的趋势,夏季湿度最低,而OTC内则呈现先上升后下降的趋势,夏季湿度高于春秋两季(表1)。温度在白天先升高后降低,在夜间持续降低,于12∶00达到最高,OTC内和对照的温度差值也随温度增加而增加(图1)。与空气温度相反,空气湿度则在白天呈现先降低后增高的变化趋势,差值最低值出现于午后两点左右。
2.2 增温、施肥、除草对土壤氮、磷、钾含量的影响
各处理间土壤全钾(K)的含量无显著差异(图2)。土壤全P含量NC处理(0.047%)和TNC处理(0.091%)间差异显著(P<0.05),但其余处理间均无显著差异。TN处理的土壤全氮(N)含量(0.752%)高于对照(0.728%),其余处理均低于对照。增温和除草对土壤N含量的影响均达到显著水平(P<0.05),增温和施肥的交互作用以及增温施肥和除草三者的交互作用均达到了显著水平(P<0.05)。这说明除草和增温影响土壤N素含量,而在增温条件下施肥对土壤N影响显著,而P和K则受处理影响较小或不受影响。
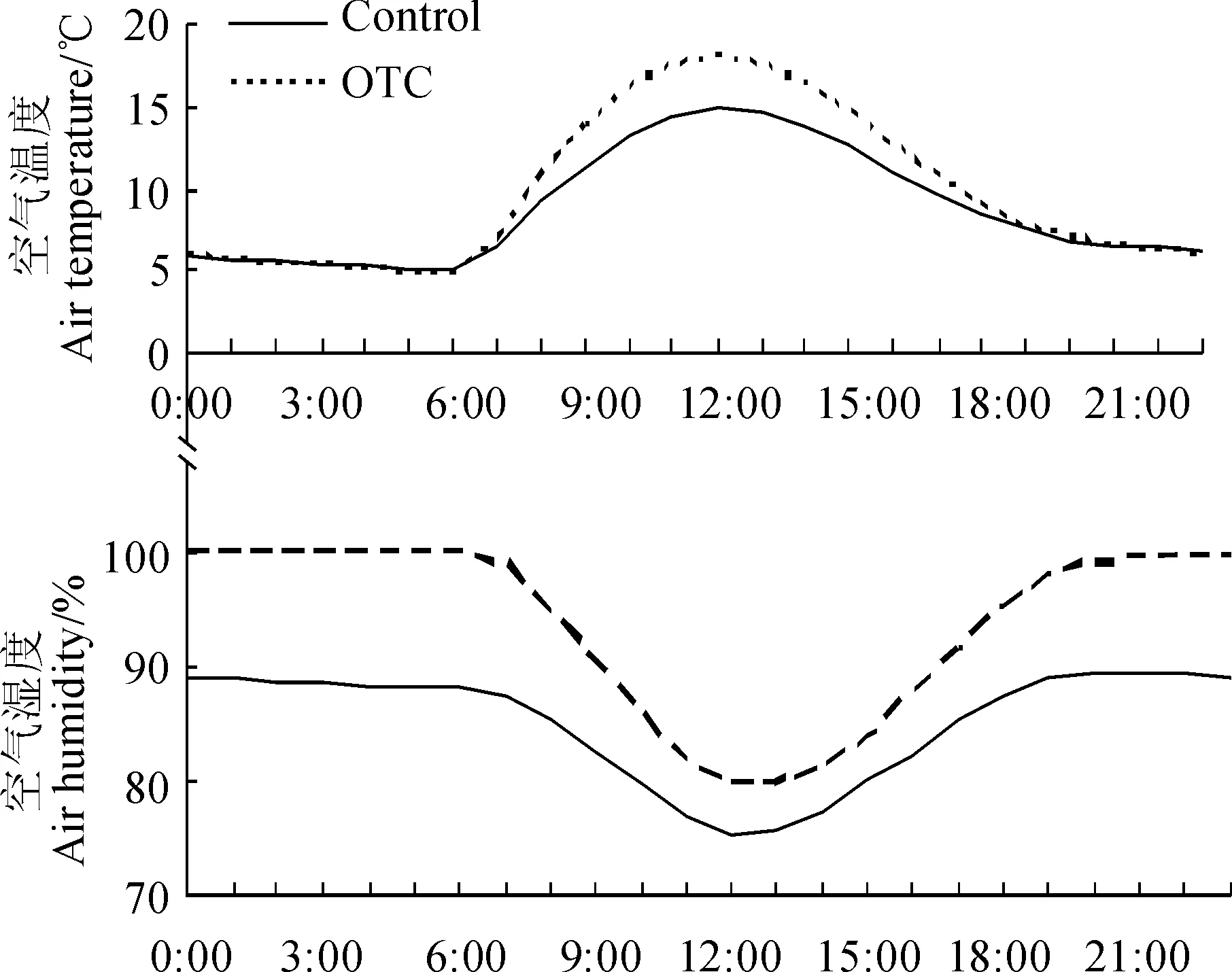
图1 空气温度和空气湿度日变化Fig. 1 Diumal variation of air temperature and air humidity
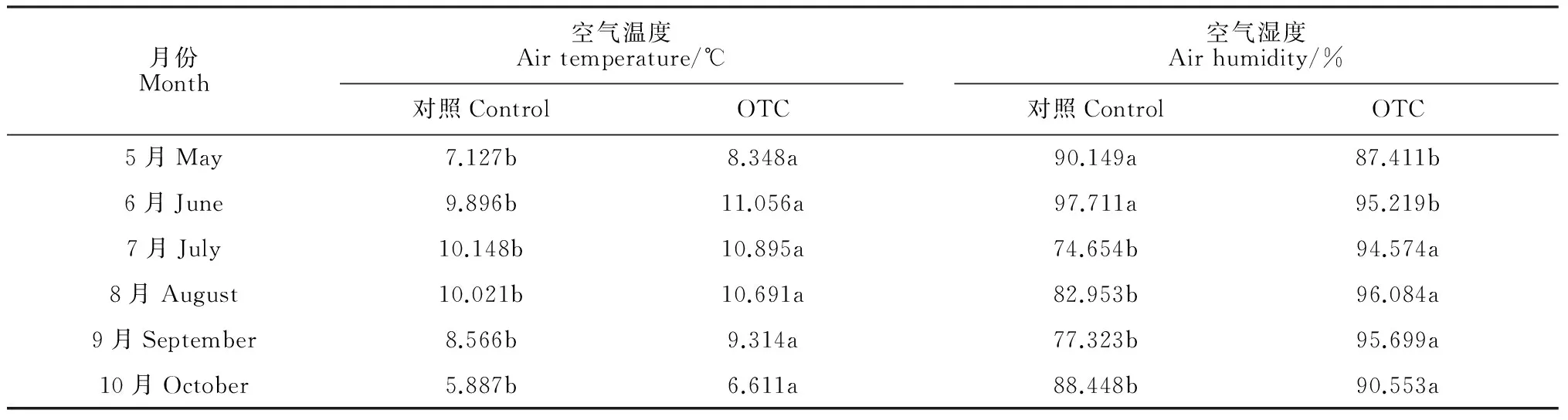
月份Month空气温度Airtemperature/℃空气湿度Airhumidity/%对照ControlOTC对照ControlOTC5月May7.127b8.348a90.149a87.411b6月June9.896b11.056a97.711a95.219b7月July10.148b10.895a74.654b94.574a8月August10.021b10.691a82.953b96.084a9月September8.566b9.314a77.323b95.699a10月October5.887b6.611a88.448b90.553a
注:不同字母表示处理间差异显著(P<0.05)
Note: The different letters indicate significant difference inside and outside the OTCs (P<0.05)
2.3 增温、施肥、除草对不同功能群数量特征的影响
表2显示,增温使禾本科的盖度、高度和重要值普遍升高,仅TNC处理中禾本科高度小于NC处理,增温在不施肥条件下提高禾本科多度,在施肥条件下降低禾本科多度。施肥在不除草条件下降低禾本科盖度和重要值,在除草条件下提高禾本科盖度和重要值,在不增温条件下提高禾本科多度,在增温条件下降低禾本科多度;同时,施肥普遍提高禾本科高度,只有TNC处理的禾本科高度低于TC处理。除草降低禾本科的高度和多度,并且在不施肥条件下降低禾本科盖度和重要值,在施肥条件下提高禾本科盖度和重要值。这说明增温有利于禾本科的生长和群落中地位的提升,施加氮素对禾本科的高生长有利,对径向生长不利,而除草抑制禾本科生长。同时,增温和施肥的交互作用降低禾本科植株数量,施氮和除草的交互作用有利于禾本科的径向生长,并且氮素对禾本科植物的作用受其他环境条件的影响较大。
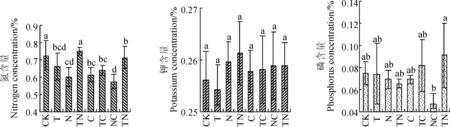
CK. 不增温、不施肥、不除草;T. 增温、不施肥、不除草; N. 不增温、施肥、不除草;TN. 增温、施肥、不除草;C. 不增温、不施肥、除草;TC. 增温、不施肥、除草;NC. 不增温、施肥、除草;TNC. 增温、施肥、除草;不同字母表示处理间差异显著(P<0.05);下同。图2 各处理对土壤全氮、全磷和全钾含量的影响CK. without warming, without fertilization, without weeding; T. warming, without fertilization, without weeding; N. without warming, fertilization, fertilization, without weeding; TN. warming, fertilization, without weeding; C. without warming, without fertilization, without weeding; TC. warming, without fertilization, weeding; NC. without warming, fertilization, weeding; TNC. warming, fertilization, weeding; The different letters indicate significant difference among the treatments (P<0.05); The same as belowFig. 2 The contents of soil total phosphorus (P), total potassium (K) and total nitrogen (N) in the treatments

功能群Functionalgroup项目Item处理TreatmentTNCNCTCCTNNTCK禾本科GrassesⅠ35.000ab25.250b30.500ab13.750b27.250ab24.250ab55.500a28.750abⅡ9.000a11.500a14.000a9.500a10.750a22.250a34.750a20.750aⅢ37.750ab47.147ab47.068ab21.764b65.592a53.564ab59.474a47.549abⅣ19.14413.06614.74110.90217.08112.96419.38314.728莎草科SedgesⅠ45.000a33.250a31.25027.500a30.750a32.500a15.000a29.500aⅡ5.333a5.500a6.500a7.000a5.000a6.000a2.750a6.750aⅢ21.493bc20.109bc20.436bc14.763c31.764a25.860ab16.786bc19.413bcⅣ5.7925.2353.9124.8674.2284.5072.4163.782杂类草FordsⅠ201.167b162.875b288.500a170.875b284.375a234.750ab299.250a290.500aⅡ53.000b54.000b78.750ab65.750ab65.000ab70.750ab94.750a88.500aⅢ25.740ab27.894ab25.536ab15.730b42.475a40.145a33.606ab29.318abⅣ72.85978.56878.64180.24076.49479.37076.86378.540小灌木SmallshrubⅠ2.667a12.000a14.500a17.000a8.750a10.000a3.250a15.000aⅡ0.333b2.000ab1.750ab2.250ab1.000ab1.000ab0.500b3.000aⅢ36.000a27.917a29.500a26.778a38.500a30.250a24.000a29.417aⅣ2.2053.1312.7063.9922.1973.1591.3382.950
注:Ⅰ. 盖度;Ⅱ. 多度;Ⅲ. 高度;Ⅳ. 重要值;不同字母表示处理间差异显著(P<0.05)
Note: Ⅰ. Coverage;Ⅱ. Abundance;Ⅲ. Height;Ⅳ. Importance value; The different letters indicate significant difference among the treatments (P<0.05)
增温降低莎草科多度;增温处理中莎草科的高度小于对照,而在施肥、除草及施肥和除草条件下,增温均使莎草科高度升高;增温在不除草条件下降低莎草科盖度,在除草条件下提高莎草科盖度;增温普遍降低莎草科重要值,但TNC处理中莎草科的重要值大于NC处理。氮素添加提高了莎草科的盖度、高度和重要值,降低了莎草科的多度。除草提高莎草科的多度和重要值;除草普遍提高了莎草科盖度,只有C处理中莎草科盖度低于对照;同时,除草普遍降低莎草科的高度,只有TC处理莎草科高度大于T处理。这说明增温抑制了莎草科的生长,但除草和施氮可能因为改善了光照和养分条件,对莎草科的生长有利。
增温提高了杂类草的盖度,降低了杂类草的重要值;同时,增温普遍提高杂类草的高度,仅有TNC处理中杂类草高度低于NC处理;在不施氮条件下,增温提高杂类草多度,而在施氮条件下,增温降低杂类草多度。施氮提高杂类草的高度,却降低杂类草的多度和盖度;施肥普遍降低杂类草重要值,仅N处理中杂类草重要值高于对照。除草降低杂类草的盖度、高度和多度;并且,除草在不施肥条件下提高杂类草的重要值,在施肥条件下降低杂类草的重要值。这说明增温促进杂类草生长,施氮促进杂类草的高生长,而除草抑制杂类草生长,同时增温和施肥降低杂类草的优势度。
2.4 增温、施肥、除草对3种常见物种非结构性碳水化合物的影响
紫地榆可溶性糖浓度高于淀粉浓度,茎中的NSC浓度,尤其是可溶性糖浓度高于叶(图3)。TN处理的紫地榆茎中可溶性糖和淀粉浓度均显著高于对照(P<0.05),但TN处理紫地榆叶中NSC浓度显著低于对照(P<0.05),说明在增温和施肥的同时作用下,紫地榆更多地将非结构性碳水化合物向茎中运输。叶中淀粉浓度在TC处理中显著升高(P<0.05),在其余处理间无显著差异。在不除草的条件下,增温使紫地榆茎中非结构性碳水化合物含量升高,但在除草条件下,增温使紫地榆茎中非结构性碳水化合物含量降低;而增温使叶中非结构性碳水化合物的浓度在不除草的条件下降低,在除草条件下升高。除草普遍降低紫地榆茎中非结构性碳水化合物的含量,在不增温条件下除草降低紫地榆叶中非结构性碳水化合物含量,在增温条件下除草提高紫地榆叶中非结构性碳水化合物含量。
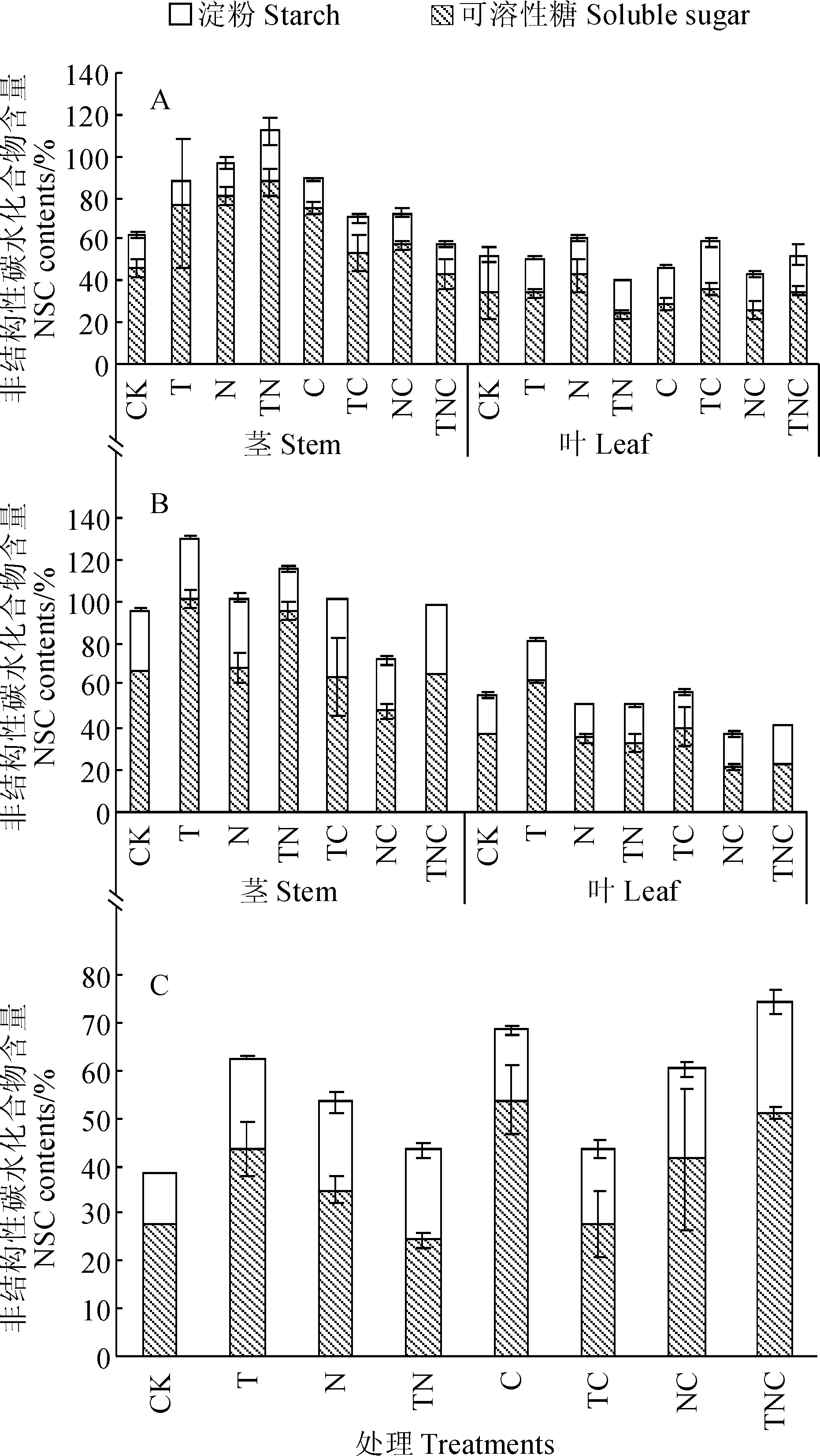
A. 紫地榆;B. 甘青大戟;C. 珠芽蓼图3 各处理间3个物种非结构性碳水化合物含量的变化A. Geranium strictipes;B. Euphorbia micractina;C. Polygonum viviparumFig. 3 Variation of non-structural carbohydrate contents of three species among different treatments
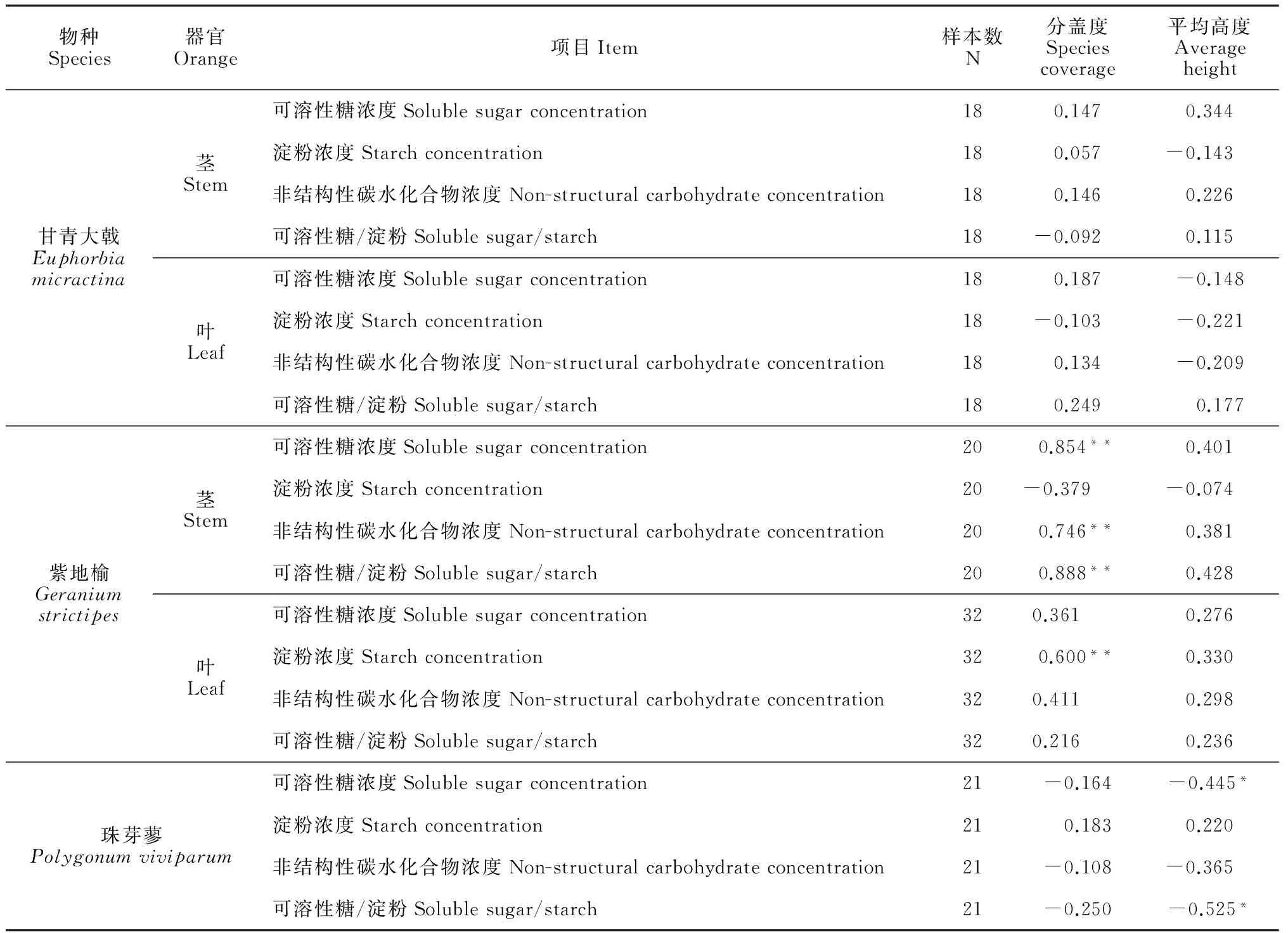
物种Species器官Orange项目Item样本数N分盖度Speciescoverage平均高度Averageheight甘青大戟Euphorbiamicractina茎Stem可溶性糖浓度Solublesugarconcentration180.1470.344淀粉浓度Starchconcentration180.057-0.143 非结构性碳水化合物浓度Non-structuralcarbohydrateconcentration180.1460.226可溶性糖/淀粉Solublesugar/starch18-0.092 0.115叶Leaf可溶性糖浓度Solublesugarconcentration180.187-0.148淀粉浓度Starchconcentration18-0.103 -0.221非结构性碳水化合物浓度Non-structuralcarbohydrateconcentration180.134-0.209可溶性糖/淀粉Solublesugar/starch180.249 0.177紫地榆Geraniumstrictipes茎Stem可溶性糖浓度Solublesugarconcentration200.854**0.401淀粉浓度Starchconcentration20-0.379 -0.074 非结构性碳水化合物浓度Non-structuralcarbohydrateconcentration200.746**0.381可溶性糖/淀粉Solublesugar/starch200.888**0.428叶Leaf可溶性糖浓度Solublesugarconcentration320.361 0.276淀粉浓度Starchconcentration320.600**0.330非结构性碳水化合物浓度Non-structuralcarbohydrateconcentration320.411 0.298可溶性糖/淀粉Solublesugar/starch320.216 0.236珠芽蓼Polygonumviviparum可溶性糖浓度Solublesugarconcentration21-0.164-0.445*淀粉浓度Starchconcentration21 0.1830.220非结构性碳水化合物浓度Non-structuralcarbohydrateconcentration21-0.108-0.365 可溶性糖/淀粉Solublesugar/starch21-0.250-0.525*
注:**表示极显著(P<0.01),*表示显著(P<0.05)
Note:**mean extremely significant difference (P<0.01),* means significant difference (P<0.05)
甘青大戟的可溶性糖浓度高于淀粉浓度,茎中NSC浓度大于叶中NSC浓度(图3)。T处理和TN处理中甘青大戟茎中可溶性糖显著高于对照,NC处理则显著低于对照;TC和N处理中茎的淀粉浓度显著高于对照(P<0.05)。T、TN和TNC处理的茎中可溶性浓度高于CK、N和NC处理,NC、TC和TNC处理的可溶性糖浓度小于N、T和TN处理,即甘青大戟茎中可溶性糖浓度在增温条件下升高,在除草条件下降低。T处理中甘青大戟叶的可溶性糖和淀粉浓度均显著高于对照,NC样地可溶性糖浓度显著低于对照(P<0.05)。NC、TC和TNC处理下甘青大戟叶中可溶性糖浓度分别低于N、T、TN处理,说明甘青大戟叶中可溶性糖浓度在除草条件下降低。
珠芽蓼中可溶性糖浓度高于淀粉浓度(图3)。C和TNC处理的可溶性糖浓度显著高于对照(P<0.05),淀粉浓度对照样地最低。C、NC和TNC处理中珠芽蓼的可溶性糖浓度和淀粉浓度高于CK、N、和NC处理,说明除草普遍提高珠芽蓼可溶性糖和淀粉浓度;同理,增温提高珠芽蓼可溶性糖浓度,施肥普遍提高了珠芽蓼的淀粉浓度。
将3种植物的可溶性糖浓度、淀粉浓度、非结构性碳水化合物浓度及可溶性糖与淀粉的比值分别与其物种分盖度及物种平均高度做相关性分析(表3)。紫地榆茎中NSC浓度、可溶性糖浓度及可溶性糖与淀粉的比值与物种分盖度有极显著的正相关关系,紫地榆叶中的淀粉浓度也与物种分盖度有极显著的正相关关系,这表示随着物种分盖度的增加,紫地榆将在叶中储存更多的淀粉,向茎中运输更多的可溶性糖;珠芽蓼的可溶性糖浓度及可溶性糖与淀粉的比值与物种平均高有显著的负相关关系,即随着高度的增加,珠芽蓼倾向于减少可溶性糖浓度,增加淀粉的储存。
3 讨 论
OTC的设置使样方温度增高,同时间接改变了湿度、风速、辐射、光照等环境条件,对植物生长发育产生影响。不同植物对于温度的敏感程度不同,一些物种对增温的响应更为迅速,导致群落结构发生变化[18-19]。本试验结果显示,增温能促进禾本科和杂类草的生长,抑制莎草科生长,同时促进群落向以禾本科植物为优势种的方向发展,增温条件下莎草科和杂类草在群落中的地位降低。这可能是因为禾本科植物处于群落上层,又对增温的响应比较敏感,其生长受温度促进明显,对群落下层的莎草科植物和部分杂类草的遮荫作用增强。赵建中等[20]在对莎草科为主的矮蒿草草甸的研究中表明增温后,禾本科对温度变化的响应最为敏感,莎草科次之,增温有利于杂类草的营养生长,不利于其生殖生长,并认为禾本科将取代莎草科的地位成为优势种,而部分杂类草将被淘汰,与本试验结果一致。张法伟[21]的研究也表明禾本类植物对温度呈现正响应,而莎草类植物则呈现负响应。
本试验显示施氮促进莎草科生长并促进禾本科和杂类草的高生长,降低杂类草和禾本科植物的盖度,提高禾本科和莎草科重要值,降低杂类草重要值,武建双[22]对垂穗披碱草群落的研究结果表明施氮能促进群落生长,曹文侠等[23]的研究表明施肥会显著降低杂类草盖度,增加禾本科植物的盖度,与本试验结果一致。郭红玉[24]在三江源对5种植物的研究表明单独增温和施氮对不同植物的高度影响不同,但同时施肥和增温能促进植物的生长,本实验中氮素添加和温度升高的共同作用提高了植株高度,但对盖度和多度的影响因功能群的不同而不同。
非结构性碳水化合物分为可溶性糖和淀粉,是植物体内重要的能源物质,其浓度不仅与植物的生长代谢相关,也关系到植物对碳的分配与利用[25]。可溶性糖不仅为植物生长提供物质和能量,同时也是调节物质的分子信号,其含量高低反映了碳吸收和消耗间的关系,而植物体内NSC浓度会在低温、干旱、盐胁迫等不利条件下升高,以平衡植株所受的胁迫[26]。李娜妮[27]等研究发现,从温带到热带,温度逐渐升高,森林常见树种叶片NSC浓度逐渐下降;Günter Hoch[28]等也发现,随着海拔的升高,温度逐渐降低,樟子松各组织中NSC浓度逐渐升高;但Gough[29]等的研究却显示温度升高会促进杨树和栎树的NSC积累;毛子军等[30]对蒙古栎幼苗进行增温处理,发现增温后其叶和茎的NSC浓度升高,但因幼苗NSC浓度根部所占比重最大,且根部NSC浓度下降,所以蒙古栎幼苗整体NSC浓度下降;说明不同植株、不同组织中的NSC浓度对增温的响应有所不同。本试验中,增温提高了甘青大戟茎和叶片以及珠芽蓼基生叶的NSC浓度,紫地榆茎中NSC浓度有所升高,叶中却有所降低,这可能是因为取样时正值紫地瑜花期,叶片产生的NSC更多地供应生殖生长,而NSC通过茎从叶运输花,使得茎中NSC含量上升而叶中NSC含量降低。
施氮处理下,甘青大戟NSC浓度降低,与蒋思思等[31]对油松幼苗的研究结果和王雪等[32]研究对大针茅和羊草的研究结果相同,而珠芽蓼和紫地榆的NSC浓度有所上升;杨焕文等[33]对烤烟的研究发现不同的施氮量对NSC浓度无显著影响;这说明不同植株体内NSC浓度在施氮条件下的变化是不同的。Scofield等[34]研究表明,小麦茎中在施氮处理下淀粉积累增多;剡斌等[35]的研究则表示施氮使胡麻茎中可溶性糖含量降低,淀粉含量升高,叶中可溶性糖含量随施氮量的增加先降低后升高,淀粉含量则先升高后减低,说明不同施氮量对植株NSC含量的影响不同。除草则普遍降低紫地榆和甘青大戟叶的非结构性碳水化合物含量,增加珠芽蓼的非结构性碳水化合物含量,这可能是因为除草使珠芽蓼的关照条件得到改善,光合作用增强,NSC含量上升。
随着未来全球变暖和氮素增加,巴郎山阳坡草甸将向以禾本科为优势种的方向发展,杂类草将受到抑制。以除草模拟的人类采药活动阻止亚高山草甸向以禾本科为优势种的方向发展,促进杂类草的优势度;但在温度升高和氮素增加的条件下,人类采药活动将进一步促进亚高山草甸向以禾本科为优势种的方向发展,并降低杂类草的优势度。
[1] ORESKES N. Beyond the ivory tower: the scientific consensus on climate change[J].Science, 2004, 306(5 702): 1 686-1 686.
[2] IPCC. Working group 1 contribution to IPCC fifth assessment report, climate change 2013: the physical science basis: summary for policymakers[R/OL].[2016-04-30].http://www.climatechange2013.org/images/report/WG1AR5_ALL_FINAL.pdf
[3] 赵艳艳, 周华坤, 姚步青, 等. 长期增温对高寒草甸植物群落和土壤养分的影响[J]. 草地学报, 2015, 23(4): 665-671.
ZHAO Y Y, ZHOU H K, YAO B J,etal. The influence of long-term simulating warming to the plant community and soil nutrient of alpine meadow[J].ActaAgrestiaSinica, 2015, 23(4): 665-671.
[4] 任 飞, 杨晓霞, 周华坤, 等.青藏高原高寒草甸3种植物对模拟增温的生理生化响应[J]. 西北植物学报, 2013, 33(11): 2 257-2 264.
REN F, YANG X X, ZHOU H K,etal. Physiological-biochemical responses of three plant species to experimental warming using OTC in alpine meadow on Qinghai-Tibetan Plateau[J].ActaBotanicaBoreali-OccidentaliaSinica, 2013, 33(11): 2 257-2 264.
[5] 石福孙, 吴 宁, 罗 鹏. 川西北亚高山草甸植物群落结构及生物量对温度升高的响应[J]. 生态学报, 2008, 28(11): 5 286-5 293.
SHI F S, WU N, LUO P. Effect of temperature enhancement on community structure and biomass of subalpine meadow in northwestern Sichuan[J].ActaEcologicaSinica, 2008, 28(11): 5 286-5 293.
[6] GRABHERR G, GOTTFRIED M, PAULI H. Climate effects of mountain plants[J].Nature, 1994, 369(6 480): 448-448.
[7] 李英年, 赵 亮, 赵新全, 等. 5年模拟增温后矮嵩草草甸群落结构及生产量的变化[J]. 草地学报, 2004, 12(3): 236-239.
LI Y N, ZHAO L, ZHAO X Q,etal. Effects of a 5-years mimic temperature increase to the structure and productivity ofKobresiahumilismeadow[J].ActaAgrestiaSinica, 2004, 12(3): 236-239.
[8] 徐满厚, 薛 娴. 青藏高原高寒草甸夏季植被特征及对模拟增温的短期响应[J]. 生态学报, 2013, 33(7): 2 071-2 083.
XU M H, XUE X. A research on summer vegetation characteristics & short-time responses to experimental warming of alpine meadow in the Qinghai-Tibetan Plateau[J].ActaEcologicaSinica, 2013, 33(7): 2 071-2 083.
[9] 刘 伟, 王长庭, 赵建中, 等. 矮嵩草草甸植物群落数量特征对模拟增温的响应[J]. 西北植物学报, 2010, 30(5):995-1 003.
LIU W, WANG C T, ZHAO J Z,etal. Responses of quantity characteristics of plant community to simulating warming in alpineKobresiahumilismeadow ecosystem[J].ActaBotanicaBoreali-OccidentaliaSinica, 2010, 30(5): 995-1 003.
[10] 樊 维, 蒙 荣, 陈全胜. 不同施氮水平对克氏针茅草原地上地下生物量分配的影响[J]. 畜牧与饲料科学, 2010, 31(2):74-76.
FAN W, MENG R, CHEN Q S. Effects of nitrogen additions on ground/underground biomass allocation ofStipakryloviicommunity[J].AnimalHusbandryandFeedScience, 2010, 31(2): 74-76.
[11] 黄忠良, 丁明懋, 张祝平, 等. 鼎湖山季风常绿阔叶林的水文学过程及其氮素动态[J]. 植物生态学报, 1994, 18(2): 194-199.
HUANG Z L, DING M M, ZHANG Z P,etal. The hydrological processes and nitrogen dynamics in a monsoon evergreen broad-leafed forest of Dinghu Shan[J].ActaPhytoecologicaSinica, 1994, 18(2):194-199.
[12] 张杰琦, 李 奇, 任正炜, 等. 氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响[J]. 植物生态学报, 2010, 34(10): 1 125-1 131.
ZHANG J Q, LI Q, REN Z W,etal. Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Plateau, China[J].ChineseJournalofPlantEcology, 2010, 34(10): 1 125-1 131.
[13] 高 嵩. 增温和氮素添加对松嫩草原羊草群落结构和功能的影响[D]. 长春: 东北师范大学, 2012.
[14] 喻理飞, 朱守谦, 叶镜中, 等. 人为干扰与喀斯特森林群落退化及评价研究[J]. 应用生态学报, 2002, 13(5): 529-532.
YU L F, ZHU S Q, YE J Z,etal. Evaluation on degradation of Karst forest community and human disturbance[J].ChineseJournalofAppliedEcology, 2002, 13(5): 529-532.
[15] 王 蓓, 孙 庚, 罗 鹏, 等. 模拟升温和放牧对高寒草甸土壤有机碳氮组分和微生物生物量的影响[J]. 生态学报, 2011, 31(6): 1 506-1 514.
WANG B, SUN G, LUO P,etal. Labile and recalcitrant carbon and nitrogen pools of an alpine meadow soil from the eastern Qinghai-Tibetan Plateau subjected to experimental warming and grazing[J].ActaEcologicaSinica, 2011, 31(6): 1 506-1 514.
[16] 仁青吉, 崔现亮, 赵彬彬. 放牧对高寒草甸植物群落结构及生产力的影响[J]. 草业学报, 2008, 17(6): 134-140.
REN Q J, CUI X L, ZHAO B B. Effects of grazing impact on community structure and productivity in an alpine meadow[J].ActaPrataculturaeSinica, 2008, 17(6): 134-140.
[17] 黎 洁. 西部贫困山区农户的采药行为分析——以西安周至县为例[J]. 资源科学, 2011, 33(6): 1 131-1 137.
LI J. On rural households’ behaviors of collecting Chinese herbs in poor mountainous areas in western China: evidence from Zhouzhi county, Xi’an City[J].ResourcesScience, 2011, 33(6): 1 131-1 137.
[18] PAULI H, GOTTFRIED M, GRABHERR G. High summits of the Alps in a changing climate[M]// “Fingerprints” of Climate Change. New York: Springer US, 2001: 139-149.
[19] ALWARD R D, DELTING J K, MILEHUNAS D G. Grassland vegetation changes and nocturnal global warming[J].Science, 1999, 283: 229-231.
[20] 赵建中, 彭 敏, 刘 伟, 等. 矮嵩草草甸不同功能群主要植物种生长特征与地表温度的相关性分析[J]. 西北植物学报, 2012, 32(3): 559-565.
ZHAO J Z, PENG M, LIU W,etal. Correlation between growing characters of main species belonging to different functional groups and ground temperature inKobresiameadow[J].ActaBotanicaBoreali-OccidentaliaSinica, 2012, 32(3): 559-565.
[21] 张法伟, 李跃清, 李英年, 等. 高寒草甸不同功能群植被盖度对模拟气候变化的短期响应[J]. 草业学报, 2010, 19(6): 72-78.
ZHANG F W, LI Y Q, LI Y N,etal. Short-term response of functional plant groups abundance to simulated climate change in alpine meadow ecosystems[J].ActaPrataculturaeSinica, 2010, 19(6): 72-78.
[22] 武建双, 沈振西, 张宪洲, 等. 藏北高原人工垂穗披碱草种群生物量分配对施氮处理的响应[J]. 草业学报, 2009,18(6): 113-121.
WU J S, SHEN Z X, ZHANG X Z,etal. Effect of nitrogen fertilizer application onElymusnutansbiomass allocation in an alpine meadow zone on the Tibetan Plateau[J].ActaPrataculturaeSinica, 2009, 18(6): 113-121.
[23] 曹文侠, 李 文, 李小龙,等. 施氮对高寒草甸草原植物群落和土壤养分的影响[J]. 中国沙漠, 2015, 35(3): 658-666.
CAO W X, LI W, LI X L,etal. Effects of nitrogen fertilization on plant community structure and soil nutrient in alpine meadow-steppe[J].JournalofDesertResearch, 2015, 35(3): 658-666.
[24] 郭红玉, 王 伟, 芦光新, 等. 增温施氮素对三江源区高寒草甸五种植物生长影响的初步研究[J]. 青海畜牧兽医杂志, 2015, 45(1): 26-29.
GUO H Y, WANG W, LU G X,etal. Impact of simulative warming and applying nitrogen on growth of five kinds of plants in alpine meadow of Sanjiangyuan region[J].ChineseQinghaiJournalofAnimalandVeterinarySciences, 2015, 45(1): 26-29.
[25] 贾 程. 巴郎山川滇高山栋NSC含量、TN含量及C/N对海拔梯度的响应[D]. 四川雅安: 四川农业大学, 2010.
[26] 史建伟, 张育平, 王孟本, 等. 植物体内非结构性碳水化合物变化及其影响因素[J]. 湖北农业科学, 2008, 47(1): 112-115.
SHI J W, ZHANG Y P, WANG M B,etal. Influential factors and change of non structural carbohydrates in plant[J].HubeiAgriculturalSciences, 2008, 47(1): 112-115.
[27] 李娜妮, 何念鹏, 于贵瑞. 中国4种典型森林中常见乔木叶片的非结构性碳水化合物研究[J]. 西北植物学报, 2015, 35(9): 1 846-1 854.
LI N N, HE N P, YU G R. Non-structural carbohydrates in leaves of tree species from four typical forests in China[J].ActaBotanicaBoreali-OccidentaliaSinica, 2015, 35(9): 1 846-1 854.
[28] HOCH G, KÖRNER C. The carbon charging of pines at the climatic treeline: a global comparison[J].Oecologia, 2003, 135(1): 10-21.
[29] GOUGH C M, FLOWER C E, VOGEL C S,etal. Phenological and temperature controls on the temporal non-structural carbohydrate dynamics ofPopulusgrandidentataandQuercusrubra[J].Forests, 2010, 1(1): 65-81.
[30] 毛子军, 贾桂梅, 刘林馨, 等. 温度增高、CO2浓度升高、施氮对蒙古栎幼苗非结构碳水化合物积累及其分配的综合影响[J]. 植物生态学报, 2010, 34(10): 1 174-1 184.
MAO Z J, JIA G M, LIU L X,etal. Combined effects of elevated temperature, elevated[CO2]and nitrogen supply on non-structural carbohydrate accumulation and allocation inQuercusmongolicaseedlings[J].ChineseJournalofPlantEcology, 2010, 34(10): 1 174-1 184.
[31] 蒋思思, 魏丽萍, 杨 松, 等. 不同种源油松幼苗的光合色素和非结构性碳水化合物对模拟氮沉降的短期响应[J]. 生态学报, 2015, 35(21): 7 061-7 070.
JIANG S S, WEI L P, YANG Y S,etal. Short term responses of photosynthetic pigments and nonstructural carbohydrates to simulated nitrogen deposition in three provenances ofPinustabulaeformisCarr seedlings[J].ActaEcologicaSinica, 2015, 35(21): 7 061-7 070.
[32] 王 雪, 雒文涛, 庾 强, 等. 半干旱典型草原养分添加对优势物种叶片氮磷及非结构性碳水化合物含量的影响[J]. 生态学杂志, 2014, 33(7): 1 795-1 802.
WANG X, LUO W T, YU Q,etal. Effects of nutrient addition on nitrogen, phosphorus and non-structural carbohydrates concentrations in leaves of dominant plant species in a semiarid steppe[J].ChineseJournalofEcology, 2014, 33(7): 1 795-1 802.
[33] 杨焕文, 耿宗泽, 李佛琳, 等. 不同施氮量的烤烟烟叶大田生长期碳水化合物的变化[J]. 云南农业大学学报, 2003,18(2): 153-157.
YANG H W, GENG Z Z, LI F L,etal. Changes of carbohydrate content in flue-cured tobacco leaves with different levels of nitrogen applied during growth phase[J].JournalofYunnanAgriculturalUniversity, 2003, 18(2): 153-157.
[34] SCOFIELD G N, RUUSKA S A, AOKI N,etal. Starch storage in the stems of wheat plants: localization and temporal changes[J].AnnalsofBotany, 2009, 103(6): 859-868.
[35] 剡 斌, 牛俊义, 崔政军, 等. 氮磷用量对胡麻非结构性碳水化合物积累转运及产量的影响[J]. 中国土壤与肥料, 2015,(2): 63-69.
YAN B, NIU J Y, CUI Z J,etal. Effect of nitrogen and phosphorus fertilization on accumulation, translocation of non-structural carbohydrate and yield of oil flax[J].SoilsandFertilizersSciencesinChina, 2015,(2): 63-69.
(编辑:潘新社)
Response of Plant Community in Subalpine Meadow to Climate Change
PANG Xiaoyu1, LEI Jingpin1*, WANG Ao2, DENG Yunpeng1
(1 Research Institute of Forestry,Key Laboratory of Forest Silviculture of the State Forestry Administration, Chinese Academy of Forestry,Beijing 100091,China;2 School of Nature Conservation, Beijing Forestry University, Beijing 100083, China)
A field experiment was conducted to understand the response of growth and non-structural carbohydrates (NSC) content of subalpine meadow plants to climate warming, nitrogen addition and human disturbance. We used open-top chamber (OTC) to simulate warming with nitrogen fertilization and weeding treatments, to analyze the growth and non-structural carbohydrates content of subalpine meadow plants in Balang Mountain, east of Qionglai Mountains in southeast margin of Tibetan Plateau (the transitional zone from Sichuan Basin to Tibetan Plateau) The results showed that: (1) the soil total phosphorus (P) and potassium (K) contents of all treatments have shown non-significant difference towards the control. Soil total nitrogen (N) content of warming+fertilization treatment has shown non-significant difference towards the control, while obviously decline of soil total nitrogen was shown in warming, fertilization, weeding, warming + weeding, fertilization + weeding, and warming + fertilization + weeding treatments. (2) Warming promoted the growth of functional group of grasses and forbs but inhibited the growth of functional group of sedges. Warming enhanced the importance value of functional group of grasses but reduced the importance value of functional group of forbs. And the effect of warming on functional group of sedges’ importance value was influenced by fertilization and weeding. Fertilization promoted the height growth of functional group of grasses and forbs, and promoted the growth of sedges. Weeding promoted the growth of functional group of sedges but weeding inhibited the growth of functional group of grasses and forbs. The interaction of fertilization and weeding was beneficial to functional group of grasses. Both fertilization and weeding enhanced functional group of sedges’ importance value but reduced functional group of grasses’ importance value. (3) Content and distribution of non-structural carbohydrates (NSC) of different species responded differently to the treatments. The non-structural carbohydrates content inGeraniumstrictipesshowed significant correlation with species coverage, and non-structural carbohydrates content inPolygonumviviparumshowed significant correlation with species height. The result indicates that both climate warming and nitrogen addition promote the growth of functional group of grasses and sedges, and cause the plants NSC changing to defend the environmental stresses.
subalpine meadow; experimental warming; nitrogen fertilization; non-structural carbohydrate(NSC).
1000-4025(2016)08-1678-09
10.7606/j.issn.1000-4025.2016.08.1678
2016-06-12;修改稿收到日期:2016-07-16
西部地区高山森林退化机制与恢复技术研究(CAFYBB2014ZD001)
庞晓瑜(1991-),女,在读硕士研究生,主要从事森林生态学研究。E-mail: 526774814@qq.com
*通信作者:雷静品,研究员,硕士生导师,主要从事森林生态学研究。E-mail: leijp@caf.ac.cn
Q948.79
A
