雅鲁藏布江缝合带南缘混杂岩放射虫组合及其地质意义
2016-09-27王天洋李国彪李新发牛晓路张金彪
王天洋,李国彪,李新发,牛晓路,张金彪,薛 嵩
(1.中国地质大学(北京) 生物地质与环境地质国家重点实验室,北京 100083;2.中国地质大学(北京) 地球科学与资源学院,北京 100083)
雅鲁藏布江缝合带南缘混杂岩放射虫组合及其地质意义
王天洋,李国彪,李新发,牛晓路,张金彪,薛嵩
(1.中国地质大学(北京) 生物地质与环境地质国家重点实验室,北京100083;2.中国地质大学(北京) 地球科学与资源学院,北京100083)
在西藏南部雅鲁藏布江缝合带南侧的吉隆地区出露有一套混杂岩,该混杂岩中见大量的放射虫化石。其研究成果可为恢复缝合带南缘混杂岩地层层序、探讨特提斯古海洋盆地演化、揭示印度-亚洲板块的全面碰撞过程等提供重要的生物地层学证据。将吉隆地区桑单林剖面自下而上划分为宗卓组、桑单林组和者雅组3个岩石地层单元;并在其中发现有晚白垩世—始新世放射虫化石,鉴定出50属、72种,划分了4个放射虫化石带:Immersothoraxcyclops带(晚白垩世Campanian期),Spongurusirregularis带(早古新世,RP1—RP3),Buryelladumitricai带(古新世中晚期,RP4—RP6),Phormocyrtisturgida带(始新世早期,RP7—RP8)。通过对古近纪时期特提斯喜马拉雅北亚带的地层、沉积等特征进行横向上的对比分析,认为古新世时处于特提斯喜马拉雅沉积带北亚带西段的吉隆地区为前渊环境,东段的江孜地区则为滨浅海沉积环境,表明西藏境内特提斯洋的闭合并不是同时发生的,而是沿雅鲁藏布江缝合带自东向西发生。
混杂岩;新特提斯洋;雅鲁藏布江缝合带;桑单林;放射虫
0 引 言
特提斯的演化过程一直是地学研究中的重要课题。特提斯的消亡及印度板块与亚洲板块的碰撞导致青藏高原的形成,它不但影响到整个地球的海陆分布格局,而且对全球气候变化造成了巨大的影响。西藏特提斯的消亡时间及过程仍然是一个长期存在争议的问题[1-6]。另一个重要问题是碰撞的方式:碰撞是由西向东迁移,还是由东向西迁移,或者由东西两侧向中间迁移?这一问题涉及碰撞后印度板块与欧亚板块的相互作用与方式。ROWLEY[7]通过对喜马拉雅地区最高海相层现有资料的总结,提出特提斯的闭合过程是自西向东发生的:50 Ma特提斯在巴基斯坦北部和印度西北部的Zanskar和Hazara地区最初关闭,渐新世末期在喜马拉雅东段完成关闭[7-9]。张克信、余光明等[10-11]通过对藏南古近纪岩相古地理的研究,认为印度与欧亚板块碰撞应首先发生于东部而后向中西部迁移。李祥辉等[12]则认为印度次大陆与亚洲板块的碰撞方式为西早东晚。
本次研究主要聚焦于雅鲁藏布江缝合带南侧吉隆桑单林剖面“混杂岩”放射虫生物地层序列的恢复(图1)。

图1 研究区构造单元划分(据李国彪等[8],有修改)Fig.1 The tectonic unit division in the study areaA.印度大陆;B.欧亚大陆;STDS.藏南拆离系

图2 西藏南部白垩纪—古近纪放射虫研究概况Fig.2 Cretaceous-Paleogene radiolarian in southern TibetA.仲巴县岗久东昌德布,蹬岗混杂岩;B.仲巴县卓弄,拉沙混杂岩;C.蹬岗组;D.郭雅拉组、盐多组(A—D据河北省地质调查院区调所[21]);E.桑单林,泥砂质混杂堆积(据李亚林等[20]);F.萨迦县赛区,宗卓组(据李国彪等[22]);G.康马县,大洋红层(据LI G B等[23]);H.萨嘎,吉隆群(据李红生[17]);I.江孜,羊卓雍混杂岩(据LIU J B, AITCHISON J C [24]);J—K.吉隆桑单林,宗卓组、蹬岗组、桑单林组(据LIANG Y P等[25]);L.萨嘎,桑单林组、者雅组(据丁林[18]);M.桑单林组,蹬岗组(据杨群等[26])
对于西藏南部地区的放射虫,前人多有研究[9,13-15];对于藏南吉隆桑单林地区沉积岩系的时代、地层划分及构造属性,一直存在着不同的观点(图2):盛金章根据放射虫组合特征将其时代确定为三叠纪(“吉隆群”)[13];万晓樵等[14]则根据浮游有孔虫组合特征认为,桑单林剖面基本包括了宗卓组的全部地层,其时代为晚白垩世早期的赛诺曼期至马斯特里赫特期末期;李红生[16-17]根据该剖面中发现的放射虫化石将其时代定为古新世晚期到早渐新世晚期;丁林[18]认为该套沉积代表了古新世欧亚大陆逆冲加载到印度被动大陆边缘产生的前陆盆地沉积,并将其定名为折巴群,包括下部富含硅质岩的桑单林组和上部以砂页岩为主的者雅组,时代为古新世;河北省区域地质矿产调查研究所[19]依据生物化石组合将桑单林—折巴地区一带的侏罗纪—白垩纪地层从原三叠系中分离出来,新建了达桑组(J1-2d)、旦嘎组(J3d)、折巴组(K1z)和桑单林组(K2s)和蹬岗组(E1-2dg)5个岩石地层单位。李亚林等[20]通过构造和变形研究认为:该套沉积建造并不是单一组成的岩石地层单元,而是在板块俯冲过程中于雅鲁藏布江缝合带前缘产生的典型构造混杂堆积,具非史密斯地层特点。本文根据在剖面中所采集到的放射虫化石及实测剖面岩性特征,将该剖面由下到上划分为宗卓组(K2z)、桑单林组(E1s)及者雅组(E1-2z)。
1 实测剖面描述
本次研究采样点位于藏南吉隆县北部的桑单林村北西的桑单林山,剖面起点坐标为29°15′33″N,85°14′46″E,高程4 805 m。剖面内可见2条断层及层间褶皱(图3)。该剖面岩性组合总体为钙质页岩,紫红色、灰绿色硅质岩,石英杂砂岩及灰褐色碳质页岩。
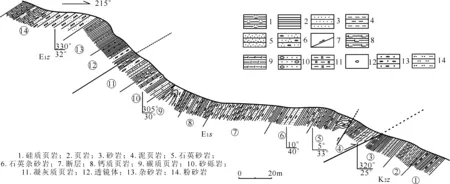
图3 西藏南部吉隆县桑单林剖面实测图Fig.3 Measured section of Sangdanlin at Gyirong,southern Tibet
现将剖面的岩性特征描述如下:
未见顶。
1.1者雅组(E1-2z)



1.2桑单林组(E1s)

⑩ 底部为厚层块状砂砾岩。砾石磨圆度、定向排列较好,成分主要为脉石英、石英砂岩、紫红色粉砂岩,砾径大小为2~5 cm。见重力流沉积,推测其为从大陆斜坡滑坡堆积形成。中下部为页岩夹透镜状分布的厚层块状岩屑石英杂砂岩。上部为深灰-灰褐色碳质、钙质页岩夹薄层状粉砂岩,砂岩具韵律层。厚56 m 。
⑨ 灰褐色碳质、钙质页岩与中厚层状岩屑石英杂砂岩互层,夹少量砂砾岩。页岩中夹眼球状砂屑灰岩结核(采集样品 :SD286—SD319)。砂屑灰岩结核中见放射虫:Calocyclomaampulla,Cromyechinuspycnopora等。厚165 m。
⑧ 黄绿色泥质页岩、灰白色厚层块状岩屑石英杂砂岩夹深灰色碳质、钙质和硅质页岩,砂岩中见重荷模及铁饼状粉砂岩结核。硅质岩及岩屑杂砂岩(采集样品:SD247—SD285)中可见放射虫:Amphicraspedumprolixum,Phaseliformasubcarinata,Stichomitracarnegiense等。厚55 m。
⑦ 灰绿色厚层块状岩屑石英杂砂岩与灰绿色、深灰色钙质页岩互层。砂岩中可见韵律层,韵律层中含有虫管。紫红色页岩中夹有砂岩透镜体,并可见大量钙质砂岩结核,呈团块状。紫红色页岩(采集样品:SD161—SD246)中见放射虫:Amphisphaeracoronata,Bekomasp.,Calocyclettatibeta,Halesiumtriacanthum,Lamptoniumfabaeformefabaeforme,Lychnocanomasp.,Lychnocanomacostata,Myllocercionaff.echtus,Orbulacomitata,Pantanelliumriedeli,Pessagnobrachiasp.,P.fabianii,Petasiformasp.,Phormocyrtisturgida,Pseudodictyomitrapentacloaensis,Squinabollumfossile,Stichomitrawero,?S.rutten,等。厚143 m 。
⑥ 灰褐色中厚层状岩屑石英杂砂岩夹页岩,页岩中发育黄铁矿结核,结核多为正方形,大小为3~4 cm。紫红色页岩夹钙质砂岩,可见破碎带,含糜棱岩。紫红色硅质页岩(采集样品SD97—SD160)中见放射虫:Archaeodictyomitralamellicostata,Buryellapentadica,Dactyliosphaeramaxima,Dictyomitraformosa,Holocryptocaniumbarbui,Lamptoniumfabaeformefabaeforme,L.pennatum,Patulibracchiumcf.davisi,Prunopyleadelstoma,P. sp.,Pseudoeucyrtispulchra,Spongodiscussp.,Spongurusirregularis,Theocapsommaamphora等。厚64 m。
⑤ 紫红色硅质、钙质页岩与灰白色中厚层块状岩屑石英杂砂岩互层,夹有岩屑石英杂砂岩透镜体。砂岩中可见韵律层及鲍马序列,岩屑含量较多。硅质页岩及页岩夹层(样品:SD68—SD96)中见放射虫:Acaeniotylestarka,Archaeodictyomitralacrimula,Cryptamphorellacf.conara,Dictyomitramulticostata,Hiscocapsasp.,Lychnocanomaauxilla,Lamptoniumsp.,Prunopyleadelstoma,Spongoprunumcrassum,Theocampelispa,T.salillum等。厚35 m。
④ 紫红色钙质、硅质页岩,黄绿色泥质页岩夹中层状硅质页岩,偶见深灰色硅质岩(采集样品:SD46—SD67)。硅质岩中产放射虫:Acaeniotylesp.,Amphisphaeracoronata,Buryelladumitricai,Dictyomitraaff.rhadina,Dictyomitrasp.,Hiscocapsacf.asseni,Lamptoniumfabaeformconstrictum,Patulibracchiumcf.davisi,Patellulaeuessceei,Pessagnobrachiasp.,Rhopalosyringiumkleinum,Stylosphaerahastate,Theocampetina,Turbocapsulacostata,Thanarlabrouweri,?Zifondiumplenum等。厚29 m。
1.3宗卓组(K2z)
③ 灰白色厚层块状石英粗砂岩、黄绿色页岩、灰绿色泥质页岩互层。顶部见粒序层理,含砾层厚约10 cm,砾石大小较为均一,分选、磨圆较好。与上覆硅质岩呈断层接触。采集样品SD34—SD45,见放射虫Archaeodictyomitralamellicostata,Cryptamphorellamacropora,C.conara,Dictyomitrasp.,Rhopalosyringiumelegans,Stichomitrastoki,Patellulaeuessceei,Pseudoaulophacusriedeli等。厚37 m。
② 深灰色钙质、碳质页岩夹透镜状硅质岩,夹灰白色中厚层块状石英粗砂岩。有冲刷接触,可见断裂破碎带,并有石英脉发育。砂岩中见放射虫:Cryptamphorellaconara,Theocampetina,Immersothoraxcyclops等。厚19 m。
① 泥页岩、灰绿色钙质页岩,页岩中夹有中薄层状杂砂岩,可见厚层块状岩屑杂砂岩,长石分化后为红色。厚20 m。
未见底。
2 材料与方法
2.1放射虫实体化石处理方法

图4 西藏吉隆桑单林剖面放射虫化石Fig.4 The radiolarian fossils of Sangdanlin section in southern Tibet图中线条比例尺为25 μm。标本均保存在中国地质大学(北京)地层与古生物教研室。1.Hiscocapsa sp.cf.H.asseni Tan, SD44; 2,10.Dictyomitra multicostata Pessagno, SD81; 3.Pseudoaulophacus riedeli Pessagno(破损), SD181; 4.Patulibracchium cf.P.davisi Pessagno, SD181; 5,12.Buryella tetradica Foreman(破损), SD379; 6.Amphisphaera aotea Hollis, SD109; 7.Holocryptocanium barbui Dumitrica, SD109; 8.Cryptamphorella conara Foreman, SD44; 9.Spongodiscus sp., SD105; 11.Lamptonium pennatum Foreman, SD109
将野外采集的样品碎至1~2 cm3的岩石碎块,放入500 mL的塑料烧杯中并用清水冲洗若干遍,倒去烧杯中浑浊的水,加入4%~5%的氢氟酸浸泡约24 h,倒入40目和300目的分析筛中进行滤洗,用清水冲洗多次至中性后将砂样盛入容器后晒干。重复多次以收集足够多的砂样。在实体显微镜下挑选放射虫化石标本,并粘在小实验台上。在扫描电镜下进行照相(图4),根据照片对比放射虫文献[25-41]进行属种的鉴定。
2.2放射虫硅质岩石切片法
将野外采集到放射虫硅质岩磨取薄片,在光学显微镜下观察放射虫内部显微结构形态,通过与实体化石对比以确定其属种。并观察岩石岩相,初步总结所采标本的古环境与古地理。
与有孔虫化石鉴定的方法类似,放射虫化石属种鉴定主要基于实体化石与切片化石两种方法。实体化石主要基于放射虫化石的外表形态及表面构造,而切片法则主要基于化石的外部轮廓与内部构造(图5),这两种方法各有优缺点。本文的放射虫化石鉴定采取实体化石与切片化石相结合的方法。
3 放射虫化石带划分
作者通过对桑单林剖面中所采集到的367个样品中所获得放射虫化石的研究,共鉴定放射虫50属72种,识别出4个带:Immersothoraxcyclops带,Spongurusirregularis带,Buryelladumitricai带,Phormocyrtisturgida带。各化石带的特征如图6所示。
3.1Immersothorax cyclops带
该带位于剖面第2—3层,优势分子为Cryptamphorellamacropora,C.conara,Dictyomitrasp.,Immersothoraxcyclops,Pseudoaulophacusriedeli等。该带化石Immersothoraxcyclops为晚白垩世Campanian期的典型属种,见于俄罗斯北部地区的晚白垩世地层中[33]。Cryptamphorellaconara一种的时代分布范围为Aptian期到Maastrichtian期,曾见于江孜地区所金剖面甲不拉组中部地层中[27]。Archaeodictyomitralamellicostata,Cryptamphorellaconara,Rhopalosyringiumelegans,Theocampetina等为晚白垩世常见放射虫分子。因此本化石带的时代归属于晚白垩世Campanian期。本带可与地中海地区放射虫Turbocapsulacostata带进行对比[28]。
3.2Spongurus irregularis 带
该带位于剖面第4—7层。优势分子为Spongurusirregularis等;其他分子有Dictyomitraaff.rhadina,Eusyringiumwoodsidensis,Haliommagr.,H. sp.,H. cf.asseni,Phaseliformasubcarinata,Prunopyleadelstoma,Pseudoeucyrtispulchra,Spongodiscussp.,Spongurusregularis,S.irregularis,Stichomitrawero,Theocapsommaamphora等。其中,带化石Spongurusirregularis的时代分布范围为早古新世(RP1—RP3),该种在本带中丰度较高。Dictyomitraaff.rhadina的时代分布范围为晚白垩世Maastrichtian到古新世早期(RP3);Haliommasp.为早古新世常见属种。因此Spongurusirregularis带的时代为早古新世。本带中混入有白垩纪放射虫

图5 西藏吉隆桑单林剖面放射虫化石Fig.5 The radiolarian fossils of Sangdanlin section in southern Tibet图中线条比例尺为25 μm;标本均保存在中国地质大学(北京)地层与古生物教研室。1. Lychnocanoma costata Nishimura, SD232; 2.Dictyomitra multicostata Pessagno, SD81; 3.Spongodiscus sp., SD105; 4.Mita regina Campell & Clark, SD393; 5.Hiscocapsa sp. cf. H.asseni Tan, SD75; 6. Petasiforma sp., SD181; 7. Dictyomitra formosa Squinabol, SD105; 8. Squinabollum fossile Squinabol, SD181; 9. Savaryella sp., SD45; 10. Lychnocanoma sp., SD75; 11. Pseudoeucyrtis pulchra Squinabol, SD109; 12,15.Buryella pentadica Foreman, SD378; 13. Pessagnobrachia sp., SD181; 14.Dictyomitra sp., SD109; 16. Patulibracchium cf. davisi Pessagno, SD181; 17. Cryptamphorella conara Foreman, SD44; 18.Calocycletta tibeta Ding, SD379; 19.Pseudoaulophacus riedeli Pessagno, SD181; 20. Hiscocapsa sp., SD81; 21.Calocycloma ampulla Ehrenberg, SD304; 22.Zifondium sp., SD44; 23.Calocycletta tibeta Ding, SD60; 24,26.Lithomespilus coronatus Squinabol, SD393; 25.?Zifondium plenum Wu, SD52; 27.Buryella tetradica Foreman, SD52; 28.Lychnocanoma sp., SD181; 29.Amphisphaera kina Hollis, SD45; 30.Phormocyrtis turgid Krasheninnikov, SD181

Archaeodictyomitralacrimula,Stichomitrawero,Theocampelispa等再沉积分子。
3.3Buryella dumitricai带
本带以Buryelladumitricai的始现面与Lychnocanomaauxilla的始现面分别作为其底、顶界线,位于剖面中上部(7—12层),其主要分子为Bekomasp.,Buryellainsensis,B.tetradica,B.dumitricai,B.pentadica,Calocyclettatibeta,Dictyomitramulticostata,Diplolegmasomphum,Lychnocanomaauxilla,L. sp.,Litheliusminor,Myllocercionaff.echtus,Spongodiscussp.,Stichomitracarnegiense。其中,Buryelladumitricai及Lychnocanomaauxilla见于新西兰Marlborough东部地区古新统,前者始现于早古新世晚期放射虫化石带RP4上部,而后者见于晚古新世晚期放射虫化石带RP6下部[28]。Buryellatetradica,B.dumitricai,Litheliusminor,Stichomitracarnegiense等为低纬度地区古新世放射虫组合常见分子,见于新西兰马尔堡东部的古新统放射虫硅质岩中[28]。按照HOLLIS等的分类,该化石带划分归属于古新世中晚期(RP4—RP6)[28]。
3.4Phormocyrtis turgida带
该带位于桑单林剖面顶部(12—14层),主要分子为Amphicraspedumprolixum,Amphisphaeracoronata,Buryellaclinata,Calocyclomaampulla,Cromyechinuspycnopora,Lamptoniumsp.,L.fabaeformefabaeforme,L.fabaeformconstrictum,Lithomespiluscoronatus,Orbulacomitata,Pseudodictyomitrapentacloaensis,Phormocyrtisturgida等。其中,Buryellaclinata,Calocyclomaampulla,Cromyechinuspycnopora,Lamptoniumfabaeformconstrictum,Phormocyrtisturgida等分子均为始新世早期属种,常见于地中海和墨西哥湾地区硅质岩中[35]。因此,该化石带时代为始新世早期(RP7—RP8)。本带见有晚白垩世放射虫Halesiumtriacanthum,Pantanelliumriedeli等再沉积分子。
4 讨 论
如上文所述,该剖面位于雅鲁藏布江缝合带南缘混杂岩带。前人对于吉隆地区中生代—古近纪沉积岩的时代、划分和构造属性一直存在着不同的观点[14,18,20]。本次研究表明:产于桑单林剖面下部(宗卓组)的放射虫分子总体上可与日喀则地区冲堆组以及西地中海等地区的白垩纪放射虫分子进行对比,而剖面中部(桑单林组)及上部(者雅组)的放射虫组合则总体上可与新西兰古近纪的放射虫组合进行对比[29]。下部宗卓组(K2z)的岩性主要为灰绿色钙质页岩、泥页岩、深灰色碳质页岩与灰白色厚层块状岩屑石英杂砂岩组成的砂泥质沉积,其中产出放射虫Immersothoraxcyclops,Theocampetina及Cryptamphorellaconara等分子,指示其年代为晚白垩世Campanian期,与上覆桑单林组硅质岩地层之间呈断层接触;中部桑单林组(E1s)沉积以紫红色、灰绿色钙质、硅质页岩,夹杂中厚层块状岩屑石英杂砂岩为主,可见放射虫Buryelladumitricai,Lychnocanomaauxilla,Spongodiscussp.等,其年代为早古新世晚期到晚古新世晚期(RP4—RP6)。顶部岩性为紫红色页岩与灰绿色凝灰质页岩互层,夹有中厚层状杂砂岩及灰岩透镜体。产出放射虫时代为晚古新世Thanetian期及早始新世。因此,综合剖面岩性特征及放射虫年代分析可以得出:本次研究所测的桑单林剖面下部宗卓组砂泥质沉积的时代应为晚白垩世,中部桑单林组富含硅质的砂页岩沉积的时代应为古新世,而上部者雅组含灰岩透镜体的凝灰质杂页岩沉积的时代则大致应为晚古新世Thanetian期及早始新世。
桑单林组岩性特征为紫红色页岩、硅质页岩,生物群为放射虫单相产出,判断其沉积相为远洋深水欠补偿盆地相,其顶部的者雅组岩性特征为灰绿色页岩与岩屑杂砂岩,夹少量硅质页岩,砂砾岩及灰岩透镜体并可见凝灰岩。生物群为放射虫及共生的浮游有孔虫,沉积相判断为半深海斜坡相。这代表的是特提斯洋壳的一部分破碎残片,说明在古新世晚期,吉隆地区为半深海—深海(前渊)环境,也就是特提斯的残留洋盆。而同处于特提斯喜马拉雅北带的江孜甲查拉组地层岩性特征为一套含岩屑和凝灰质的砂砾岩夹页岩沉积,含较为丰富的沟鞭藻和孢粉化石,其指示时代应属古新世—始新世早期[44]。值得指出的是:该地区虽有古近纪放射虫化石的报道[26],但其赋存岩石可能系外来岩块,未见古近纪完整的深水沉积序列,因此无法证明在该区发育古近纪深水沉积。综合其所含古生物与沉积特征以及所处构造位置分析,江孜地区应相当于周缘前陆盆地紧邻褶冲带前缘逆冲断裂的碰撞型扇三角洲沉积。同时甲查拉组的物源分析显示其物源来自于日喀则弧前盆地、雅鲁藏布江缝合带以及上三叠统郎杰学群,也倾向于指示甲查拉组发育在周缘前陆盆地中而非残留海盆地[45]。因而,江孜地区的甲查拉组在古新世晚期指示了一个水体较浅的滨浅海沉积环境,说明此时在江孜地区,特提斯洋壳已完全消失,印度与亚洲板块陆壳已经接触并发生碰撞。综上所述,可以得出:在西藏境内,特提斯洋海水的撤退总体上应是沿雅鲁藏布江缝合带自东向西发生的。
5 结 论
(1)本次研究在吉隆桑单林剖面发现大量放射虫化石,鉴定出50属72种,划分了Immersothoraxcyclops带(晚白垩世Campanian期),Spongurusirregularis带(早古新世,RP1—RP3)和Buryelladumitricai带(古新世中晚期,RP4—RP6)及Phormocyrtisturgida带(始新世早期,RP7—RP8)等4个放射虫化石带。
(2)桑单林剖面主要由3个岩石地层单位组成,下部宗卓组含有少量硅质成分的灰绿色钙质页岩、泥页岩、深灰色碳质页岩与灰白色厚层块状岩屑石英杂砂岩组成的砂泥质沉积,时代属晚白垩世;中部桑单林组主要为一套紫红色、灰绿色钙质、硅质页岩夹中厚层块状岩屑石英杂砂岩沉积,时代属于古新世;上部者雅组主要为一套含灰岩透镜体的凝灰质杂页岩沉积,其时代大致应为晚古新世Thanetian期—早始新世。
(3)江孜地区甲查拉组与吉隆地区者雅组为特提斯喜马拉雅沉积带北亚带东西两部晚古新世两套大致“同时异相”沉积地层,在古新世晚期,江孜地区为一个水体较浅的滨浅海沉积环境;吉隆地区为半深海-深海(前渊)环境,由此我们推断在东部特提斯洋壳消失、印度与亚洲板块已经发生陆陆碰撞的同时,在西部的吉隆地区尚残留着一个晚期洋盆、两侧陆壳也尚未进入碰撞阶段。因此,在西藏境内特提斯洋的闭合、海水的退去、印度板块与亚洲板块的碰撞过程,并不是同时发生的,而是沿雅鲁藏布江缝合带自东向西逐步发生。
[1]JAEGER J J, COURITILLOT V, TAPPONNIER P. Paleontological view of the ages of the Deccan Traps, the Cretaceous/Tertiary boundary and the India-Asia collision[J]. Geology, 1989, 17(4): 316-319.
[2]SAHNI A. The structure, sedimentation, and evolution of Indian continental margins[J]. Ocean Basins and Margins, 1982, 6: 353-398.
[3]KLOOTWIJK C, GEE J, PEIRCE J, et al. An early India-Asia contact: Paleomagnetic constraints from Ninetyeast Ridge, ODP Leg 121[J]. Geology, 1992, 20(5): 395.
[4]莫宣学,赵志丹,周肃,等. 印度—亚洲大陆碰撞的时限[J]. 地质通报, 2007, 26(10): 1240-1244.
[5]李国彪,万晓樵,刘文灿,等. 雅鲁藏布江缝合带南侧古近纪海相地层的发现及其构造意义[J]. 中国科学(D辑):地球科学, 2004, 34(3): 228-240.
[6]万晓樵. 西藏第三纪有孔虫生物地层及地理环境[J]. 现代地质, 1987, 1(1): 15-17.
[7]ROWLEY D B. Age of initiation collision between India and Asia: A review of the stratigraphic data[J]. Earth and Planetary Science Letters, 1996, 145(1): 1-13.
[8]SEARLE M P, WINDLEY B F, COWARD M P, et al. The closing of Tethys and the tectonics of the Himalaya[J]. Geological Society of America Bulletin, 1987, 98(6): 678-701.
[9]李国彪,万晓樵,刘文灿,等. 西藏南部古近纪微体古生物及盆地演化特征[M]. 北京: 地质出版社, 2005:1-157.
[10]张克信,王国灿,骆满生,等. 青藏高原新生代构造岩相古地理演化及其对构造隆升的响应[J]. 地球科学——中国地质大学学报, 2010, 35(5): 697-712.
[11]余光明,王成善,张哨楠. 西藏地区特提斯中生代沉积特征及沉积盆地演化[J]. 中国科学(B辑), 1989(9): 982-991.
[12]李祥辉,王成善,胡修棉,等. 朋曲组——西藏南部最高海相层位一个新的单元[J].地层学杂志, 2000, 24(3): 243-248.
[13]盛金章. 珠穆朗玛峰地区吉隆群的放射虫动物群:珠穆朗玛峰地区科学考察报告(1966-1968), 古生物(二)[M].北京: 科学出版社, 1976:125-136.
[14]万晓樵,丁林. 西藏吉隆白垩纪末期浮游有孔虫的发现及其年代意义[J]. 古生物学报, 2002, 41(1): 89-95.
[15]吴浩若. 西藏南部下鲁硅岩晚侏罗世罩笼虫(放射虫)新材料[J]. 现代地质, 2000, 14(3): 301-306.
[16]李红生,吴浩若. 西藏南部白垩系冲堆组的一些放射虫化石[J]. 微体古生物学报, 1985, 2(1): 61-77.
[17]李红生. 藏南古近纪放射虫岩的发现——一个迟到的发现报告[M]//第三届全国地层会议委员会.第三届全国地层会议论文集. 北京: 地质出版社, 2000: 354-358.
[18]丁林. 西藏雅鲁藏布江缝合带古新世深水沉积和放射虫动物群的发现及对前陆盆地演化的制约[J]. 中国科学(D辑), 2003, 33(1): 47-58.
[19]河北省区域地质矿产调查研究所, 石家庄经济学院. 1∶25万萨嘎县幅(H45C003001)、吉隆县幅(H45C004001)区域地质调查报告[R]. 石家庄: 河北省区域地质矿产调查研究所, 2002:1-70.
[20]李亚林,王成善,胡修棉,等. 西藏南部始新世早期放射虫动物群及其对特提斯闭合时间的约束[J]. 科学通报, 2007, 52(12): 1430-1435.
[21]河北省地质调查院区调所.1∶25万亚热幅、青兰县幅、霍尔巴幅、巴巴扎东幅(国内部分)地质调查成果与进展[J].沉积与特提斯地质,2005,25(1):80-86.
[22]李国彪,万晓樵,刘文灿,等. 藏南萨迦县赛区“混杂岩”放射虫的发现及其地质意义[J]. 地质通报, 2003, 22(10/11): 30-37.
[23]LI G B, JANSA L, WAN X Q, et al. Discovery of radiolaria from Upper Cretaceous Oceanic Red Beds in Daba, Kangmar and its paleogeographic implication[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 312(1/2): 127-137.
[24]LIU J B, AITCHISO J C. Upper Paleocene radiolarians from the Yamdrok mélange, south Xizang(Tibet), China[J]. Micropa-leontology, 2002, 48(Suppl. 1): 145-154.
[25]LIANG Y P, ZHANG K X, XU Y D, et al. Late Paleocene radiolarian fauna from Tibet and its geological implications[J]. Canadian Journal of Earth Sciences, 2012, 49(11): 1364-1371.
[26]杨群,松岗笃,王玉净. 西藏南部放射虫微体古生物研究进展[J]. 微体古生物学报, 2002, 19(2): 105-111.
[27]赵胜楠. 藏南江孜所金白垩纪放射虫研究[D]. 北京: 中国地质大学(北京), 2013: 1-64.
[28]HOLLIS C J. Cretaceous-Paleocene Radiolaria from Eastern Marlborough, New Zealand[M]. Lower Hutt: Institute of Geological & Nuclear Sciences Limited, 1997: 1-152.
[29]HOLLIS C J. Biostratigraphy and paleoceanographic significance of Paleocene radiolarians from offshore eastern New Zealand[J]. Marine Micropaleontology, 2002, 46(3/4): 265-316.
[30]HOLLIS C J. Latest Cretaceous to Late Paleocene radiolarian biostratigraphy: A new zonation from the New Zealand region[J]. Marine Micropaleontology, 1993, 21(4): 295-327.
[31]O’DOGHERTY L. Biochronology and Paleontology of Mid-Cretaceous Radiolarians from Northern Apennines (Italy) and Betic Cordillera (Spain)[M]. Lausanne:Universite de Lausanne, 1994: 1-413.
[32]LI G B, WAN X Q, LIU W C, et al. A new Cretaceous age for the Saiqu “mélange,” southern Tibet: evidence from radiolaria[J]. Cretaceous Research, 2009, 30(1): 35-40.
[33]POPOVA G I, VISHNEVSKAYA V, BAUMGARTNER P O. Upper Cretaceous (Santonian-Campanian) radiolarians from Voronesh Anticline, southwestern Russia[J]. Micropaleontology, 2005, 51(1): 1-37.
[34]RIEDEL W R, SANFILIPPO A. Stratigraphy and evolution of tropical Cenozoic radiolarians[J]. Micropaleontology, 1978, 24(1): 61-96.
[35]SANFILIPPO A, HAKYEMAZ A, TEKIN U K. Biostratigraphy of Late Paleocene-Middle Eocene radiolarians and foraminifera from Cyprus[J]. Micropaleontology, 2003, 49(1): 47-64.
[36]O’CONNOR B. Buryella (Radiolaria, Artostrobiidae) from DSDP Site 208 and ODP Site 1121[J]. Micropaleontology, 2001, 47(1): 1-22.
[37]HARRISON T M, LOVERA O M, GROVE M, et al. New insights into the origin of two contrasting Himalayan granite belts[J]. Geology, 1997, 25 (10): 899-902.
[38]SEIDERS V M, PESSAGNO E A, HARRIS A G. Radiolarians and conodonts from pebbles in the Franciscan assemblage and the Great Valley Sequence of the California Coast Ranges[J]. Geological Society of America Bulletin, 1979, 7(1): 37-40.
[39]RADHAKRISHNA T, JOSEPH M. Geochemistry and paleomagnetism of Late Cretaceous mafic dikes in Kerala, southwest coast of India in relation to large igneous provinces and mantle plumes in the Indian Ocean region[J]. Geological Society of America Bulletin, 2012, 124(1/2): 240-255.
[40]ROWLEY D B. Minimum age of initiation of collision between India and Asia North of Everest based on the subsidence history of the Zhepure Mountain Section[J]. The Journal of Geology, 1998, 106(2): 229-235.
[41]SCHELLING D. The tectonostratigraphy and structure of the eastern Nepal Himalaya[J]. Tectonics, 1992, 11(5): 925-943.
[42]刘云龙,李国彪. 西藏南部岗巴地区白垩纪化石碳酸盐微相与沉积环境研究[J]. 现代地质, 2012, 26(1): 22-35.
[43]ZEITLER P K, CHAMBERLAIN C P. Petrogenetic and tectonic significance of young leucogranites from the northwestern Himalaya, Pakistan[J]. Tectonics, 1991, 10(4): 729-741.
[44]李国彪,万晓樵,丁林,等. 藏南古近纪前陆盆地演化过程及其沉积响应[J]. 沉积学报, 2004, 22(3): 455-464.
[45]蔡福龙,丁林,王厚起,等. 特提斯喜马拉雅带江孜地区古近纪地层源区分析:对造山带早期地壳加厚的制约[J]. 地质科学, 2013, 48(2): 435-448.
Radiolarian Assemblage from the Mélange in the Southern Margin to Yarlung-Zangbo Suture and Its Geological Significance
WANG Tianyang, LI Guobiao, LI Xinfa, NIU Xiaolu, ZHANG Jinbiao, XUE Song
(1.StateKeyLaboratoryofEnvironmentalGeologyandBiogeology,ChinaUniversityofGeosciences,Beijing100083,China;2.SchoolofEarthSciencesandResources,ChinaUniversityofGeosciences,Beijing100083,China)
A set of mélange is exposed in Sangdanlin, Gyirong, south to the Yarlung-Zangbo Suture (YZS). The measured Sangdanlin section is lithostratigraphically divided into Zongzhuo Formation, Sangdanlin Formation and Zheya Formation from bottom to top. Abound radiolarian fossils were gained from the mélange and 72 species of 50 genera were identified, which allows the recognizing of four radiolarian assemblages:Immersothoraxcyclops,Spongurusirregularis,BuryelladumitricaiandPhormocyrtisturgidazones. The radiolarian assemblage points to a late Cretaceous to early Eocene age for the strata of the Sangdanlin section. The research results provide the important paleontologic evidence to restore the stratigraphic sequence of “mélange” in the southern margin of YZS, and they also explore the evolution of the Neotethys and reveal the comprehensive collision process between the India and the Asia plates.
mélange; Neotethys; Yarlung-Zangbo Suture; Sangdanlin; radiolarian
2015-10-25;改回日期:2016-01-24;责任编辑:戚开静。
中国科学院战略性先导科技专项(B类)子课题(XDB050105003);国家重点基础研究发展计划(2012CB 822001);中国地质大学(北京)生物地质与环境地质国家重点实验室开放性课题(GBL21312);国家自然科学基金项目(40972026);中央高校基本科研业务费优秀教师研究项目(2011YXL012)。
王天洋,男,硕士研究生,1991年出生,古生物学与地层学专业,主要从事微体古生物学方面的研究工作。
Email:guess19911029@163.com。
李国彪,男,教授,博士生导师,1968年出生,古生物学与地层学专业,主要从事微体古生物学的科研和教学工作。Email:liguobiao@cugb.edu.cn。
P542;Q91
A
1000-8527(2016)02-0445-10
