香菇多糖增强黄瓜幼苗对炭疽病菌抗性机理
2016-10-13张文华
张文华
(包头轻工职业技术学院,包头 014030)
香菇多糖增强黄瓜幼苗对炭疽病菌抗性机理
张文华
(包头轻工职业技术学院,包头 014030)
旨在考察香菇多糖诱导黄瓜(Cucumis sativus L. cv. Jinfeng)幼苗叶片细胞产生防御响应的效果。经0.75 g/L香菇多糖处理后,叶片胞内H2O2和水杨酸浓度迅速增加,分别在10 min和4 h达到峰值,随后均逐渐下降至诱导前水平。经香菇多糖诱导后,胞内抗性相关蛋白β-1,3-葡聚糖酶、几丁质酶和苯丙氨酸氨解酶比活性均显著增加,96 h后酶活力仍然保持较高水平。进一步研究发现,经香菇多糖诱导后,黄瓜对炭疽病菌(Colletotrichum orbiculare)抗性明显增强。经2.00 g/L香菇多糖处理后,叶片菌斑面积和数量分别显著下降至对照水平的53.1%和51.3%,结果表明香菇多糖具有诱导黄瓜细胞产生防御响应的作用。
香菇多糖;黄瓜;防御响应;炭疽病菌
黄瓜(Cucumis sativus L.)是一种世界范围内广泛种植的重要经济作物,但是其产量易受病原微生物如炭疽病菌(Colletotrichum orbiculare)的影响而下降。尽管化学杀菌剂可以有效缓解病害损失,但是农药的大量使用加重了社会和生态负担。
采用诱导剂激活植物细胞防御响应以抵抗病原微生物侵染是一种较为理想的方法[1]。细胞防御响应包含一系列的级联过程,包括活性氧粒子的积累、信号分子浓度增加、苯丙素途径强化以及病程相关蛋白的生成[2,3]。病程相关蛋白包含多种蛋白酶抑制剂及水解酶作用于外源微生物而起到防御作用。苯丙素类物质除激活信号传导以外,还能够改变植物细胞壁结构以起到屏障作用[4]。
研究表明,β-葡聚糖能够激活多种植物的防御响应。如昆布多糖和热凝胶寡糖分别被证实能够对烟草和马铃薯产生诱导作用[5,6]。香菇多糖(Len-tinan,Ltn)是一种线性β-1,3-葡聚糖,具有潜在的激活植物防御响应的活性,但是其对黄瓜细胞是否具有诱导活性仍需要进一步研究。综上所述,本研究通过对黄瓜叶片细胞内信号分子浓度以及关键酶活性的测定,结合黄瓜对炭疽病菌抗性效果统计结果,对香菇多糖诱导黄瓜细胞防御响应的效果进行考察,旨在为黄瓜病害的生态防治提供数据支持。
1 材料与方法
1.1 材料
1.1.1 黄瓜和菌种 黄瓜植株(C. sativus L. cv. Jinfeng)于无菌温室中培养(培养温度25℃,光照周期16 h/d)。炭疽病菌(C. orbiculare)购自广东菌种保藏中心(广州),菌种培养及孢子收集参照Zhang等[7]的方法。
1.1.2 试剂 香菇多糖,蓝色几丁质(Chitin azure),反式肉桂酸,水杨酸购自Sigma公司(美国);其余试剂均购自国药集团化学试剂有限公司(上海)。
1.1.3 仪器 液相色谱仪Hitachi CM5000 HPLC system(日本);离心机Sigma 2K5S(美国)。
1.2 方法
1.2.1 不同浓度香菇多糖对炭疽病菌侵染黄瓜幼苗的抑制 分别以0.25、0.75和2.00 g/L香菇多糖溶液均匀喷撒黄瓜幼苗叶片表面,以无菌去离子水作为空白对照(CK)。喷洒24 h后,以浓度为107孢子/mL C. orbiculare孢子悬液均匀喷洒叶片表面。培养168 h后对叶片病斑数量和病斑面积进行统计。每种处理采用10株马铃薯幼苗,所有植株培养条件一致。
1.2.2 香菇多糖对黄瓜的诱抗作用机理
1.2.2.1 黄瓜叶片处理与取样 以0.75 g/L香菇多糖溶液喷撒培养6周的黄瓜幼苗叶片表面,以无菌去离子水作为空白对照(CK)。然后,分别于喷洒后剪取叶片用于胞内H2O2和水杨酸(salicylic acid,SA)浓度测定或胞内β-1,3-葡聚糖酶、几丁质酶和苯丙氨酸氨解酶(phenylalanine ammonia lyase,PAL)比酶活测定。每项指标测定均采用不同植株的叶片进行3次重复。
1.2.2.2 各种诱抗物质测定方法 (1)叶片胞内H2O2和水杨酸浓度测定:H2O2与SA的提取及浓度测定分别采用Li等[8]和van Spronsen等[9]报道的方法。(2)胞内蛋白提取方法:称取200 mg黄瓜叶片并迅速于液氮中研磨成粉末,随后加入1.0 mL蛋白提取溶液(含有50 mmol/L乙酸钠缓冲液,5 mmol/L EDTA,2 mmol/L DTT,0.04% 硫代硫酸钠,0.1%聚乙烯吡咯烷酮,pH5.5)并于4℃震荡溶解10 min。然后将混合液于4℃,10 000×g离心30 min并收集上清,于-20℃保存。(3)β-1,3-葡聚糖酶活力测定:测定反应体系包含200 μL 10 g/L昆布多糖溶液(溶于50 mmol/L乙酸钠缓冲液,pH5.0),200 μL 50 mmol/L乙酸钠缓冲液(pH5.0)和100 μL蛋白提取液。该反应体系于37℃反应2 h,随后沸水浴10 min终止反应。反应体系中加入预先灭活的蛋白提取液作为空白对照。反应体系中每1 min生成1 μg还原糖的酶量定义为1酶活单位(1 U)。(4)几丁质酶活力测定:几丁质酶活力测定采用Ippolito等[10]报道的方法。反应体系包含140 μL 25 g/L蓝色几丁质溶液(溶于50 mmol/L乙酸钠缓冲液,pH5.0)和70 μL蛋白提取液。该反应体系于37℃反应2 h,随后加入100 μL 2 mol/L HCl终止反应。(5)苯丙氨酸氨解酶活力测定:PAL活力测定采用Wang等[11]报道的方法。反应体系包含500 μL 20 mmol/L L-苯丙氨酸(溶于50 mmol/L Tris-HCl缓冲液,pH8.5),400 μL 50 mmol/L Tris-HCl缓冲液(pH8.5)和100 μL蛋白提取液。该反应体系于37℃反应2 h,随后加入200 μL 6 mol/L HCl终止反应。
1.2.3 统计方法 数据统计采用SPSS统计软件(V-er. 18.0,SPSS Inc.,美国)。单因素方差分析(one-way ANOVA)用于各测定指标不同处理组之间的比较。最小显著差数法(least significant difference,LSD)用于测定指标间显著性分析(显著性水平P<0.05)。
2 结果
2.1 香菇多糖提高黄瓜对炭疽病菌的抗性
三种浓度的Ltn溶液处理黄瓜叶片后对叶片上炭疽病菌侵染形成的菌斑数量和菌斑面积进行统计,结果表明经Ltn处理后黄瓜植株对炭疽病抗性明显增加,并且抗性效果随着Ltn浓度增加而逐渐增强。Ltn浓度达到0.75 g/L后,叶片感染面积和病变数量与对照组相比均显著性下降,而Ltn浓度提高至2.00 g/L时,与对照组相比抗性效力接近50%,表现出较为理想的效果。

表1 香菇多糖诱导黄瓜叶片对炭疽病的抗性效果
2.2 香菇多糖浓度对诱抗效果的影响
采用Ltn处理黄瓜叶片24 h后,胞内SA浓度随着Ltn浓度提高而迅速增加(图1)。采用0.75 g/L Ltn处理叶片后,胞内SA达到1.785 μg/g,而继续提高Ltn浓度,胞内SA水平保持稳定。结果表明,Ltn浓度0.75 g/L是达到稳定诱导效果的临界水平,后续实验均在此水平条件下进行。

图1 香菇多糖处理黄瓜叶片24 h后胞内水杨酸浓度变化
2.3 香菇多糖提高黄瓜胞内信号分子浓度
测定经Ltn诱导后黄瓜叶片细胞内H2O2和SA浓度变化趋势。结果(图2)显示,经Ltn诱导后胞内H2O2浓度急剧升高,至10 min后达到峰值,随后迅速下降。尽管诱导30 min后H2O2浓度仍高于诱导前水平但是两者之间无显著性差异。60 min后胞内H2O2浓度恢复至诱导前水平。胞内SA浓度变化趋势与H2O2类似。胞内SA水平迅速增加至诱导4 h后达到峰值,随后逐渐下降至120 h后恢复至诱导前水平。

图2 香菇多糖处理后黄瓜叶片胞内H2O2(A)和水杨酸(B)浓度变化曲线
2.4 香菇寡糖增强黄瓜胞内抗性蛋白比活性
由图3可知,与空白对照(CK)相比,经0.75 g/L Ltn处理24 h后胞内β-1,3-葡聚糖酶比酶活显著增加至115 U/mg,诱导96 h后酶活下降至89 U/mg,但是仍显著高于CK水平。另一方面,经炭疽病菌(CO)处理后,胞内β-1,3-葡聚糖酶的酶活水平同样出现了显著上升,但低于Ltn处理组的水平。经Ltn和CO联合处理(Ltn+CO)的叶片胞内酶活与Ltn组处于同一水平。结果表明,Ltn处理能够模拟炭疽病菌的侵染作用而提高胞内β-1,3-葡聚糖酶的比酶活,且在96 h后仍然处于较高水平。测定表明,Ltn、CO和Ltn+CO组胞内几丁质酶活性相比于对照组均有了显著增加,且3种处理方式间并无显著性差异。该结果与β-1,3-葡聚糖酶测定结果一致,即Ltn能够诱导胞内病程相关蛋白的表达。分别经Ltn和CO处理后,黄瓜叶片胞内PAL活性均产生了显著的增加,且96 h后仍然保持较高活性。该结果与β-1,3-葡聚糖酶和几丁质酶测定结果一致。
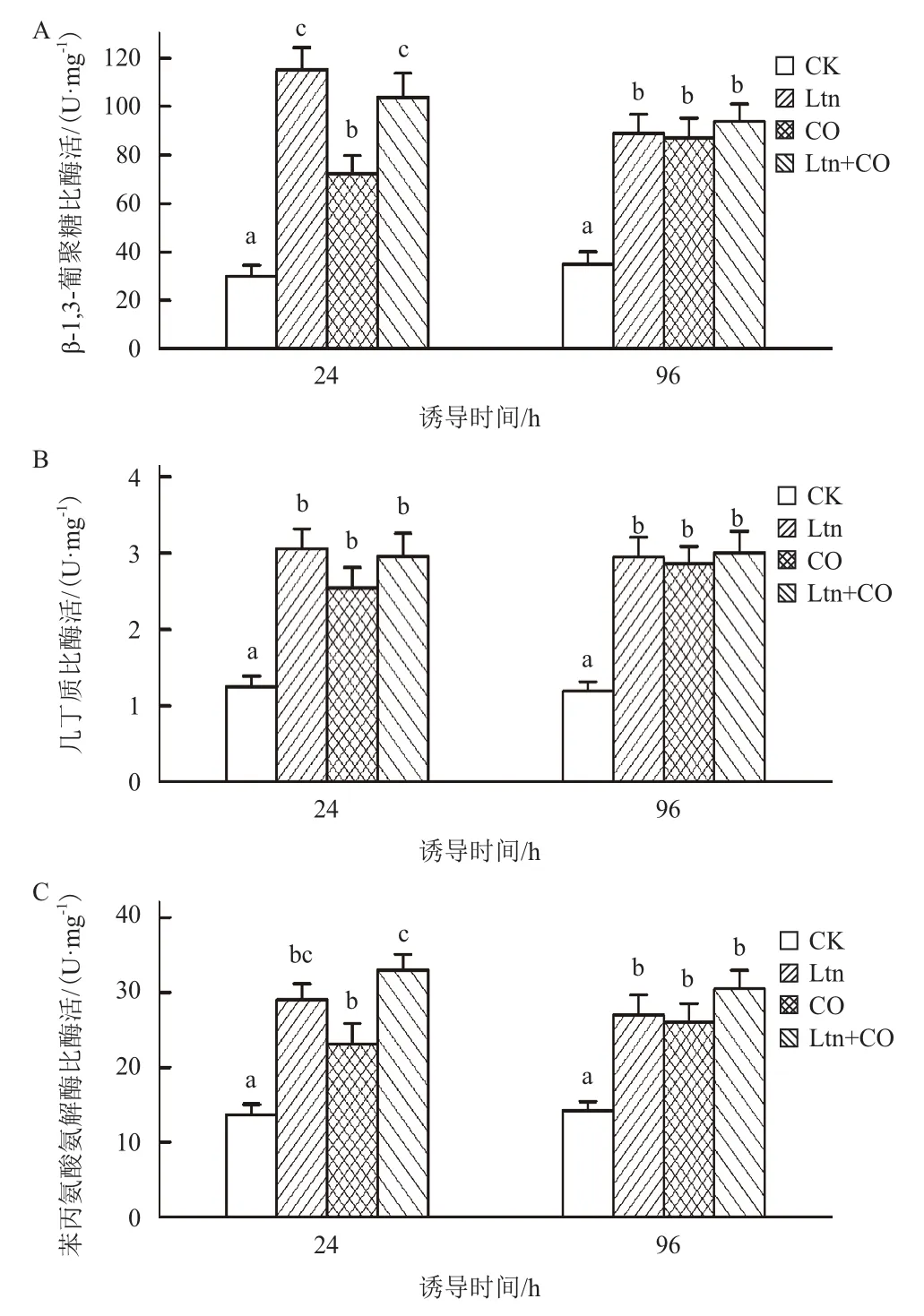
图3 处理方式对黄瓜叶片胞内β-1,3-葡聚糖酶(A)、几丁质酶(B)和苯丙氨酸氨解酶(C)活力测定影响
3 讨论
水杨酸(SA)作为胞内信号分子在植物组织建立系统性的防御响应过程中起到关键作用[12,13]。因此,以胞内SA积累浓度作为评价指标考察了香菇多糖(Ltn)浓度对黄瓜叶片组织诱抗效果的影响。植物细胞膜表面受体识别到诱导子后迅速激活氧代谢爆发产生活性氧中间体(ROI),如O2-和H2O2,并进一步激活依赖ROI的防御响应二级信使水杨酸和防御相关基因的表达以及抗菌代谢产物的合成[3,14]。因此,胞内信号分子H2O2和SA积累水平产生显著变化是确定细胞防御响应发生的关键指标。另一方面,胞内一级和二级信号分子H2O2和SA,浓度分别在10 min和4 h时达到最高水平,表明胞内信号放大存在一定的时间滞后效应,该现象与前述研究结果一致[5,15]。胞内SA浓度在4-96 h均显著高于诱导前水平,表明Ltn处理能够在较长时间内对黄瓜叶片细胞的防御响应产生诱导作用。
为进一步验证Ltn对黄瓜细胞防御抗性的诱导作用,本研究考察了经Ltn处理后胞内3种与防御相关的典型蛋白的酶活性的变化情况。葡聚糖酶因能够水解致病性真菌细胞壁的主要组分β-1,3-葡聚糖而具有较高的抗真菌活性,属于病程相关蛋白PR-2家族[16]。同时,β-1,3-葡聚糖水解后的寡糖能够进一步作为诱导子强化植物的防御响应[17]。几丁质酶属于病程相关蛋白PR-3、-4、-8和-11家族[16],作用于病原菌细胞壁以保护植物自身组织[18]。苯丙氨酸氨解酶(PAL)催化L-苯丙氨酸脱氨基生成反式-肉桂酸,是苯丙素途径的关键酶[19]。而苯丙素途径的产物和中间物除作为物理和化学屏障以防御病原菌侵染外,还能够激活局部信号通路以强化诱导基因的表达[4]。因此,PAL活性是评价植物防御响应的重要指标。分析表明,香菇多糖处理能够显著激活黄瓜胞内抗性相关蛋白的活性。
测定结果表明,Ltn处理能够显著提高黄瓜叶片胞内信号分子浓度,同时对细胞防御响应相关蛋白的表达产生显著促进作用。采用Ltn处理黄瓜叶片后,胞内防御相关蛋白的表达与经炭疽病菌处理组具有相同水平。由此可知,预先采用Ltn处理黄瓜叶片具有增强植物防御炭疽病菌侵染的潜在作用。而炭疽病菌侵染实验表明,经Ltn处理后叶片感染面积和病变数量与对照组相比均显著性下降。
4 结论
香菇多糖Ltn处理能够显著提高黄瓜叶片胞内信号分子浓度,对细胞防御响应产生激活作用,能够显著激活黄瓜胞内抗性相关蛋白的活性,并能显著降低叶片被病原菌感染面积和病变数量。由上可知,香菇多糖预处理明显提高黄瓜对炭疽病菌的抗性。作为一种环境安全性的生物多糖,香菇多糖具有发展成环境友好型生物农药的潜力,进一步拓展了其应用范围。
[1]Jones JDG, Dangl JL. The plant immune system[J]. Nature, 2006,444(7117):323-329.
[2]Kombrink E, Somssich IE. Defense responses of plants to pathogens[M]// Callow JHA, Inez CT, editor. Advances in Botanical Research:Academic Press, 1995:1-34.
[3]Mcdowell JM, Dangl JL. Signal transduction in the plant immune response[J]. Trends in Biochemical Sciences, 2000, 25(2):79-82.
[4]Dixon RA, Achnine L, Kota P, et al. The phenylpropanoid pathway and plant defence-a genomics perspective[J]. Molecular Plant Pathology, 2002, 3(5):371-390.
[5]Klarzynski O, Plesse B, Joubert JM, et al. Linear β-1, 3 glucans are elicitors of defense responses in tobacco[J]. Plant Physiology,2000, 124(3):1027-1037.
[6] Li J, Zhu L, Lu G, et al. Curdlan β-1, 3-Glucooligosaccharides induce the defense responses against Phytophthora infestans infection of potato(Solanum tuberosum L. cv. McCain G1)leaf cells[J]. PLoS One, 2014, 9(5):e97197.
[7] Zhang PY, Wang JC, Liu SH, et al. A novel burdock fructooligosaccharide induces changes in the production of salicylates, activates defence enzymes and induces systematic acquired resistance to Colletotrichum orbiculare in cucumber seedlings[J]. Journal of Phytopathology, 2009, 157:201-207.
[8]Li SJ, Zhu TH. Biochemical response and induced resistance against anthracnose(Colletotrichum camelliae)of camellia(Camellia pitardii)by chitosan oligosaccharide application[J]. Forest Pathology, 2013, 43:67-76.
[9] van Spronsen PC, Tak T, Rood AM, et al. Salicylic acid inhibits indeterminate-type nodulation but not determinate-type nodulation[J]. Molecular Plant-Microbe Interactions, 2003, 16(1):83-91.
[10]Ippolito A, El Ghaouth A, Wilson CL, et al. Control of postharvest decay of apple fruit by Aureobasidium pullulans and induction of defense responses[J]. Postharvest Biology and Technology,2000, 19(3):265-272.
[11]Wang JW, Wu JY. Involvement of nitric oxide in elicitor-induced defense responses and secondary metabolism of Taxus chinensis cells[J]. Nitric Oxide-Biology and Chemistry, 2004, 11(4):298-306.
[12]Delaney TP, Uknes S, Vernooij B, et al. A central role of salicylic acid in plant disease resistance[J]. Science, 1994, 266(5188):1247-1250.
[13] Zhang ZG, Wang YC, Ji R, et al. The role of SA in the hypersensitive response and systemic acquired resistance induced by elicitor PB90 from Phytophthora boehmeriae[J]. Physiological and Molecular Plant Pathology, 2004, 65(1):31-38.
[14] Jones JD, Dangl JL. The plant immune system[J]. Nature, 2006,444(7117):323-329.
[15]El Modafar C, Elgadda M, El Boutachfaiti R, et al. Induction of natural defence accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to bioelicitors isolated from green algae[J]. Scientia Horticulturae, 2012, 138:55-63.
[16]Van Loon L, Van Strien E. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins[J]. Physiological and Molecular Plant Pathology, 1999,55(2):85-97.
[17]Vigers AJ, Roberts W, Selitrennikoff CP. A new family of plant antifungal proteins[J]. Molecular Plant-Microbe Interactions,1991, 4(4):315-323.
[18]Kuć J. Molecular aspects of plant responses to pathogens[J]. Acta Physiologiae Plantarum, 1997, 19(4):551-559.
[19]Matsuda F, Morino K, Ano R, et al. Metabolic flux analysis of the phenylpropanoid pathway in elicitor-treated potato tuber tissue[J]. Plant and Cell Physiology, 2005, 46(3):454-466.
(责任编辑 马鑫)
The Mechanism of Lentinan Enhancing the Resistance of Cucumber Seedlings to Colletotrichum orbiculare
ZHANG Wen-hua
(Baotou Light Industry Vocational Technical College,Baotou 014030)
This study is to investigate the inducing effect of lentinan on the defense responses of cucumber(Cucumis sativus L. cv. Jinfeng)seedling leaves. The seedlings were treated with 0.75 g/L of lentinan. The concentration of H2O2and salicylic acid in cells of leaves increased rapidly and reached the peak levels at 10 min and 4 h,respectively. Afterwards,the levels of both H2O2and salicylic acid gradually declined to the ones before treatment. Moreover,the activities of defense related proteins,including β-1,3-glucanase,chitinase,and phenylalanine ammonia lyase,significantly increased after lentinan induction,and still kept stable after 96 hours. In addition,the resistance of cucumber treated with lentinan to Colletotrichum orbiculare was notably enhanced after lentinan induction. Both the number and area of lesions on cucumber leaves significantly decreased to 53.1% and 51.3% in comparison with the control,respectively,which indicated that lentinan played a role in inducing the defense responses of cucumber cells.
lentinan;Cucumis sativus L. cv. Jinfeng;defense response;Colletotrichum orbiculare
10.13560/j.cnki.biotech.bull.1985.2016.03.015
2015-06-09
张文华,女,讲师,研究方向:微生物技术与应用;E-mail:1072418227@qq.com
