阿尔巴斯绒山羊骨髓间充质干细胞体外分离培养及鉴定
2016-10-13马苏日古嘎彭航刘佳佳张亦婷桂花刘鹏霞
马苏日古嘎彭航刘佳佳张亦婷桂花刘鹏霞
(1.内蒙古大学生命科学学院,呼和浩特 010021;2.西安交通大学前沿科学技术研究院 骨骼关节疾病与治疗研究中心,西安 710061;3.中国科学院微生物研究所,北京100101;4.中国科学院上海药物研究所 药物安全性评价中心,上海 201203)
阿尔巴斯绒山羊骨髓间充质干细胞体外分离培养及鉴定
马苏日古嘎1彭航2刘佳佳3张亦婷4桂花1刘鹏霞1
(1.内蒙古大学生命科学学院,呼和浩特 010021;2.西安交通大学前沿科学技术研究院 骨骼关节疾病与治疗研究中心,西安 710061;3.中国科学院微生物研究所,北京100101;4.中国科学院上海药物研究所 药物安全性评价中心,上海 201203)
探讨阿尔巴斯绒山羊骨髓间充质干细胞(Arbas cashmere goat bone marrow-derived mesenchymal stem cells,gBMMSCs)体外分离培养,并对其生物学特性进行系统的鉴定,进一步深入分析体外诱导时多向分化相关基因动态表达情况。结果显示,原代培养的gBM-MSCs呈梭形或纺锤形、放射状排列的细胞集落,能够连续稳定传代至15代以上;免疫荧光及流式细胞术检测显示,gBM-MSCs表达间质系特异性表面标志物,如:CD13、CD29、CD44、CD106、CD90及CD166,不表达造血干细胞表面标志物CD45及CD34;体外诱导实验证实该细胞具有多向分化潜能,能够向脂肪细胞、成骨细胞和软骨细胞分化;荧光定量PCR实验结果显示,gBM-MSCs多向分化相关基因的表达水平随着诱导天数的增加呈上升趋势。实验结果证实阿尔巴斯绒山羊骨髓中分离培养的细胞具备gBM-MSCs的生物学特性。
阿尔巴斯绒山羊;间充质干细胞;培养;分化;生物学特性
1966年,Friedenstein等[1-3]首次从大鼠骨髓中分离鉴定出具有造血支持功能的干细胞,并证明其在体外可以分化为骨细胞及脂肪细胞。在此基础上,Owen及其同事提出间充质干细胞(Mesenchymalstem cells,MSCs)这一概念[4,5]。目前,已经证实MSCs是中胚层来源的一类多能干细胞,具有自我更新及多向分化潜能,广泛存在于胎儿和成体多种组织器官中,例如:骨髓、脂肪、羊水、骨骼肌、胎肺、胎肝、脐血、脐带和胎盘等[6-11]。其中,骨髓中的含量高,分离及获取操作简便,可于自身获取等特点一直深受研究人员的青睐,因此骨髓来源的MSCs研究比较普遍;但是,目前绒山羊骨髓MSCs体外分离培养的文献报道极少。
绒山羊作为经济价值很高的家畜,有关其生物学研究对当代畜牧业发展具有重大的战略意义。目前的绒山羊转基因克隆大多使用的是成纤维细胞,它的一个显著的缺点是培养传代数目有限、基因转染后存活率低,这一缺点已成为绒山羊转基因新品种培育的瓶颈。本研究从阿尔巴斯绒山羊骨髓中分离培养MSCs,并对其生物学特性进行系统研究,旨在深入了解大型偶蹄类动物骨髓MSCs的生物学特性,为后续转基因动物新品种培育及组织工程提供参考。
1 材料与方法
1.1 材料
1.1.1 实验动物 阿尔巴斯绒山羊3-5月龄流产胎儿5个。
1.1.2 主要试剂 DMEM/F12、IMDM、胎牛血清(Hyclone公司产品);胰酶、L-谷氨酰胺、地塞米松、吲哚美辛、β-磷酸甘油、维生素C、黄嘌呤、Giemsa染液、DAPI染液(Sigma公司产品);油红O(上海紫一试剂厂);转化生长因子-β、人碱性成纤维细胞生长因子、人表皮生长因子(Pepro Tech公司产品);阿利新蓝染液(阿拉丁公司产品);氨水(永大公司产品);CCK-8试剂盒(Beyotime公司产品);TRLzol 裂解液(Ambion公司产品);反转录试剂盒(TaKaRa公司产品);兔多克隆抗体 CD13、CD29、CD34、CD44、CD45、CD90、CD106和CD166(博士德产品);山羊抗兔 FITC-IgG(Protein Tech产品);定量PCR引物均由大连宝生物(TaKaRa)公司设计并合成,序列见表1。

表1 实时定量PCR引物序列
1.2 方法
1.2.1 细胞分离和培养 无菌条件下取阿尔巴斯绒山羊3-5月龄流产胎儿腿骨,用培养基冲洗获取骨髓,加入肝素抗凝。置20 mL DMEM/F12培养基中,1 000 r/min离心10 min,收集细胞;再加入完全培养基(含有DMEM/F12,50 ng/mL人表皮生长因子,10 ng/mL人碱性成纤维细胞生长因子,10-8mol/L地塞米松,2 mmol/L谷氨酰胺,5%胎牛血清,100 U/mL青霉素/链霉素),将细胞悬浮后接种到10 mm培养皿中,随后置于37℃、5% CO2、饱和湿度培养箱中培养,48 h后全量换液,以后每隔2 d换液,当细胞生长至80%融合时用0.25%胰蛋白酶进行1∶2的消化传代。培养的过程中,倒置相差显微镜下观察细胞形态和生长状况。
1.2.2 细胞增殖能力检测 取分离至3个不同个体的处于对数生长期的第4代gBM-MSCs,加入完全培养基稀释为2×104/mL细胞悬液,按每孔100 μL接种到96孔板中,每隔2 d换液;接种后的第1天起,每隔24 h加入10 μL CCK-8试剂,置于37℃恒温箱中孵育4 h,用酶标仪检测450 nm处的吸光值,连续检测7 d,并用GraphPad Prism 5软件进行数据处理。
1.2.3 细胞核型分析 取第15代处于对数生长期的gBM-MSCs,加入80 μL 0.1 mg/mL秋水仙素,置37℃恒温培养箱中孵育3.5 h;消化收集细胞,加入8 mL 0.075 mol/L KCl低渗液,置于37℃水浴锅,孵育30 min,固定,滴片;Gimesa染色15 min,自来水冲洗,自然晾干,油镜下观察细胞中期染色体形态及数目,并用Genus软件分析。
1.2.4 免疫荧光法检测细胞表面标志物CD13、CD-29、CD44、CD90和CD106 取第4代gBM-MSCs,在6孔板内进行细胞爬片,细胞生长至80%融合时,每孔加入1.5 mL 4%多聚甲醛固定30 min;加入封闭液(PBS+2%牛血清蛋白+2%山羊封闭血清+2%脱脂奶粉+0.15 mol/L甘氨酸)室温封闭2 h;滴加1∶200稀释兔多克隆抗体CD13、CD29、CD44、CD106、CD90,4℃孵育过夜,PBS冲洗;之后加入1∶50稀释异硫氰酸荧光素(FITC)标记的山羊抗兔IgG,室温孵育2 h;DAPI染色15 min后共聚焦显微镜下观察结果。阴性对照以磷酸盐缓冲液(PBS)代替第一抗体。
1.2.5 流式细胞术检测细胞表面标志物CD34、CD45和CD166 消化收集第4代gBM-MSCs,用F-PBS(含1%胎牛血清PBS)重悬,将细胞以106个/管分装至2 mL离心管中,离心弃上清,分别加5 μL兔多克隆抗体 CD34、CD45和CD166,避光常温孵育2 h,PBS冲洗,之后加入1∶50稀释异硫氰酸荧光素(FITC)标记的山羊抗兔IgG,室温孵育2 h;补加F-PBS冲洗2次,离心弃上清,加800 μL 2%多聚甲醛固定,流式细胞仪检测,用FlowJo 7.6.1软件分析数据。
1.2.6 多向分化潜能检测 消化收集第4代gBMMSCs,加完全培养基,将细胞浓度调整至2×104个/mL,接种于六孔板中,培养24 h后,更换诱导培养液,以后每隔2 d换液,第21天检测诱导结果。对于成脂肪诱导的细胞,用1%油红O染色,倒置荧光显微镜观察脂滴形成情况;用RT-PCR法检测脂肪细胞经典基因过氧化物酶体增殖物激活受体γ PPARG(Peroxisome proliferator-activated receptor gamma)的表达。成骨诱导的细胞,用20 ng/mL硝酸银染色,观察钙盐沉积情况;用RT-PCR法检测骨细胞经典基因骨钙素蛋白GLA(Galactosidase alpha)的表达。成软骨诱导的细胞,用1%阿利新蓝染色,观察酸性黏多糖合成情况,用RT-PCR法检测软骨细胞经典基因光蛋白聚糖LUM(lumican)的表达。
成脂肪诱导液:IMDM,5%牛胎血清,1 μmol/L地塞米松,5 μg/mL胰岛素,0.5 mmol/L黄嘌呤,60 μmol/L吲哚美辛;成骨诱导液:IMDM,5%牛胎血清,0.1 μmol/L地塞米松,0.05 mmol/L维生素C,10 mmol/L β-甘油磷酸;成软骨诱导液:IMDM,5%牛胎血清,10 ng/mL 转化生长因子-β,10-7mol/L地塞米松,50 μg/mL维生素C。
1.2.7 多向分化相关基因动态表达分析 按照上述1.2.6中的实验方法诱导培养gBM-MSCs后,分别提取诱导7、14和21 d 后的细胞总RNA,通过荧光定量PCR法检测诱导不同天数后相应分化基因动态表达情况。对于成脂肪诱导的细胞,检测脂肪经典基因PPARG的表达趋势;对于成骨诱导的细胞,检测骨经典基因GLA的表达趋势;对于成软骨诱导的细胞,检测软骨经典基因LUM的表达趋势;以未诱导的gBM-MSCs作为对照,基因相对表达量用GAPDH标准化并用 2-ΔΔCt法运算。
2 结果
2.1 细胞分离和培养
实验对5个绒山羊流产胎儿进行了骨髓分离培养,获得了5株gBM-MSCs。gBM-MSCs原代培养时4-6 h开始贴壁,大约培养14 d后达到80% 汇合,通过培养基筛选和传代培养去除其他类型的干细胞和终末分化细胞,传至P5时呈现均一的成纤维样细胞,贴壁生长,状态良好(图1)。
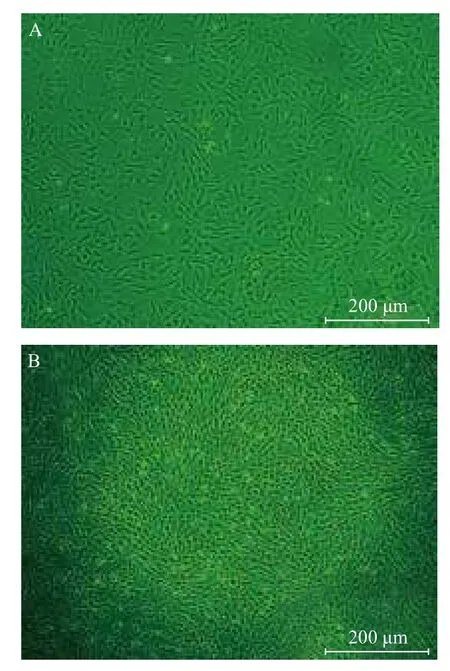
图1 阿尔巴斯绒山羊骨髓间充质干细胞形态
2.2 细胞增殖能力检测
第5代gBM-MSCs生长曲线呈S型(图2),接种后第0-1天为潜伏适应期,从第1天起快速增殖,进入对数生长期,第5天进入平台期,平台期持续1-2 d,之后细胞增殖速度显著下降,进入衰退期。
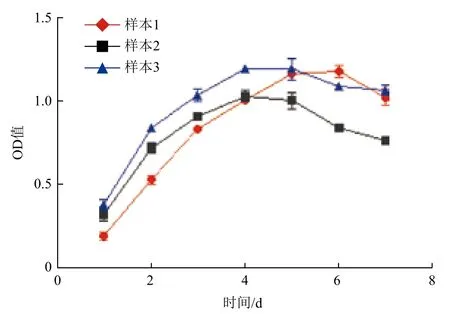
图2 第5代阿尔巴斯绒山羊骨髓间充质干细胞生长曲线
2.3 细胞核型分析
为验证本实验所分离出的gBM-MSCs在体外传代过程中细胞染色体数目是否完整,我们收集第15代gBM-MSCs做染色体数目分析,随机选取20个样本进行染色体计数。结果(图3)显示,所做的第15代gBM-MSCs中有95%(19/20)的细胞染色体为2n=60条,具有正常二倍体核型,表明体外培养的gBM-MSCs能够进行正常染色体复制和分裂。
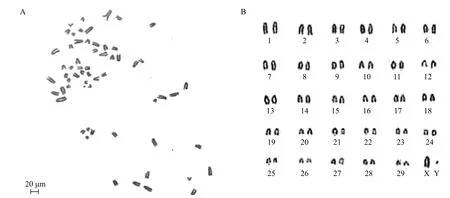
图3 第15代阿尔巴斯绒山羊骨髓间充质干细胞核型(2n=60)
2.4 免疫荧光法检测细胞表面标志物CD13、CD29、CD44、CD90和CD106
免疫荧光检测结果显示,第5代gBM-MSCs表达间质系表面标志物CD13(图4-B)、整合素家族成员CD29(图4-C)、黏附分子CD44(图4-D)、CD106(图4-E)及CD90(图4-F)。
2.5 流式细胞术检测细胞表面标志物CD34、CD45和CD166
流式细胞术检测结果显示,第5代gBM-MSCs表达粘附分子CD166(图5-A)、不表达造血干细胞表面标志物CD34(图5-B)、CD45(图5-C)。
2.6 多向分化潜能检测
第5代gBM-MSCs分别用成脂肪、成骨和成软骨培养基诱导21 d后,进行染色和RT-PCR法鉴定。结果(图6)显示,成脂肪诱导后细胞质中形成脂肪颗粒,经油红O染色后,能观察到桔红色的脂滴(图6-A1),RT-PCR显示成脂肪诱导后的细胞阳性表达脂肪经典基因PPARG(图6-A3);成骨诱导后细胞质中出现钙盐沉积,硝酸银染色呈阳性(图6B1),RT-PCR显示成骨诱导后的细胞阳性表达成骨经典基因GLA(图6-B3);成软骨诱导后细胞质中形成酸性黏多糖,经阿尔新兰染色后,能特异性着蓝色(图6-C1);RT-PCR显示成软骨诱导后的细胞阳性表达软骨经典基因LUM(图6-C3)。以上结果显示,gBM-MSCs具有多向分化潜能。

图4 免疫荧光法检测第5代阿尔巴斯绒山羊骨髓间充质干细胞表面标志物
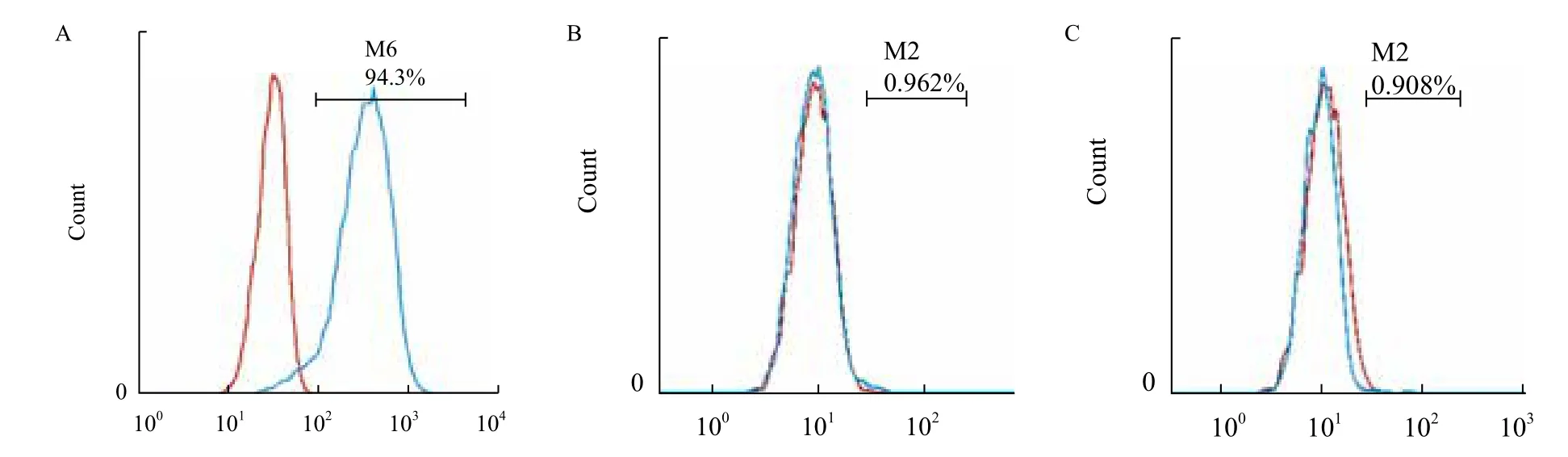
图5 流式细胞术检测第5代阿尔巴斯绒山羊骨髓间充质干细胞表面标志物
2.7 多向分化相关基因动态表达分析
进一步通过荧光定量PCR法检测诱导7、14和21 d后相关分化基因动态表达情况,结果(图7)显示,成脂肪细胞诱导后脂肪经典基因PPARG表达量随着诱导天数的增加逐步上升,诱导21 d后达到最高,约上调1 270倍(图7-A);成骨细胞诱导后骨细胞经典基因GLA的表达也随着诱导天数的增加逐步上调,诱导21 d后达到最高,约上调165倍(图7-B);成软骨细胞诱导后软骨经典基因LUM的表达量也逐步增加,诱导21 d后达到最高,约上调63倍(图7-C)。本实验进一步证实我们分离培养的gBM-MSCs具有多向分化潜能,多向分化相关基因的表达水平随着诱导天数的增加呈上升趋势。
3 讨论
本研究采用全骨髓贴壁培养法,对5个绒山羊流产胎儿进行了骨髓分离培养,用含人表皮生长因子(EGF)和人碱性成纤维细胞生长因子(bFGF)等的扩增培养基培养,获得了5株细胞,该细胞具备MSCs的生物学特征。有研究证实,培养基中添加人碱性成纤维细胞生长因子有利于细胞贴壁生长[12],同时不影响其多向分化能力[13];而人表皮生长因子的添加能够促进细胞增殖,缩短细胞的倍增时间[14,15]。
目前,MSCs的鉴定采用2006年国际细胞治疗协会(International Soeiety for Cellular Therapy,ISCT)提出的标准,包括以下几方面:①细胞形态为成纤维细胞样,具有贴壁生长特性;②阳性表达间质系表面标志物,如CD73(SH3和SH4),CD105(SH2)和CD90(Thy1)等,阴性表达造血干细胞表面标志物,如:CD45,CD19,CD19或CD79、CD14或CD11b和HLA-DR等;③具有向脂肪细胞、成骨细胞和软骨细胞分化的多向分化潜能[16,17]。本实验对gBM-MSCs生物学特征进行了系统研究,结果显示,gBM-MSCs体外培养时具有贴壁特性,呈梭形或纺锤形,具有较强的增殖能力,符合MSCs的基本特征;可连续稳定传代至15代以上,第15代gBMMSCs仍然具备正常二倍体核型;第5代gBM-MSCs表达间质系表面标志物,如:CD13、CD29、CD44、CD106、CD90及CD166,而不表达造血干细胞表面标志物CD45及CD34;体外诱导培养后,gBMMSCs能够向脂肪细胞、成骨细胞和软骨细胞分化;以上结果说明我们分离获得的细胞具备MSCs的生物学特性。这一结果与Ren等[18,19]的报道相似,他们研究显示,绒山羊gBM-MSCs表达间质系表面标志物CD29、CD44和CD90,不表达内皮及造血干细胞表面标志物CD31及CD45,体外可以向骨细胞、神经细胞诱导分化。
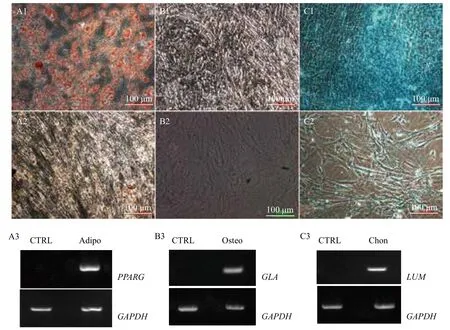
图6 体外诱导第5代阿尔巴斯绒山羊骨髓间充质干细胞向脂肪、成骨和软骨细胞分化
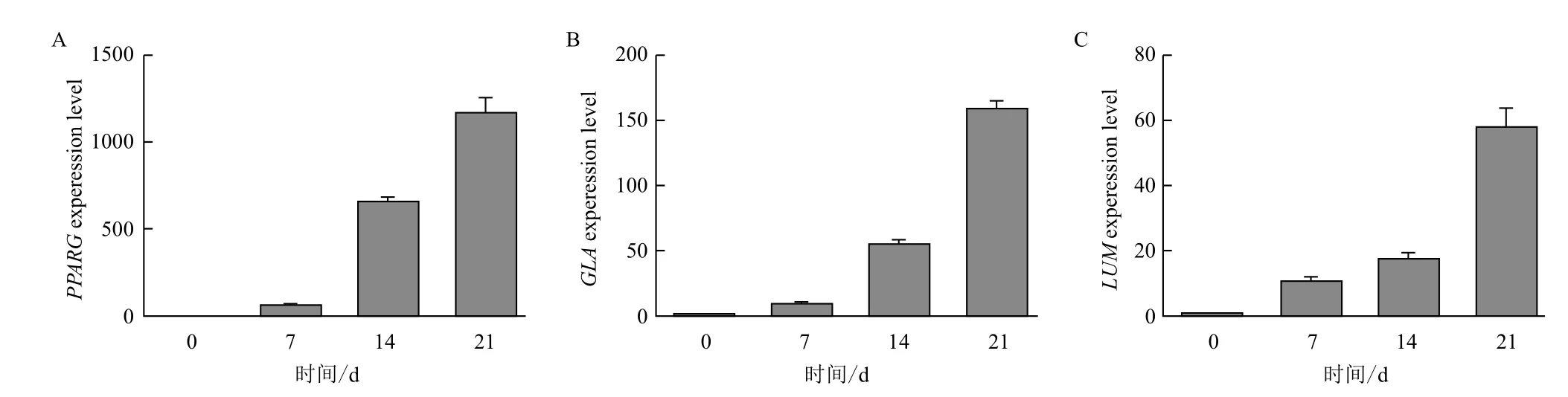
图7 实时定量PCR法检测诱导后分化相关基因的动态表达
在此基础上,我们通过荧光定量PCR法检测多向分化相关基因PPARG、GLA和LUM动态表达情况。Rosen等研究显示,PPARG通过结合脂肪细胞特异基因,如脂肪酸结合蛋白和脂肪细胞P2基因,在脂肪形成过程中扮演重要角色[20,21]。我们发现成脂肪诱导后PPARG表达量随着诱导天数的增加逐步上升。GLA是成骨特异性表达基因,与骨矿化基质的形成有关[22,23]。我们发现成骨诱导后GLA表达随着诱导天数的增加逐步上调,此结果与Açil等[24]的研究一致,他们发现人骨髓MSCs成骨分化过程中,碱性磷酸酶和骨钙蛋白表达上调。LUM编码形成亮氨酸丰富的蛋白多糖家族,包括聚糖、双链蛋白聚糖和纤维调节蛋白,也可能参与到胶原原纤维的构建[25]。我们发现成软骨诱导后LUM表达量同样随着诱导天数的增加逐步上升,诱导21 d时,表达显著上调。本实验发现体外诱导培养gBM-MSCs后,多向分化相关基因的表达水平随着诱导天数的增加呈上升趋势,进一步证实gBM-MSCs具有多向分化潜能。
综上所述,采用全骨髓贴壁培养法能够从阿尔巴斯绒山羊骨髓中分离gBM-MSCs,获得的gBMMSCs具有自我更新及多向分化潜能。此培养法操作简单,可大量分离及纯化gBM-MSCs,可以为后续转基因动物新品种培育及组织工程提供充足种子细胞,具有重要现实意义。
4 结论
本研究从阿尔巴斯绒山羊骨髓中分离培养获得了gBM-MSCs,该细胞符合国际细胞治疗协会提出的MSCs鉴定标准,具备MSCs生物学特性,可以用于后续实验研究。
[1]Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV. Osteogenesis in transplants of bone-marrow cells[J]. J Embryol Exp Morphol,1966, 16:381-390.
[2]Friedenstein AJ, Gorskaja JF, Kulagina NN. Fibroblast precursors in normal and irradiated mouse hematopoietic organs[J]. Exp Hematol, 1976, 4(5):267-274.
[3]Owen M, Friedenstein AJ. Stromal stem cells:marrow-derived osteogenic precursors[J]. Ciba Found Symp, 1988, 136:42-60.
[4]Caplan AI. Mesenchymal stem cells[J]. J Orthop Res, 1991, 9:641-650.
[5]Ahn MY, Zhang ZG, Tsang W, et al. Endogenous plasminogen activator expression after embolicfocal cerebral ischemia in mice[J]. Brain Res, 1999, 83(7):169-176.
[6]Campagnoli C, Roberts IA, Kumar S, et al. Identification of mesenchymal stem/progenitorcells in human first-trimester fetal blood, liver, and bone marrow[J]. Blood, 2001, 98:2396-2402.
[7]Zvaifler NJ, Marinova-Mutafchieva L, Adams G, et al. Mesenchymal precursor cells in theblood of normal individuals[J]. Arthritis Res, 2000, 2:477-488.
[8]Erices A, Conget P, Minguell JJ. Mesenchymal progenitor cells in human umbilical cord blood[J]. Br J Haematol, 2000, 109:235-242.
[9]Wang HS, Hung SC, Peng ST, et al. Mesenchymal stem cells in the Wharton's jelly of thehuman umbilical cord[J]. Stem Cells,2004, 22:1330-1337.
[10]In ‘t Anker PS, Scherjon SA, Kleijburg-van der Keur C, et al. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta[J]. Stem Cells, 2004, 22:1338-1345.
[11]Bruder SP, Kurth AA, Shea M, et al. Bone regeneration by implantation of purified, culture-expanded human mesenchymal stem cells[J]. J Orthop Res, 1998, 16:155-162.
[12]Shur I, Lokiec F, Bleiberg I, et al. Differential gene expression of cultured human osteoblasts[J]. J Cell Biochem, 2001, 83:547-553.
[13]Solmesky L, Lefler S, Jacob-Hirsch J, et al. Serum free cultured bone marrow mesenchymal stem cells as a platform to characterize the effects of specific molecules[J]. PLoS One, 2010, 5(9):e12689.
[14]Sotiropoulou PA, Perez SA, Salagianni M, et al. Characterization of the optimal culture conditions for clinical scale production of human mesenchymal stem cells[J]. Stem Cells, 2006, 24:462-471.
[15] Horwitz E, Le BK, Dominici M, et al. Clarification of the nomenclature for MSC:the international society for cellular therapy position statement[J]. Cytotherapy, 2005, 7:393-395.
[16]Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. the international society for cellular therapy position statement[J]. Cytotherapy,2006, 8:315-317.
[17] Burgener A. Cell culture technology for pharmaceutical and cellbased therapies[M]. USA:CRC Press, 2006:41-79.
[18]Ren Y, Wu H, Wang X, et al. Analysis of the stem cell characteristics of adult stem cells from Arbas white Cashmere goat[J]. Biochemical, 2014, 448(2):121-128.
[19]Ren Y, Wu H, Wang X, et al. The effect of arbas cashmere goatbone marrow stromal cells on production of transgenic cloned embryos[J]. Theriogenology, 2014, 81(9):1257-1267.
[20]Rosen ED, Spiegelman BM. PPARγ:a nuclear regulator of metabolism, differentiation, and cell growth[J]. J Bio Chem,2001, 276(41):37731-37734.
[21] Tontonoz P, Hu E, Graves RA, et al. PPAR γ2:tissue-specific regulator of an adipocyte enhancer[J]. Genes Dev, 1994, 8:1224-1234.
[22] Ryoo HM, Hoffmann HM, Beumer T, et al. Stage-specific expression of dlx-5 during osteoblast differentiation:involvement in regulation of osteocalcin gene expression[J]. Mol Endocrinol,1997, 11(11):1681-1694.
[23]Hauschka PV, Lian JB, Cole DE, et al. Osteocalcin and matrix gla protein vitamin K-dependent proteins in bone[J]. Physiol Rev,1989, 69(3):990-1047.
[24]Açil Y, Ghoniem AA, Wiltfang J. Optimizing the osteogenic differentiation of human mesenchymal stromal cells by the synergistic action of growth factors[J]. J Craniomaxillofac Surg,2014, 42(8):2002-2009.
[25]Pietraszek K, Chatron-Colliet A, Brezillon S, et al. Lumican:A new inhibitor of matrix metalloproteinase-14 activity[J]. FEBS Lett, 2014, 588(23):4319-4324.
(责任编辑 马鑫)
Isolation,Culture and Identification of Bone Marrow-derived Mesenchymal Stem Cells from Arbas Cashmere Goat
MA Su-ri-gu-ga1PENG Hang2LIU Jia-jia3ZHANG Yi-ting4GUI Hua1LIU Peng-xia1
(1. College of Life Sciences,Inner Mongolia University,Hohhot 010021;2. Bone and Joints Research Center,Frontier Institute of Science and Technology,Xi'an Jiaotong University,Xi'an 710061;3. Institute of Microbiology,Chinese Academy of Sciences,Beijing 100101;4. Center for Drug Safety Evaluation and Research,Shanghai Institute of Materia Medica,Chinese Academy of Sciences,Shanghai 201203)
Here we isolated and cultured Arbas cashmere goat bone marrow-derived mesenchymal stem cells(gBM-MSCs)in vitro,identified the biological characteristics of gBM-MSCs, and analyzed the dynamic expressions of differentiation genes while induced in vitro. The primary cultured gBM-MSCs were spindle cell-based, showing radial colony arrangement, and stably and continuously passed down for over 15 generations. The detections by immunofluorescence and flow cytometry showed that gBM-MSCs were positive for mesenchymal specific markers,such as CD13, CD29, CD44, CD106, CD90 and CD166, but negative for hematopoietic stem cell surface markers CD45 and CD34. The induction in vitro confirmed that gBM-MSCs owned the multiple differentiation potential, and differentiated into adipocytes, osteoblasts and chondrocyte. Real-time PCR further indicated that the expressions of differentiation genes increased throughout the culture period. Our results confirmed that the isolated and cultured cells from the bone marrow of Arbas cashmere goat possessed the biological characteristics of gBM-MSCs. Our findings promoted the basic research of MSCs in livestock and laid a theoretical foundation for the further application of gBM-MSCs in transgenic breeding and tissue engineering.
Arbas cashmere goat;mesenchymal stem cells;culture;differentiate;biological characteristics
10.13560/j.cnki.biotech.bull.1985.2016.02.028
2015-09-06
国家自然科学基金项目(31101698),内蒙古杰出青年自然科学基金项目(2011JQ03)
马苏日古嘎,女,硕士研究生,研究方向:干细胞生物学;E-mail:1017955583@qq.com
刘鹏霞,女,副教授,硕士生导师,研究方向:干细胞生物学;E-mail:Liupengxia@imu.edu.cn
