迁飞对草地螟生殖行为及种群暴发的影响
2016-09-14宋文一程云霞罗礼智仵均祥江幸福
宋文一, 程云霞, 罗礼智, 张 蕾, 仵均祥, 江幸福*
(1. 西北农林科技大学植物保护学院,应用昆虫学重点实验室,杨凌 712100;2. 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193)
迁飞对草地螟生殖行为及种群暴发的影响
宋文一1,2,程云霞2,罗礼智2,张蕾2,仵均祥1*,江幸福2*
(1. 西北农林科技大学植物保护学院,应用昆虫学重点实验室,杨凌712100;2. 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京100193)
为明确野外种群迁飞对草地螟生殖行为的影响及其在种群大暴发中的作用,对草地螟野外迁飞和未迁飞种群的生殖特征,包括首次产卵历期(period of first oviposition, PFO)及其他生殖参数进行了系统分析。结果表明,经历迁飞的种群与未经历迁飞的种群相比,成虫产卵前期、产卵历期、PFO和寿命均显著缩短;交配次数及交配率均显著增加;但平均每雌产卵量和卵孵化率无显著变化。这说明,尽管经历迁飞的草地螟种群产卵量未显著增加,但成虫到达迁入地后将很快多次交配、产卵,且产卵前期显著提前、产卵同步性显著增加,从而加剧后代种群暴发。
草地螟;迁飞种群;未迁飞种群;PFO
迁飞行为是昆虫对季节性变化和不稳定生境的一种适应性对策,也是寻觅新生境的一种行为对策[1-4],因其高能耗特征而常常以牺牲成虫的部分生殖资源为代价[3,5-6],如迁飞过程中消耗掉大量储备能源物质、昆虫种群中迁飞型个体飞行器官发达而产卵量降低和产卵前期延长等[7-8]。但目前也有研究结果表明,昆虫的迁飞与生殖之间不单纯是能量竞争的关系[9-10],一些昆虫在迁飞过程中消耗了能量,但其迁飞行为也的确促进了成虫的生殖,从而使后代种群暴发成灾[11-12]。
草地螟(LoxostegesticticalisLinnaeus)(鳞翅目:草螟科)是一种世界性迁飞害虫,新中国成立以来已有三次大规模暴发成灾,给我国农牧业造成了巨大的经济损失。草地螟幼虫种群的暴发往往出现在成虫大规模的迁飞之后,且通过对迁入地和迁出地成虫量的历史数据分析结果表明,迁入地和迁出地相比往往只需要一半数量的成虫就能引起草地螟后代种群的大发生[13-16]。另外,已有的室内研究结果表明,适宜蛾龄的飞行能增加草地螟雌蛾的产卵同步性[17],这是迁飞后种群暴发的一个重要促进因素,也是揭示草地螟迁飞与种群大暴发的关键问题,而在野外种群中是否也存在这样的促进因素尚未可知,因此本文将通过系统研究野外草地螟迁飞与未迁飞种群成虫生殖行为的差异,明确迁飞对其生殖的影响,从而为正确评价迁飞行为在草地螟后代种群暴发中的作用及地位,提升其预测预报和综合防治水平提供更多的理论依据。
1 材料与方法
1.1虫源及饲养方法
迁飞种群:2014年6月18日(迁入高峰的起始日)到河北省康保县草地螟成虫比较集中的草场地上,用捕虫网捕捉成虫带回室内饲养作为试验材料。
未迁飞种群:2014年7月中旬采集河北省康保县一代幼虫虫茧和内蒙古巴彦淖尔市杭锦后旗5龄末幼虫作为虫源,带回室内待成虫羽化后作为试验材料。
饲养方法:将带回室内的成虫及羽化的成虫均按一雌一雄配对后分别饲养在成虫饲养器具内,温度为(22±1)℃,相对湿度70%±5%,光周期为L∥D=16 h∥8 h,每日以10%的葡萄糖溶液喂养,直至雌雄蛾全部死亡为止。
1.2生殖参数
生殖能力主要通过产卵前期、成虫寿命、产卵历期、产卵量、交配次数、交配率、孵化率和首次产卵历期PFO(period of first oviposition)等参数来衡量,其中迁飞种群的产卵前期确定方法如下:由于试验地6月16日为草地螟始见期,经取样解剖鉴定此时的成虫卵巢多数为Ⅱ级,根据草地螟卵巢发育与日龄的对应关系[18]及Cheng等[17]报道的3日龄为草地螟开始迁飞的日龄,判断当日迁入的草地螟成虫应该多为3日龄,由此可以确定这批越冬代成虫的羽化最早日期为6月14日。结合观察和记录试验中6月18日成虫迁入高峰期捕获的每头雌蛾的具体产卵日期,从而计算出经历迁飞的种群每头雌蛾的产卵前期。由于6月18日捕获的成虫可能有部分是在6月14日以后羽化的,所以理论上本试验计算出的迁飞种群产卵前期应该是最长的,这就更增加了与未迁飞种群较长的产卵前期相比时显著缩短的可信度。成虫寿命也是以6月14日为最早羽化期来确定的,其他生殖参数以及未迁飞种群的产卵前期、产卵历期、产卵量、成虫寿命计算方法参照已有文献报道[19],另外,试验中还引用PFO用于描述一组成虫中所有个体首次产卵的时间同步性,指在一个试验处理内的每头草地螟雌成虫的产卵前期都已知的情况下,该组中的每个产卵前期值减去该组中产卵前期的最小值,则为每头雌成虫所对应的PFO,PFO的值越小,则成虫产卵同步性越高,反之,PFO值越大,成虫产卵同步性越低[17];交配次数是通过雌蛾死亡后解剖并确认其腹部交配囊中的精珠数量获得,精珠个数即为交配次数;交配次数为0的雌雄个体对应的所有生殖参数都将被排除,只在计算交配率被计入总的观测数中;孵化率的计算是随机抽取成虫某日产卵量超过50粒的卵量作为基数,置入6号自封袋中,孵化的幼虫数与卵量基数的比例来获得孵化率,每对成虫只挑选一天符合上述条件的卵量,如果成虫在整个产卵历期中没有符合条件的卵量则该成虫无孵化率数据。
1.3数据统计
所有数据均为平均值±标准误。产卵前期、成虫寿命、产卵历期、产卵量、交配次数、卵的孵化率和PFO的统计分析用t测验(independent two samplesttest)方法,显著水平为α=0.05,其中卵孵化率在统计分析前先进行一次反正弦转化(arcsin)再做后续分析。成虫产卵率和交配率的数据分析均采用卡方检验(χ2)。数据的统计分析在软件SPSS16.0中进行。
2 结果与分析
2.1产卵前期
对野外迁飞和未迁飞草地螟种群的产卵前期进行观察统计(图1),结果表明,迁飞种群的产卵前期平均为5.13 d,未迁飞种群的产卵前期平均为8.70 d,迁飞种群的产卵前期显著短于未迁飞种群(t=-3.58,df=26.13,P=0.001),表明迁飞促进草地螟生殖系统发育导致产卵提前。
2.2其他生殖参数
根据对野外草地螟迁飞和未迁飞种群的其他生殖参数的测定显示(表1),迁飞种群的总产卵量(t=1.50,df=33.90,P=0.143)和卵孵化率(t=-0.22,df=29.27,P=0.829)与未迁飞种群的均无显著差异;而迁飞种群的产卵历期平均为7.40 d,未迁飞种群的平均为10.52 d,相比较迁飞种群的产卵历期显著缩短(t=-3.05,df=32.42,P=0.005);与未迁飞种群相比,迁飞种群的交配次数(t=3.55,df=92.83,P=0.001)显著增多,并且交配率也显著提高(χ2=24.90,df=1,P<0.01)。这表明迁飞对草地螟成虫产卵量影响不显著,但显著增强成虫交配活动的发生及提高其发生频率。

表1 草地螟迁飞与未迁飞种群的其他生殖参数1)Table 1 The other reproductive traits of the migrant and non-migrant populations in Loxostege sticticalis
1) 表中数据为平均数±标准误,括号中的数字表示样本量,*表示在α=0.05水平差异极显著。
Data are presented as mean±SE. Numbers in the brackets represent the sample size. The symbol of *represents it is significantly different at α=0.05 level.
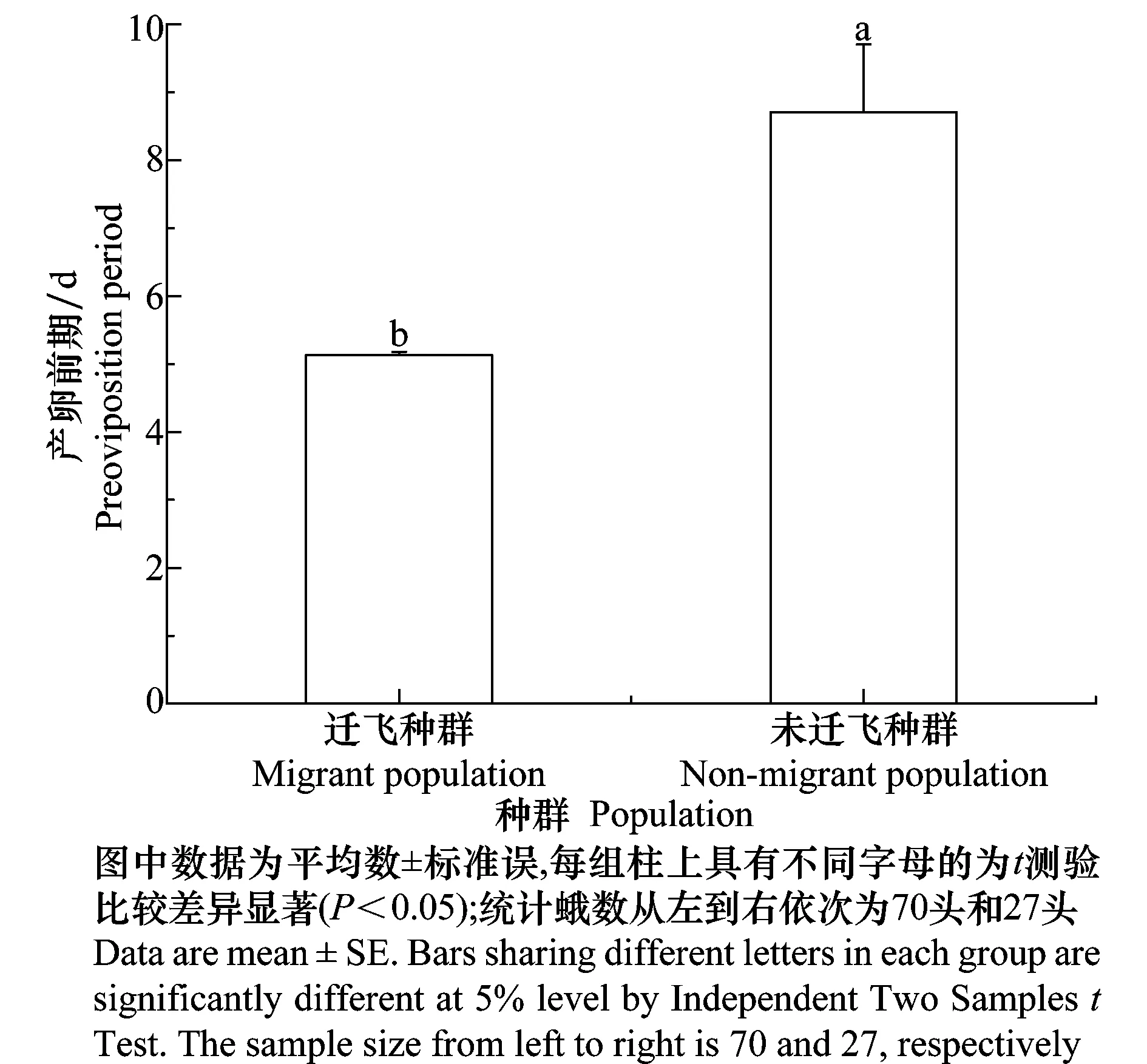
图1 草地螟迁飞与未迁飞种群的产卵前期Fig.1 Preoviposition period of the migrant and non-migrant populations in Loxostege sticticalis
2.3成虫寿命
对野外草地螟成虫寿命的统计分析结果表明(图2),迁飞种群雌蛾的寿命平均为13.30 d,未迁飞种群雌蛾的寿命平均为25.78 d,显著高于迁飞种群的(t=-6.27,df=28.23,P<0.001);迁飞种群雄蛾的寿命平均为15.80 d,未迁飞种群雄蛾的寿命平均为23.07 d,且显著高于迁飞种群的(t=-4.17,df=35.00,P<0.001)。这表明迁飞显著缩短了草地螟雌、雄蛾的寿命。
2.4PFO
对野外迁飞和未迁飞草地螟种群PFO变化的测定结果(图3)显示, 迁飞种群成虫的PFO与未迁飞种群相比平均缩短4.58 d,且显著低于未迁飞种群的(t=-4.58,df=26.13,P<0.001),表明迁飞后的种群产卵起始的同步性显著高于未迁飞种群。另外,未迁飞种群成虫的PFO上四分位数和下四位分数的跨度是4 d,而迁飞种群中两者重叠为一条线,再次证实经历迁飞后成虫之间产卵开始的时间差异性显著缩小。

图2 草地螟迁飞与未迁飞种群的成虫寿命Fig.2 Adult longevity of the migrant and non-migrant populations in Loxostege sticticalis
3 讨论
草地螟幼虫暴发往往紧随成虫的大规模迁飞,迁飞促进种群的暴发已经是一个普遍接受的观点,但迁飞如何导致种群的暴发却始终没有明确的定论,最近的研究结果表明,室内3日龄草地螟成虫经历12 h的飞行后能够显著增加成虫产卵同步性,而对产卵量和其他生殖参数均无显著影响[17]。本文对草地螟野外迁飞种群生殖行为特征的研究表明,迁飞不仅增加了成虫产卵同步性,而且显著缩短了成虫产卵前期、产卵历期和寿命,增加了成虫的交配次数和交配率,这与已有的室内研究结果中飞行对这些生殖参数无显著影响的结果不尽相同。产生这种差异的原因可能有两个方面:第一,迁飞行为本身就是一次自然选择的过程,在迁飞之前及迁飞过程中,不适宜迁飞或不能完成迁飞过程的个体已经被淘汰掉,能够顺利到达迁入地的个体均是被选择出来的相对“优秀”者,与未经迁飞选择的种群相比原本就存在着较高程度的生殖起始和进程的同步性及交配能力的优越性,而室内饲养的种群是没有这样的选择过程的;第二,野外种群的迁飞与室内种群在飞行磨上的被动吊飞相比更能促进草地螟生殖提前、生殖同步性和交配能力增加,这也是为何草地螟室内研究结果中吊飞后的成虫产卵前期、产卵历期和寿命只有缩短的趋势而没有达到统计学上显著水平[17]的主要原因。

图3 草地螟成虫迁飞与未迁飞种群的PFOFig.3 Periods of first oviposition in adults migrant and non-migrant populations of Loxostege sticticalis
昆虫飞行对成虫生殖行为的影响常因其种类的不同而异,如黏虫(Mythimnaseparata)的产卵前期在飞行后有缩短趋势[10],这与草地螟的研究结果类似,但甜菜夜蛾(Spodopteraexigua)的产卵前期不受飞行影响[20]。草地螟飞行后总产卵量和卵孵化率也无显著变化,但褐飞虱(Nilaparvatalugens)飞行后产卵量显著下降[21]。产生上述差异的原因主要与不同昆虫中迁飞行为的适应性有关,如有些昆虫可以通过增加取食补充营养、重新利用飞行肌降解物质[22-26]等方法迅速补充因飞行消耗的物质及能量从而保证后续生殖所需的能量,而不会导致成虫的生殖力下降;另外,飞行过程虽然消耗了能源物质,但是生殖时间的提前和寿命的缩短,客观上都减少了用于生存的能量消耗比例,从而提高了用于生殖的能量消耗比例,这也抵消了飞行过程中的部分能量消耗。
田间调查研究中发现迁入地与迁出地相比往往只需要一半数量的成虫就能够引起草地螟后代种群的大发生[13-16],表明生殖同步性极有可能与迁飞后草地螟种群大暴发紧密相关。首先,草地螟迁入种群的PFO缩短,产卵同步性增加,进而促进了后代卵孵化的同步性和幼虫为害的同步性,即增加了相同生境特定时间段内的种群数量;其次,迁飞行为通常是聚集大量的成虫一起迁飞、运转和降落,当大量成虫集中降落在同一生境,增加了迁入地单位空间内幼虫种群密度急速上升的可能性。综上所述,草地螟迁飞行为引起的成虫产卵同步性在空间和时间上的增加是迁飞行为与种群频繁暴发成灾紧密相连的重要纽带,也是迁飞加剧后代种群暴发的重要原因。
[1]Dingle H. Migration strategies of insects [J]. Science, 1972, 175(4028): 1372-1335.
[2]Dingle H, Drake V A. What is migration?[J]. Bioscience, 2007, 57(2): 113-121.
[3]Rankin M A, Burchsted J C A. The cost of migration in insects[J]. Annual Review of Entomology, 1992, 37(1): 533-559.
[4]Southwood T R E. Habitat, the temple for ecological strategies?[J]. Journal of Animal Ecology, 1977, 46: 337-465.
[5]Dingle H. Animal migration: is there a common migratory syndrome?[J]. Journal of Ornithology,2006,147(2):212-220.
[6]Gunn A, Gatehouse A G. The migration syndrome in the African armyworm moth,Spodopteraexempts: allocation of resources to flight and reproduction [J]. Physiological Entomology, 1993, 18(2): 149-159.
[7]Denno R F, Olmstead K L, McCloud E S.Reproductive cost of flight capability: a comparison of life history traits in wing dimorphic planthoppers[J]. Ecological Entomology, 1989, 14(1): 31-44.
[8]Zhang Y, Wu K, Wyckhuys K A G, et al. Trade-offs between flight and fecundity in the soybean aphid (Hemiptera: Aphididae)[J]. Journal of Economic Entomology, 2009, 102(1): 133-138.
[9]Highnam K C, Haskall P T. The endocrine system of isolated and crowedLocustaandSchistocercain relation to oocyte growth, and the effects of flying upon maturation [J]. Journal of Insect Physiology, 1964, 10(6): 853-864.
[10]罗礼智, 江幸福, 李克斌, 等. 粘虫飞行对生殖及寿命的影响[J]. 昆虫学报, 1999, 42(2): 150-158.
[11]Hanski I, Saastamoinen M, Ovaskainen O. Dispersal-related life-history trade-offs in a butterfly metapopulation [J]. Journal of Animal Ecology, 2006, 75(1): 91-100.
[12]吕伟祥, 江幸福, 张蕾, 等. 一日龄粘虫不同时长吊飞对生殖及寿命的影响[J]. 应用昆虫学报, 2014, 51(4): 914-921.
[13]陈瑞鹿, 暴祥致, 王素云, 等. 草地螟迁飞活动的雷达观测[J]. 植物保护学报, 1992, 19(2): 171-174.
[14]顾成玉, 梁艳春, 张广芝. 草地螟种群数量变动及预测预报技术的研究[J]. 黑龙江农业科学, 1986 (5): 18-21.
[15]魏倩, 崔万里, 杜俊岭, 等. 黑龙江省草地螟发生规律、预测预报及综合防治研究(1981-1985年)[J].中国植保导刊, 1987 (S1): 98-107.
[16]朱世模. 山西中部草地螟发生规律及防治研究[J].山西农业科学, 1963 (2): 16-20.
[17]Cheng Yunxia, Luo Lizhi, Jiang Xingfu, et al. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm [J]. PLoS ONE,2012, 7(2): e31562.
[18]孟正平. 草地螟雌蛾卵巢解剖技术[J]. 中国植保导刊,2007 (12): 28-29.
[19]罗礼智, 李光博. 温度对草地螟成虫产卵和寿命的影响[J]. 昆虫学报, 1993, 36(4): 459-464.
[20]Jiang Xingfu, Luo Lizhi, Sappington T W. Relationship of flight and reproduction in beet armyworm,Spodopteraexigua(Lepidoptera: Noctuidae), a migrant lacking the oogenesIS-flight syndrome [J]. Journal of Insect Physiology, 2010, 56(1): 1631-1637.
[21]沈丽,程遐年.迁飞对褐飞虱生殖的影响[J].南京农业大学学报,1998,21(3):32-35.
[22]李克斌,高希武,罗礼智,等.粘虫飞行过程中四种相关酶的活性变化[J].昆虫学报,2005,48(4):643-647.
[23]李克斌,曹雅忠,罗礼智,等.飞行对粘虫体内甘油酯积累与咽侧体活性的影响[J].昆虫学报,2005,48(2):155-160.
[24]Slansky F J. Food consumption and reproduction as affected by tethered flight in female milkweed bugs (Oncopeltusfasciatus)[J]. Entomologia Experimentalis et Applicata,1980,28(3):277-286.
[25]Willers J L, Schneider J C, Ramaswamy S B. Fecundity, longevity and caloric patterns in femaleHeliothisvirescens: changes with age due to flight supplemental carbohydrate [J]. Journal of Insect Physiology, 1987, 33(1): 803-808.
[26]罗礼智, 李光博, 曹雅忠, 等. 粘虫幼虫密度对成虫飞行与生殖的影响[J].昆虫学报, 1995, 38(1): 38-45.
(责任编辑:田喆)
Effects of migration on reproduction and population outbreak in the beet webworm,Loxostegesticticalis
Song Wenyi1,2,Cheng Yunxia2,Luo Lizhi2,Zhang Lei2,Wu Junxiang1,Jiang Xingfu2
(1. Key Laboratory of Applied Entomology, College of Plant Protection, Northwest A & F University,Yangling712100, China; 2. State Key Laboratory for Biology of Plant Diseases and Insect Pests,Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing100193, China)
In order to further identify the reproductive consequences of insect migration and understand its ecological and evolutionary significance, reproductive traits including period of first oviposition (PFO) and other reproductive parameters of migrant and non-migrant populations were studied systematically. The results showed that preoviposition period, oviposition period, PFO and adult longevity of migrant population were significantly reduced as compared with those of non-migrant population. However, mating frequency and mating percentage were significantly increased, while adult lifetime fecundity and egg hatch rate did not vary significantly. The results indicated that, although adult egg production did not significantly increase, the migrant population could mate for several times quickly and oviposited in advance obviously, and the egg-laying synchrony increased significantly as well, which serve to increase the subsequent larval density and population outbreak.
Loxostegesticticalis;migrant population;unmigrant population;period of first oviposition (PFO)
2015-05-26
2015-05-31
公益性行业(农业)科研专项(201303057,201403031);国家自然科学基金(31301656,31371947);国家国际科技合作专项(2014DFR31250)
E-mail:junxw@nwsuaf.edu.cn;xfjiang@ippcaas.cn
Q 968
A
10.3969/j.issn.0529-1542.2016.02.005
