大豆GmAP1基因的功能初探
2016-09-14宋拉拉张晓玫陈福禄傅永福赵德刚
宋拉拉张晓玫陈福禄傅永福赵德刚
(1. 贵州大学农业生物工程研究院 生命科学学院山地植物资源保护与种质创新省部共建教育部重点实验室,贵阳 550025;2. 中国农业科学院作物科学研究所 国家农作物基因资源与遗传改良重大科学工程 农业部北京大豆生物学重点实验室,北京 100081)
大豆GmAP1基因的功能初探
宋拉拉1张晓玫2陈福禄2傅永福2赵德刚1
(1. 贵州大学农业生物工程研究院 生命科学学院山地植物资源保护与种质创新省部共建教育部重点实验室,贵阳 550025;
2. 中国农业科学院作物科学研究所 国家农作物基因资源与遗传改良重大科学工程 农业部北京大豆生物学重点实验室,北京 100081)
以大豆天隆1号为材料,根据Glycine max Wm82.a2.v1中大豆基因组的预测序列,克隆出一个APl同源基因,命名为GmAPl。该基因编码区CDS长度为711 bp,编码一个含236个氨基酸的蛋白质。对蛋白质序列的结构分析结果显示GmAP1蛋白符合MADS-box基因家族特征。为了初步分析GmAPl的功能,利用实时定量RT-PCR分析GmAPl在不同花器官中表达,并且在拟南芥中过表达该基因。在不同花器官中,GmAPl表达量不同,在花萼中表达量较高;花瓣次之。GmAPl过表达植株表现出早花、植株矮小以及花器官(花瓣、雄蕊和雌蕊)数量增多等表型。结果表明,大豆的GmAPl是一个功能保守的APl基因,在花的发生和花器官发育中起着重要作用。
开花;花萼;GmAPl;MADS基因;大豆
植物开花是植物生长发育的一个重要过程[1]。植物响应并整合环境和内源信号,产生相应的反应信号,然后以网络应答的方式将新信号传递给开花整合因子,再由其促进开花基因LEAFY(LFY)和APETALAl(APl)表达,最终促使植物开花[2,3]。APl对控制植物花分生组织特性与花器官形成具有重要的作用[4,5],属于MADS-box基因家族[6,7]。在大豆[8]、苹果[9]、百合[10]、樱桃[11]、建兰[12]和杨树[13]等多种植物中已分别克隆出APl同源基因。异位表达各种植物的APl基因,均促进拟南芥或烟草开花,且此性状可遗传到子代[14]。
大豆(Glycine max)起源于中国,是植物性蛋白的主要来源和重要的工业原料[15]。大豆开花影响籽粒产量,对高产育种具有重要作用[16]。传统方法通过人工调控(控制光照、改变温度、使用激素等)花期,而转基因技术较多集中于大豆抗病虫害、抗逆性及大豆油分等方面研究[17],应用转基因控制大豆开花从而提高产量方面研究较少。采用分子遗传学和分子生物学改良品种,将促进开花的APl基因导入短日照植物中,改变植物开花对短日照的依赖,促进开花,提高产量,可加快育种进程。大豆基因组具有多拷贝特征,APl同源基因有4个拷贝[18],本研究以其中一个APETALl(GmAPl)同源基因为目标,利用植物遗传转化技术将该基因转入到拟南芥中,对其功能进行初步分析,探讨其功能与大豆花发育的关系,为进一步研究大豆花发育的分子机理提供理论依据。
1 材料与方法
1.1材料
实验中植物材料为大豆品种天隆1号,种植于中国农业科学院作物科学研究所人工气候室(16 h光照,8 h黑暗,25℃),开花期取花蕾,提取RNA,用于基因克隆,分别取花蕾的花萼、花瓣、雄蕊和雌蕊,提取RNA,用于表达分析。拟南芥(Arabidopsis thaliana)为Columbia(Col-0)生态型背景。拟南芥突变体apl.3由中国科学院植物研究所孟征研究员惠赠。大肠杆菌DH5α,农杆菌GV3101,载体Fu76-35S、Fu28和Fu39-2[19]均由本实验室保存备用。实验所用的高保真DNA聚合酶,反转录试剂盒、质粒提取试剂盒,琼脂糖凝胶回收试剂盒分别购自TaKaRa公司、博迈德生物公司和天根生物技术公司。实验用引物由华大科技公司合成,序列测定由北京睿博兴科生物技术有限公司完成。
1.2方法
1.2.1APl克隆及表达载体构建 根据Phytozome数据库中Glycine max Wm82.a2.v1预测基因序列,设计引物(表1)扩增GmAPl基因的编码序列,根据北京全式金生物技术有限公司RNA提取试剂盒提取天隆1号RNA,反转录反应总体积10 μL:5× PrimeScript Buffer 2 μL,PrimeScript RT Enzyme Mix I 0.5 μL,Oligo dT Primer 0.5 μL,Random 6 mers 0.5 μL,总RNA 1 μL(500 ng/μL)用RNase Free H2O补足至10 μL。通过37℃ 15 min,85℃ 5 s,完成反转录反应。PCR体系:2×PrimeSTAR GC Buffer 25 μL,dNTP Mixture 4 μL,正反向引物各1 μL(10 μmol/L),模板5 μL,PrimeSTAR HS DNA Polymerase 0.5 μL,用灭菌水补足至50 μL。PCR反应程序为:94℃ 3 min;98℃ 10 s,60℃ 30 s,72℃ 1 min/kb,循环35次;72℃ 7 min。用1.0%琼脂糖凝胶电泳检测扩增产物。回收PCR产物,与Fu28载体连接;连接产物转化大肠杆菌DH5α,挑取单克隆,通过菌落PCR鉴定阳性克隆。阳性克隆委托北京睿博兴科生物技术有限公司测序,序列测序正确的质粒通过LR反应到表达载体Fu39-2,转化大肠杆菌DH5α,挑取单克隆,通过菌落PCR鉴定阳性克隆。阳性克隆委托北京睿博兴科生物技术有限公司测序,最后转化农杆菌GV3101,农杆菌花浸染法转化拟南芥。
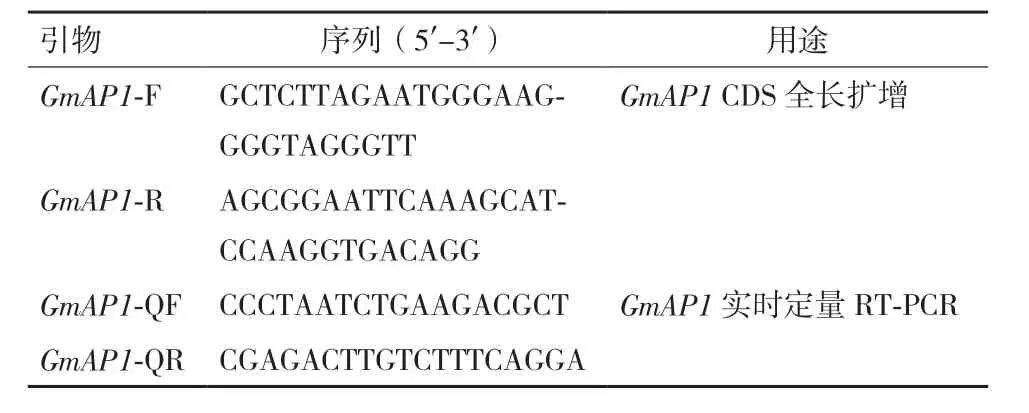
表1 本研究所用引物序列
1.2.2实时定量RT-PCR 在转录本序列设计实时定量PCR特异引物(表1),扩增长度113 bp。PCR产物转化大肠杆菌DH5α,挑取单克隆,通过菌落PCR鉴定阳性克隆,委托北京睿博兴科生物技术有限公司测序,序列与扩增序列完全匹配。实时定量PCR的反应体系:SYBP Premix Ex Taq 7.5 μL,ROX reference Dye 0.3 μL,正反引物各1.5 μL(2 μmol/L),cDNA 1.5 μL(RNA 325 ng),用超纯水补足15 μL。生物重复3次。实时定量PCR反应程序:95℃ 30 s;95℃ 5 s,60℃ 30 s,40个循环。以ACTll为内参基因,超纯水作模板为阴性对照,采用2-△△Ct法计算相对表达量。
2 结果
2.1GmAPl编码蛋白聚类分析及其序列特征
分析表明GmAPl(基因序列号:Glyma08g36380)编码236个氨基酸(图1),其中包含MADS-box结构域和K-box结构域(图2),属于MADS-box基因家族成员。经Blast进行同源序列比对,GmAPl氨基酸序列与豇豆(Vigna unguiculata)、百脉根(Lotus japonicus)、鸡豆(Cicer arietinum)、蒺藜苜蓿(Medicago truncatula)、豌豆(Pisum sativum)、红豆(Vigna angularis)和大枣(Ziziphus jujuba)等物种中的APl或APl-like的同源性达到80%以上(图3)。其中MADS-box结构域和K-box结构域高度保守,C-末端以CF结尾,而不是以法尼基转移酶的识别位点CFAA结尾。

图1 GmAP1编码区的核酸序列及编码的氨基酸序列
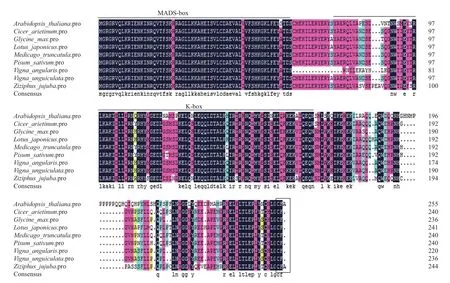
图2 GmAP1与其他AP1同源基因氨基酸序列比对
2.2大豆GmAPl cDNA全长的获得
以拟南芥APETALAl(APl)基因为参照,根据Phytozome数据库中大豆基因组Glycine max Wm82.a2.v1序列预测的GmAPl基因序列设计引物,以大豆cDNA为模板进行PCR扩增,得到750 bp左右的特异条带(图4)。测序结果表明该片段长度为711 bp,与Glycine max Wm82.a2.v1中的预测基因序列相同,且和已报道APl同源基因的同源性较高,表明该片段是大豆的APl同源基因。
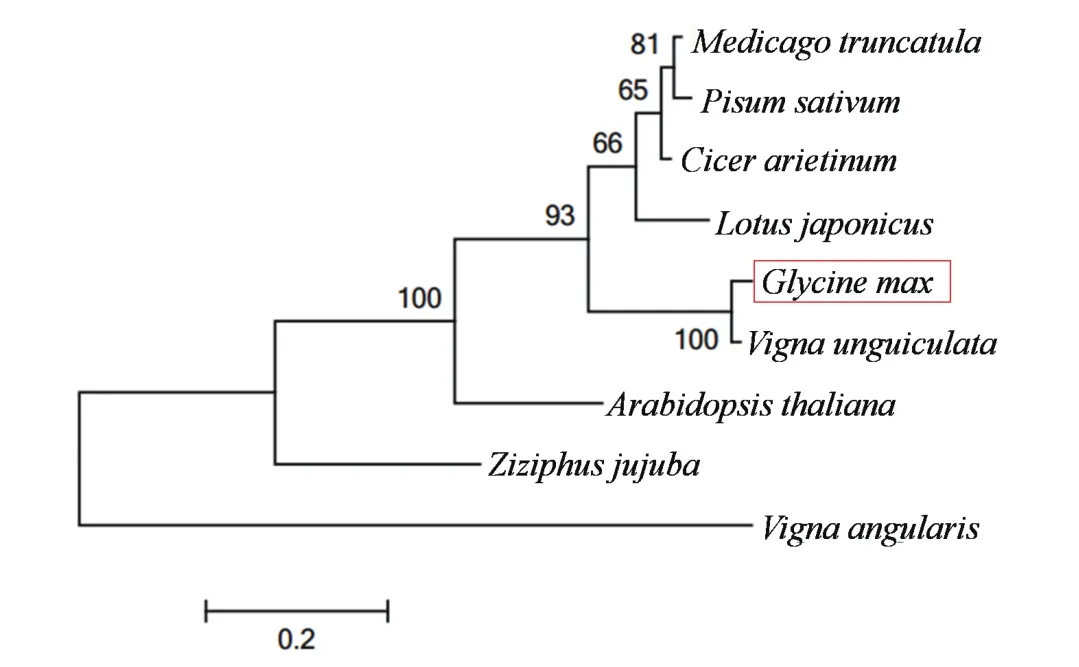
图3 GmAP1与其他AP1同源基因的聚类分析

图4 GmAP1 CDS全长扩增产物
2.3GmAPl表达分析
利用实时定量RT-PCR(Real time-PCR)的方法检测GmAPl在大豆不同花器官的表达情况。结果(图5)表明,在大豆不同花器官中GmAPl的表达有显著差异。GmAPl在萼片中表达量较高,在花瓣中表达量次之,在雄蕊和心皮中也有少量表达量。
2.4花浸染法转化拟南芥
为进一步分析GmAPl的功能,将由35S启动GmAPl基因的载体转化拟南芥,获得转基因植株,观察p35S∷GmAPl转基因植株和野生型表型。结果(图6)显示,在长日条件下(16 h光照,8 h黑暗),p35S∷GmAPl转基因植株比野生型植株早花,p35S∷GmAPl转基因植株在播种后13-15 d就开花,此时莲座叶数仅为两片,且株型小,花数量少;而野生型在播种后22-24 d才开花,莲座叶数为9-11片。p35S∷GmAPl转基因植株与野生型的花型不同,拟南芥野生型花有4个花萼,4个花瓣,6个雄蕊,1个雌蕊,而p35S∷GmAPl转基因植株花有4个花萼,8个花瓣,10雄蕊和2个雌蕊(图7)。

图5 GmAP1在大豆不同花器官中的表达

图6 GmAP1过表达对植株开花的影响

图7 异位表达GmAP1转基因拟南芥表型分析
3 讨论
本研究成功克隆得到大豆APl同源基因GmAPl,其氨基酸序列与许多物种中APl同源基因编码的氨基酸序列同源性达80%以上,并与豇豆(Vigna unguiculata)、百脉根(Lotus japonicus)、鸡豆(Cicer arietinum)、蒺藜苜蓿(Medicago truncatula)、豌豆(Pisum sativum)等豆科植物在一个分支上。GmAPl的MADS-box和K结构域相对比较保守,这些结果表明,大豆GmAPl属于MADS-box家族基因。但是,GmAP1蛋白C-末端以CF结尾,而不是以法尼基转移酶的识别位点CFAA结尾。
APl基因属于A类基因[20],调控花萼和花瓣的发育[21-23]。实时定量RT-PCR分析表明,GmAPl在大豆不同花器官中表达量不同,在花萼和花瓣中表达量相对于雄蕊和雌蕊较高。在拟南芥中,APl同源基因作为花分生组织决定基因,在花原基发育早期阶段发挥作用[6,24];作为花器官特异基因,决定花器官起始和发育[4,5]。不同物种的APl同源基因异位表达均早花,但花器官结构的改变却不相同。在烟草中过表达大豆Jackson品种的APl-like基因(Glyma16g13070.1),表现出早花,植株矮小,正常花瓣处出现了雄蕊状的花瓣[8]。苹果中APl-like基因MdMADS5过表达使拟南芥早花,花序较短,花蕾不同,尤其是花萼[9]。在烟草和拟南芥中过表达桦树PsnAPl-l和PsnAPl-2基因,表现出早花,植株矮小,融合终端花,花畸形[25]。本研究在拟南芥中过表达大豆天隆1号品种的GmAPl基因,植株表现出早花,植株矮小,花数量少,花器官(花瓣、雄蕊和雌蕊)数增多。因此,GmAPl基因对大豆开花时间及花器官的形成可能有重要作用。
大豆APl基因是多拷贝基因[18],Chi等[8]在烟草中过表达大豆Jackson品种的APl-like基因,表现出早花,植株矮小,正常花瓣处出现了雄蕊状的花瓣。本研究以另一个APl同源基因为目标,对该基因进行表达分析,以及在拟南芥中过表达该基因进行初步的功能分析,与Chi等早花结果一致,但花器官结构的改变不同。结果说明,大豆中不同GmAPl基因拷贝的功能并不完全相同,它们在进化过程中发生了功能的分化。对这两个GmAPl基因进行比较研究分析,将进一步解析它们在功能上的协作与特化机制。
4 结论
本研究克隆获得大豆天隆1号GmAPl基因,并证实该基因参与大豆开花时间以及花器官形态建成的调控。
[1]陈晓, 陈彦惠, 任永哲. 植物开花转换的分子生物学研究[J].分子植物育种, 2005(4):557-565.
[2]Weigel D, Alvarez J, Smyth DR, et al. LEAFY controls floral meristem identity in Arabidopsis[J]. Cell, 1992, 69(5):843-859.
[3]Wagner D, Wellmer F, Dilks K, et al. Floral induction in tissue culture:a system for the analysis of LEAFY-dependent gene regulation[J]. The Plant Journal, 2004, 39(2):273-282.
[4] Gustafson-Brown C, Savidge B, Yanofsky MF. Regulation of the Arabidopsis floral homeotic gene APETALA1[J]. Cell, 1994, 76(1):131-143.
[5] Wagner D, Sablowski RWM, Meyerowitz EM. Transcriptional activation of APETALA1 by LEAFY[J]. Science, 1999, 285(5427):582-584.
[6] Mandel MA, Gustafson-Brown C, Savidge B, et al. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1[J]. Nature, 1992, 360(19):273-277.
[7] Weigel D, Meyerowitz EM. Activation of floral homeotic genes in Arabidopsis[J]. Science, 1993, 261(24):1723-1726.
[8]Chi Y, Huang F, Liu H, et al. An APETALA1-like gene of soybean regulates flowering time and specifies floral organs[J]. Journal of Plant Physiology, 2011, 168(18):2251-2259.
[9]Kotoda N, Wada M, Kusaba S, et al. Overexpression of MdMADS5,an APETALA1-like gene of apple, causes early flowering in transgenic Arabidopsis[J]. Plant Science, 2002, 162(5):679-687.
[10] Chen MK, Lin IC, Yang CH. Functional analysis of three lily(Lilium longiflorum)APETALA1-like MADS box genes in regulating floral transition and formation[J]. Plant and Cell Physiology,2008, 49(5):704-717.
[11]Wang J, Zhang X, Yan G, et al. Over-expression of the PaAP1 gene from sweet cherry(Prunus avium L. )causes early flowering in Arabidopsis thaliana[J]. Journal of Plant Physiology, 2013, 170 (3):315-320.
[12]吴菁华, 吴少华, 杨超, 等. 建兰 AP1 基因的克隆, 表达及其与 MADS-box 转录因子相互作用的分析[J]. 园艺学报,2013, 40(10):1935-1942.
[13]李爽, 郑唐春, 代丽娟, 等. 小黑杨 PsnAP1-1 及 PsnAP1-2 基因的克隆及原核表达分析[J]. 植物研究, 2014, 34(4):498-504.
[14]Peña L, Martín-Trillo M, Juárez J, et al. Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time[J]. Nature Biotechnology, 2001, 19(3):263-267.
[15]邱丽娟, 王昌陵, 周国安, 等. 大豆分子育种研究进展[J].中国农业科学, 2007, 40(11):2418-2436.
[16]赵双进, 唐晓东, 赵鑫, 等. 大豆开花落花及时空分布的观察研究[J]. 中国农业科学, 2013, 46(8):1543-1554.
[17]任海祥, 南海洋, 曹东, 等. 大豆转基因技术研究进展[J].东北农业大学学报, 2012, 43(7):6-12.
[18]胡瑞波, 范成明, 李宏宇, 等. 大豆MIKC型MADS—box 基因家族分析[J]. 分子植物育种, 2009, 7(3):429-436.
[19]Wang X, Fan CM, Zhang XM, et al. BioVector, a flexible system for gene specific-expression in plants[J]. BMC Plant Biology, 2013,13(198):1471-2229.
[20]胡丽芳, 金志强, 徐碧玉. MADS-box 基因对花的发育及开花早晚的影响[J]. 生命科学研究, 2004, 8(4):7-12.
[21]Bowman JL, Alvarez J, Weigel D, et al. Control of flower development in Arabidopsis thaliana by APETALA1 and interacting genes[J]. Development, 1993, 119:721-743.
[22]Yalovsky S, Rodríguez-Concepción M, Bracha K, et al. Prenylation of the floral transcription factor APETALA1 modulates its function[J]. The Plant Cell, 2000, 12(8):1257-1266.
[23]Ng M, Yanofsky MF. Activation of the Arabidopsis B class homeotic genes by APETALA1[J]. The Plant Cell, 2001, 13(4):739-753.
[24] Kempin SA, Savidge B, Yanofsky MF. Molecular basis of the cauliflower phenotype in Arabidopsis[J]. Science, 1995, 267 (5197):522-525.
[25]Zheng TC, Li S, Zang LN, et al. Overexpression of two PsnAP1 genes from Populus simonii× P. nigra causes early flowering in transgenic tobacco and Arabidopsis[J]. PLoS One, 2014, 9(10):e111725.
(责任编辑 马鑫)
Functional Analysis of Soybean GmAP1
SONG La-la1ZHANG Xiao-mei2CHEN Fu-lu2FU Yong-Fu2ZHAO De-gang1
(1. The Key Lab of Plant Resources Conservation and Germplasm Innovation in Mountainous Region,Ministry of Education,Institute of Agrobioengineering and College of Life Sciences,Guizhou University,Guiyang 550025;2. MOA Key Lab of Soybean Biology(Beijing),National Key Facility of Crop Gene Resource and Genetic Improvement,Institute of Crop Sciences,Chinese Academy of Agricultural Sciences,Beijing 100081)
The full length GmAPl gene,a homologous gene of APl,was cloned from the soybean cultivar Tianlong 1 based on the putative sequence of soybean genome in Glycine max Wm82.a2.v1 database. GmAPl was 711 bp in length and encoded a protein of 236 amino acids. The structure analysis of protein sequence showed that GmAP1 protein characterized as MADs-box gene family. To analyze the function of GmAPl,the expression patterns of GmAPl in different floral organs were analyzed by RT-PCR,and GmAPl was overexpressed in Arabidopsis. The results showed that GmAPl differently expressed in different floral organs with the highest level in sepals,following by the petals. Transgenic Arabidopsis plants displayed phenotypes of early flowering,dwarf,and increasing number of sepals,petals,and stamens. The result indicated that GmAPl was one of APl homologous genes in soybean with conserved function and played an important role in flowering and floral organ development morphogenesis.
flowering;sepals;GmAPl;MADS gene;soybean
10.13560/j.cnki.biotech.bull.1985.2016.08.013
2015-12-21
国家自然科学基金项目(31371703),转基因生物新品种培育重大专项(2014ZX08004-005)
宋拉拉,女,硕士研究生,研究方向:植物发育分子生物学;E-mail:lalasong163@163.com
傅永福,男,研究员,博士生导师,研究方向:植物发育生物学;E-mail:fuyongfu@caas.cn赵德刚,男,教授,博士生导师,研究方向:植物基因工程;E-mail:dgzhao@gzu.edu.cn
