依据减数分裂相关基因数量判别赫氏球石藻(Emiliania huxleyi)繁殖方式研究
2016-09-08胡永梅杨官品
胡永梅, 郭 栗, 杨官品
依据减数分裂相关基因数量判别赫氏球石藻()繁殖方式研究
胡永梅, 郭 栗, 杨官品
(中国海洋大学 海洋生命学院, 山东 青岛 266003)
繁殖方式是生物重要生物学特征之一。了解生物繁殖方式对遗传和育种研究具有重要意义。赫氏球石藻()呈世界性分布, 在生态系统中扮演重要角色, 但其繁殖方式未知。本研究用分子系统学方法确定有基因组序列且行有性生殖的真核生物的减数分裂相关基因数量, 建立了“减数分裂相关基因≥19个, 其中减数分裂特异基因≥5个”的有性生殖判别标准; 发现赫氏球石藻具有25个减数分裂相关基因, 包括6个减数分裂特异基因。依据建立的标准判定赫氏球石藻行有性生殖。研究结果将有助于赫氏球石藻生活史和遗传学研究。
赫氏球石藻(); 减数分裂; 减数分裂相关基因; 繁殖方式
有性生殖是生物重要的生物学特征之一。已有研究表明有性生殖起源于真核生物演化早期[1-2], 与遗传和育种研究密切相关, 因而备受关注。目前, 有性生殖判定主要有直接记录和间接判别两类方法。直接记录是在显微镜下观察并描述有性生殖过程, 但因有性生殖只发生在生物生活史特定时期, 受环境影响较大, 一般难以记录, 尤其是单细胞生物。间接判定可依赖减数分裂事件、DNA序列和分子标记重组[3-4]、等位基因相异度[5-6]、转座子丰富度等[7]。但这些方法要么耗时太多, 要么跨物种使用通用性低。减数分裂是生物行有性生殖的必要事件, 且广泛存在。Mailk等[8]和Karolina等[9]进行减数分裂相关基因系统学分析, 判别了阴道毛滴虫()和小球藻()的繁殖方式, 同时开发出依据减数分裂相关基因是否存在的生物繁殖方式判别工具。
减数分裂相关基因(包括减数分裂特异基因)是从不同物种生物中鉴别出来的。物种间可能存在“蛋白不同但功能相似、序列差异大无法确定同源性”等情况。因此, 不是每种有性生殖生物都必须具有在不同物种中鉴定出的所有减数分裂相关基因和减数分裂特异基因。Mailk等[8]依据减数分裂相关基因是否存在开发的判别有性生殖工具箱时, 没有设置具体的有性生殖相关基因数量。我们曾设置这一判别数量标准, 并对三角褐指藻等微藻的繁殖方式进行了判别。但因当时已知的有性生殖物种基因组序列数据有限, 设置的“行有性生殖必须具有≥19个减数分裂相关基因, 包括≥6个减数分裂特异基因”的判别数量标准[10-11]需要修改完善。近几年基因组序列越来越丰富, 为修改完善提供了数据基础。
球石藻(coccolithophore)是一类海洋微藻, 属定鞭藻门(Haptophyta)[12], 其生活史特定阶段可形成碳酸钙质地的球石粒(coccolith), 因此得名[13]。赫氏球石藻()是中最受关注并被广泛研究的一种, 1858年在深海沉积物中首次记录, 但其球石粒质地1902年才阐明[14]。从热带到亚极地, 从远洋区到浅海区, 赫氏球石藻都有分布[13]。赫氏球石藻可引发大规模赤潮[15-17], 伴随大规模有机碳沉积[18-19]。赫氏球石藻摄取大气CO2钙化自身, 缓解温室效应, 固定CO2[20-22]。因此, 赫氏球石藻在海洋碳循环中扮演极其重要角色。赫氏球石藻也被认为是方解石最主要的生产者[23]。
赫氏球石藻细胞形态多样, 有C细胞(不运动且覆盖着球石粒)、N细胞(不运动且裸露)和S细胞(运动且覆盖球石粒)[14, 24-26]。S细胞曾被认为是配子[25-26]。这三类细胞都有无性生殖并形成集群的能力。S细胞DNA含量是C细胞的一半[13-14]。人们认为S细胞是单倍体, 而C细胞是二倍体[15], N细胞也是二倍体。赫氏球石藻单倍体和二倍体细胞形态差异明显, 生理学和遗传特性各异[25]。单倍体细胞光敏性较高[24], 不易受感染, 而二倍体细胞易被感染[27]。有人推断可能是减数分裂产生了单倍体细胞, 以抵抗病毒感染, 被比喻为“柴郡猫逃跑策略(Cheshire cat escape strategy)”[27]。单倍体和二倍体细胞转录组也差异显著, 只有不到50%的相同转录本[28-29]。遗憾的是, 赫氏球石藻有性生殖过程从未被记录过, 也没有用其他方法判别过。因此, 赫氏球石藻是否存在有性生殖至今尚未彻底阐明。
本研究用分子系统学方法确定有基因组测序且行有性生殖的真核生物的减数分裂相关基因数量, 首先建立了行有性生殖必须具有的减数分裂相关基因, 包括减数分裂特异基因数的判别标准, 然后判别了赫氏球石藻繁殖方式, 以期促进其生物学和遗传改良研究。
1 材料与方法
Ramesh等[10]罗列了17个减数分裂相关基因。Malik等[8]将减数分裂相关基因扩展到30个, 其中包括9个减数分裂特异基因。本研究: (1)下载了截止2015年7月的减数分裂相关蛋白, 更新了Malik等[8]判别有性生殖工具箱的减数分裂相关蛋白, 重构了这些蛋白的分子系统学树; (2)下载了已知行有性生殖的25个物种的基因组序列, 用本地BLAST工具, 据E-value 2 结果 基于E-value 分析发现赫氏球石藻基因组中共有49个序列完整、具有特征结构域的减数分裂相关基因拷贝。该藻减数分裂相关基因拷贝冗余现象非常突出, 仅次于(16︰24)。经过分子系统学分析, 在赫氏球石藻基因组中共发现25个减数分裂相关基因, 其中包括6个减数分裂特异基因(表1, 图1)。依据本研究建立的判定标准, 赫氏球石藻可进行减数分裂, 营有性生殖。 3 讨论 本研究曾设置“行有性生殖必须具有≥19个减数分裂相关基因, 包括≥6个减数分裂特异基因”的有性生殖判定标准[10-11], 但依据的物种数较少。本研究中, 基于现有已知行有性生殖的物种的基因组, 将这一标准修改为“≥19个相关基因和≥5个特异基因”。由于使用的物种具有了足够宽的代表性, 这一标准应该不会再有改变。减数分裂相关基因(包括减数分裂特异基因)是从不同种生物中鉴别出来的, 而物种间可能存在“蛋白不同但功能相似、序列差异大无法确定同源性”等情况。因此, 这些基因存在与否仅仅依据序列及结构域完整和分子系统学归宿。当真实有性生殖过程难以记录时, 该方法是判定是否存在有性生殖的分子生物学方法之一, 可促进相关物种的基础生物学研究。但最终证实是否存在有性生殖仍需要有性生殖过程描述和其他证据支持。另外, 分子系统学分析确定的有性生殖相关基因或特异基因还需要功能确认。研究发现赫氏球石藻具有30个减数分裂相关基因中的25个, 其中包括9个减数分裂特异基因的6个。依据本研究建立的判定标准, 该藻应具有减数分裂能力, 行有性生殖。 表1 已知存在有性生殖物种和繁殖方式未知的赫氏球石藻基因组中减数分裂相关基因统计 Tab.1 Copy number of meiosis associating genes in known sexually propagating species and 物种总拷贝1冗余拷贝2排除拷贝3相关基因4特异基因 小鼠(Mus musculus)[31]4495309 智人(Homo sapiens)[32]4284309 绿海龟(Chelonia mydas)[33]3973299 剑鱼(Xiphophorus maculatus)[34]4284309 玻璃海鞘(Ciona intestinalis)[35]3214279 热带爪蟾(Xenopus(Silurana) tropicalis)[36]4385309 囊舌虫(Saccoglossus kowalevskii)[37]3725309 原鸡(Gallus gallus)[38]4075288 海葵(Nematostella vectensis)[39]3043237 水螅(Hydra vulgaris)[40]3854299 堡礁海绵(Amphimedon queenslandica)[41]3977258 文昌鱼(Branchiostoma floridae)[42]3152246 丝盘虫(Trichoplax adhaerens)[43]3474236 灰盖鬼伞(Coprinopsis cinerea)[44]3232276 粗糙脉孢菌(Neurosporacrassa)[45]2610255 利什曼虫(Leishmania infantum)[46]2203195 拟南芥(Arabidopsis thaliana)[47]4396288 无油樟(Amborella trichopoda)[48]4286288 可可(Theobroma cacao)[49]4185288 水稻(Oryza sativa)[50]3867256 江南卷柏(Selaginella moellendorffii)[51]51245227 小立碗藓(Physcomitrella patens)[52]3153236 团藻(Volvox carteri)[53]3034235 皱波角叉菜(Chondrus crispus)[54]2832236 莱茵衣藻(Chlamydomonas reinhardtii)[55]2622225 Total---≥19≥5 赫氏球石藻(Emiliania huxleyi)49168256 注:1.氨基酸序列完整, 具有特定结构域的基因拷贝总数;2.系统学分析被确定为相同基因拷贝, 一个计为减数分裂相关基因, 其余记为冗余拷贝;3.系统学分析不能确定为减数分裂相关基因;4.包含特异基因 基因组遗传物质加倍现象广泛存在于自然界中。加倍过程主要通过非依赖机制进行, 这些机制包括DNA复制和重组过程等。在加倍过程中, 遗传物质的加倍程度亦有所区别, 在DNA复制和重组过程中遗传物质发生串联加倍和片段加倍, 而运用其他方法可使整个基因组序列加倍, 从而产生多倍体[56]。其中备受学者关注的是基因加倍。最近相关观点认为基因加倍对植物的多样性具有重要意义, 其关键过程是产生植物适应性进化所必须的原材料[57]。因此减数分裂相关基因的加倍也影响着减数分裂基因的进化和对其基因功能的评估。理论推测加倍基因的进化主要有三种途径: (1)一个拷贝由于退化突变造成基因沉默, 从而成为假基因(即基因的无功能化); (2)一个基因拷贝仍然保持原有功能, 而另一个基因拷贝可能获得一个新的, 有益的功能并由于自然选择而保留下来成为新功能基因(即新功能化); (3)两个基因拷贝可能由于突变的累积, 使得这两个拷贝的功能逐渐退化到二者总的功能相当于其单拷贝祖先基因的水平(即亚功能化)[58]。例如赫氏球石藻存在的2个同源蛋白基因, 经分子系统学分析, 将二者全部排除在减数分裂相关功能之外。这些基因拷贝可能已经演变成为假基因, 也可能正在演变成为新功能基因。由于许多减数分裂基因具有有丝分裂的旁系同源基因(比如,和), 同时这些基因之间拥有着数量众多的相似性序列。但通过分子系统学分析可以清楚的区别出减数分裂相关基因的旁系基因。例如, 通过分子系统学所构建的树形系统树可以将减数分裂相关的同源基因,与无减数分裂功能的同源基因分开, 并将赫氏球石藻中存在的3个同源基因的拷贝区别开, 1个拷贝编码Spo11-2, 另外2个拷贝编码Spo11-3。,等亦存在多个同源基因的拷贝。然而这些拷贝是否具有减数分裂相关功能、具有何种功能, 需进一步的研究。赫氏球石藻中的2个Rad50同源蛋白融入Rad50两个不同的小进化分支中, 这可能是由于在基因加倍的过程中, 同一基因的碱基序列出现差异或是同一基因所编码氨基酸序列出现差异, 也可能是在物种进化过程中两个基因出现趋同进化所造成的。这些推测需要进一步的研究和确定。 A. Msh2, Msh3, Msh4, Msh5和Msh6; B. Mad2和Hop1; C. Rad51和Dmc1, 根置于古菌RadA; D. Mnd1和Hop2; E. Spo11 A. Msh2, Msh3, Msh4, Msh5, and Msh6; B. Mad2 and Hop1; C. Rad51 and Dmc1 rooted into archaeal RadA; D. Mnd1 and Hop2; E. Spo11 虽然现阶段判定有性生殖的方法繁多, 例如显微镜观察, DNA序列和标记重组[3-4]、等位基因相异度[5-6]等, 但仍无法避免实验周期较长, 跨物种使用困难的固有缺点。运用群体遗传学方法, 则需大量个体及多样的地理来源, 在实际运用过程中也存在一定的局限。随着测序技术的发展和完善, 基因组测序变得迅速而高效。依据蛋白序列相似性及结构域保守性等, 通过搜索基因组相关序列可迅速地在最大程度上发现和了解已标识的减数分裂相关蛋白。依据一组减数分裂相关基因(包括减数分裂特异基因), 运用分子系统学分析方法, 可强烈支持有性生殖的存在, 这就提高了赫氏球石藻存在有性生殖的可信度。虽然无法验证这些基因是否具有减数分裂相关功能, 但是可以进一步借助其他分子生物学方法, 例如基因敲除, 遗传互补分析等直接进行验证。 赫氏球石藻具有单、双倍体生活史。Aude等经流式细胞仪分析发现4种球石藻(包括赫氏球石藻)存在两种倍性, 并认为异型球石粒是二倍体向单倍体转换的阶段[24]。本研究的分析结果使赫氏球石藻存在有性生殖的可能性大大提高。有性生殖可有效减少有害突变积累, 有利于物种进化, 可提高物种环境适应性。本研究判定赫氏球石藻具有行有性生殖能力, 但减数分裂和配子融合未直接观察到。因此, 加强对赫氏球石藻有性生殖过程的追踪, 彻底认识其繁殖方式, 不仅是阐明赫氏球石藻完整生活史的需要, 也是其遗传改良研究的需要。 [1] Schurko A M, Logsdon J M. Using a meiosis detection toolkit to investigate ancient asexual scandals and the evolution of sex[J]. Bioessays, 2008, 30(6): 579-589. [2] Ramesh M A, Malik S B, Logsdon J M. A phylogenomic inventory of meiotic genes: Evidence for sex inand an early eukaryotic origin of meiosis[J]. Current Biology, 2005, 15(2): 185-191. [3] Geiser D M, Pitt J I, Taylor J W. Cryptic speciation and recombination in the aflatoxin producing fungus[J]. Proceedings of the National Academy of Sciences, 1998, 95(1): 388-393. [4] Litvintseva A P, Thakur R, Vilgalys R, et al. Multilocus sequence typing reveals three genetic subpopulations ofvar.(serotype A), including a unique population in Botswana[J]. Genetics, 2006, 172(4): 2223-2238. [5] Welch D B M, Meselson M. Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange[J]. Science, 2000, 288(5469): 1211-1215. [6] Birky C W. Heterozygosity, heteromorphy, and phylogenetic trees in asexual eukaryotes[J]. Genetics, 1996, 144(1): 427-437. [7] Wright S, Finnegan D. Genome evolution: Sex and the transposable element[J]. Current Biology, 2001, 11(8): R296-R299. [8] Malik S B, PightlingA W, Stefaniak L M, et al. An expanded inventory of conserved meiotic genes provides evidence for sex in[J]. PLoS ONE, 2008, 3(8): e2879. [9] Fučíková K, Pažoutová M, Rindi F. Meiotic genes and sexual reproduction in the green algal class Trebouxiophyceae (Chlorophyta)[J]. Journal of Phycology, 2015, 51(3): 419-430. [10] Guo L, Yang G. Predicting the reproduction strategies of several microalgae through their genome sequences[J]. Journal of Ocean University of China, 2014, 14(3): 491-502. [11] 郭栗, 杨官品. 依据减数分裂特异基因判定三角褐指藻生殖方式的研究[J]. 中国海洋大学学报(自然科学版), 2013, 43(12): 41-47. [12] Falkowski P G, Knoll A H. Evolution of primary producers in the sea[M]. California: Elsevier Academic Press, 2007: 251-285. [13] Medlin L K, Barker G L A, Campbell L, et al. Genetic characterisation of(Haptophyta)[J]. Journal of Marine Systems, 1996, 9(1): 13-31. [14] Green J C, Course P A, Tarran G A. The life-cycle of: A brief review and a study of relative ploidy levels analysed by flow cytometry[J]. Journal of Marine Systems, 1996, 9(1): 33-44. [15] Laguna R, Romo J, Read B A, et al. Induction of phase variation events in the life cycle of the marine coccolithophorid[J]. Applied and Environmental Microbiology, 2001, 67(9): 3824-3831. [16] Brown C W, Yoder J A. Coccolithophorid blooms in the global ocean[J]. Journal of Geophysical Research, 1994, 99(C4): 7467-7482. [17] Holligan P M, Fernández E, Aiken J, et al. A biogeochemical study of the coccolithophore,, in the North Atlantic[J]. Global Biogeochemical Cycles, 1993, 7(4): 879-900. [18] Westbroek P, De Jong E W, Van Der Wal P, et al. Mechanism of calcification in the marine alga[J]. Philosophical Transactions of the Royal Society of London, 1984, 304(1121): 435-444. [19] Coxall H K, Wilson P A, Pälike H, et al. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean[J]. Nature, 2005, 433(7021): 53-57. [20] Riebesell U, Körtzinger A, Oschlies A. Sensitivities of marine carbon fluxes to ocean change[J]. Proceedings of the National Academy of Sciences, 2009, 106(49): 20602-20609. [21] Gattuso J P, Hansson L. Ocean acidification[M]. New York: Oxford University Press, 2011: 99-121. [22] Westbroek P, Brown C W, Van Bleijswijk J, et al. A model system approach to biological climate forcing: The example of[J]. Global and Planetary Change, 1993, 8(1): 27-46. [23] Robertson J E, Robinson C, Turner D R, et al. The impact of a coccolithophore bloom on oceanic carbon uptake in the northeast Atlantic during summer 1991[J]. Deep Sea Research Part I: Oceanographic Research Papers, 1994, 41(2): 297-314. [24] Houdan A, Billard C, Marie D, et al. Holococcolithophore-heterococcolithophore (Haptophyta) life cycles: Flow cytometric analysis of relative ploidy levels[J]. Systematics and Biodiversity, 2004, 1(4): 453-465. [25] Paasche E, Klaveness D.A physiological comparison of coccolith-forming and naked cells of[J]. Archives of Microbiology, 1970, 73(2): 143-152. [26] Klaveness D.(Lohm.) Kamptn II. The flagellate cell, aberrant cell types, vegetative propagation and life cycles[J]. British Phycological Journal, 1972, 7(3): 309-318. [27] Frada M, Probert I, Allen M J, et al. The Cheshire Cat escape strategy of the coccolithophorein response to viral infection[J]. Proceedings of the National Academy of Sciences, 2008, 105(41): 15944- 15949. [28] Rokitta S D, De Nooijer L J, Trimborn S, et al. Transcriptome analyses reveal differential gene expression patterns between the life-cycle stages of(Haptophyta) and reflect specialization to different ecological niches[J]. Journal of phycology, 2011, 47(4): 829-838. [29] Von Dassow P, Ogata H, Probert I, et al. Transcriptome analysis of functional differentiation between haploid and diploid cells of, a globally significant photosynthetic calcifying cell[J]. Genome Biology, 2009, 10(10): R114. [30] Read B A, Kegel J, Klute M J, et al. Pan genome of the phytoplanktonunderpins its global distribution[J]. Nature, 2013, 499(7457): 209-213. [31] Chinwalla A T, Cook L L, Delehaunty K D, et al. Initial sequencing and comparative analysis of the mouse genome[J]. Nature, 2002, 420(6915): 520-562. [32] Lander E S, Linton L M, Birren B, et al. Initial sequencing and analysis of the human genome[J]. Nature, 2001, 409(6822): 860-921. [33] Wang Z, Pascual-Anaya J, Zadissa A, et al. The draft genomes of softshell turtle and green sea turtle yield insights into the development and evolution of the turtlespecific body plan[J]. Nature Genetics, 2013, 45(6): 701-706. [34] Schartl M, Walter R B, Shen Y, et al. The genome of the platyfish,, provides insights into evolutionary adaptation and several complex traits[J]. Nature Genetics, 2013, 45(5): 567-572. [35] Dehal P, Satou Y, Campbell R K, et al. The draft genome of: Insights into chordate and vertebrate origins[J]. Science, 2002, 298(5601): 2157-2167. [36] Hellsten U, Harland R M, Gilchrist M J, et al. The genome ofthe western clawed frog[J]. Science, 2010, 328(5978): 633-636. [37] Lowe T M, Eddy S R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence[J]. Nucleic Acids Research, 1997, 25(5): 955-964. [38] Hiller L W, Miller W, Birney E, et al. Sequence and comparative analysis ofthe chicken genome provide uniqueperspectives on vertebrate evolution[J]. Nature, 2004, 432(7018): 695-716. [39] Putnam N H, Srivastava M, Hellsten U, et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization[J]. Science, 2007, 317(5834): 86-94. [40] Chapman J A, Kirkness E F, Simakov O, et al. The dynamic genome of[J]. Nature, 2010, 464(7288): 592-596. [41] Srivastava M, Simakov O, Chapman J, et al. Thegenomeand the evolution of animal complexity[J]. Nature, 2010, 466(7307): 720- 726. [42] Putnam N H, Butts T, Ferrier D E K, et al. The amphioxus genome and the evolutionof the chordate karyotype[J]. Nature, 2008, 453(7198): 1064-1071. [43] Srivastava M, Begovic E, Chapman J, et al. Thegenome and the nature of placozoans[J]. Nature, 2008, 454(7207): 955-960. [44] Stajich J E, Wilke S K, Ahrén D, et al. Insights into evolution of multicellular fungi from theassembled chromosomes of the mushroom()[J]. Proceedings of the National Academy of Sciences, 2010, 107(26): 11889-11894. [45] Galagan J E, Calvo S E, Borkovich K A, et al. The genome sequence of the filamentous fungus[J]. Nature, 2003, 422(6934): 859-868. [46] Peacock C S, Seeger K, Harris D, et al. Comparative genomic analysis of threespecies that cause diverse human disease[J]. Nature Genetics, 2007, 39(7): 839-847. [47] Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant[J]. Nature, 2000, 408(6814): 796-815. [48] Amborella Genome Project. Thegenome and the evolution of flowering plants[J]. Science, 2013, 342(6165). [49] Argout X, Salse J, Aury J M, et al. The genome of[J]. Nature Genetics, 2011, 43(2): 101- 108. [50] Goff S A, Ricke D, Lan T H, et al. A draft sequence of the rice genome (L. ssp.)[J]. Science, 2002, 296(5565): 92-100. [51] Banks J A, Nishiyama T, Hasebe M, et al. Thegenome identifies genetic changes associated with the evolution of vascular plants[J]. Science, 2011, 332(6032): 960-963. [52] Rensing S A, Lang D, Zimmer A D, et al. Thegenome reveals evolutionary insights into the conquest of land by plants[J]. Science, 2008, 319(5859): 64-69. [53] Prochnik S E, Umen J, Nedelcu A M, et al.Genomic analysis of organismal complexity in the multicellular green alga[J]. Science, 2010, 329(5988): 223-226. [54] Collén J, Procel B, Carré W, et al. Genome structure and metabolic features in the red seaweedshed light on evolution of the Archaeplastida[J]. Proceedings of the National Academy of Sciences, 2013, 13(110): 5247-5252. [55] Merchant S S, Prochnik S E, Vallon O, et al.Thegenome reveals the evolution of key animal and plant functions[J]. Science, 2007, 318(5848): 245-250. [56] Ramsey J, Schemske D W. Pathways, mechanisms, and rates of polyploid formation in flowering plants[J]. Annual Review of Ecology and Systematics, 1998, 29: 467-501. [57] Flagel L E, Wendel J F. Gene duplication and evolutionary novelty in plants[J]. New Phytologist, 2009, 183(3): 557-564. [58] Lynch M, Conery J S. The evolutionary fate and consequences of duplicate genes[J]. Science, 2000, 290(5494): 1151-1155. (本文编辑: 康亦兼) Ascertaining the reproduction strategy ofby numbering its meiosis-associated genes HU Yong-mei, GUO Li, YANG Guan-pin (College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China) Reproduction strategy is one of the most important biological characteristics of an organism. Ascertaining such a strategy is crucial for its genetic research and improvement.plays a key role in the ocean ecosystem, but nothing is known about its reproductive strategy. Here, we numbered the meiosis-associated genes of sexually reproductive eukaryotes with genome sequences that were available and found that they held ≥19 meiosis-associated genes, of which ≥5 were meiosis-specific. We also found thatcontained 25 meiosis-associated genes, of which 6 were meiosis-specific. Based on this information, we concluded thatperforms sexual reproduction. This ascertainment may aid to studying its life cycle and genome and further its genetic improvement. Nov.17, 2015 ; meiosis; meiosis-associated gene; reproductive strategy Q178.53 A 1000-3096(2016)06-0001-07 10.11759/hykx20151117002 2015-11-17; 2016-02-05 国家自然科学基金(3127408) [Foundation: National Natural Science Foundation of China, No.3127408] 胡永梅(1989-), 女, 山东青岛人, 硕士研究生, 主要从事海洋微藻研究, 电话: 18561538935, E-mail: xiaoping0331hu@163.com; 杨官品, 通信作者, 教授, 博士生导师, 电话: 0532-82031636, E-mail: yguanpin@mail.ouc.edu.cn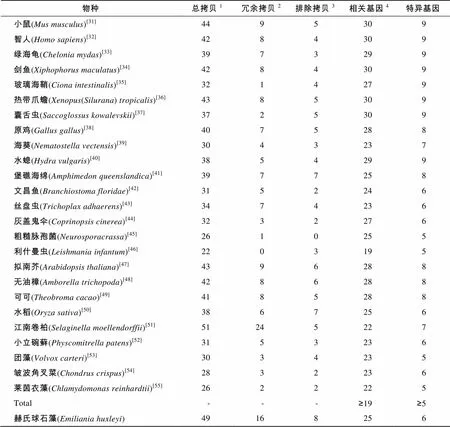
Guo L, Yang Guanpin. Study on the determination ofreproduction strategy with the presence of meiosis-specific genes[J]. Periodical of Ocean University of China, 2013, 43(12): 41-47.
