富营养化湖泊中的鲢、鳙控藻问题:争议与共识*
2016-09-01刘其根
刘其根,张 真
(上海海洋大学水产与生命学院,上海 201306)
富营养化湖泊中的鲢、鳙控藻问题:争议与共识*
刘其根,张真
(上海海洋大学水产与生命学院,上海 201306)
我国湖泊富营养化的治理方向正在从污染控制走向湖泊的生态修复与管理,因此以鲢、鳙为主导的非经典生物操纵技术受到了关注和重视. 然而,鲢、鳙控藻的研究有成败两方面的案例,导致人们对能否利用鲢、鳙控制富营养化湖泊中藻类的过度增长的看法仍有分歧. 本文通过对国内大量相关研究案例进行剖析,指出了导致鲢、鳙控藻试验产生不同结果的可能原因,提出了对鲢、鳙控藻研究可以得到的共识,以便为我国湖泊环境的管理和保护提供借鉴.
湖泊;富营养化;鲢;鳙;藻类过度增长;藻类控制
鲢和鳙是我国最常见、也是世界上养殖产量最大的两种淡水经济鱼类[1]. 鲢、鳙、青鱼和草鱼被并称为“四大家鱼”,在我国的淡水渔业发展史上具有举足轻重的地位. 鲢和鳙,不但曾是我国池塘的主要养殖鱼类,也被广泛用于全国各地湖泊(含水库,下同)的增殖放流,其在一些湖泊的渔产量中常可占到40%以上(如鲢、鳙在浙江千岛湖占到全湖渔产量的50%以上,而在武汉东湖占到90%以上[2]). 因此鲢、鳙无疑是我国湖泊中最具影响的优势鱼类类群,也是我国湖泊食物网结构区别于国外湖泊的最显著特征[3].
然而,人们对鲢、鳙的兴趣并不只限于其渔业利用价值. 由于鲢、鳙主要以浮游生物为食,因而利用鲢、鳙控制富营养化湖泊中藻类的过度增长或蓝藻水华的可能性也早已引起了国内外的广泛兴趣[4-6]. 利用鲢、鳙控藻的这一技术现在也都以“非经典生物操纵”而广为人们所熟知[7],然而人们对鲢、鳙能否控藻的认识却始终存有争议,至今未达成共识. 实际上,关于鲢、鳙生态学效应或影响的国内外研究可谓不少,但令人遗憾的是,这些研究结果之间常相互矛盾,不但使开展这些研究的学者们对其认识不能达成统一,也会使广大读者对这些结果感到莫衷一是,致使对鲢、鳙控藻的意见分歧在国内外都长期和广泛存在. 这些分歧意见的存在,或可使鲢、鳙对水环境的积极作用得不到应有的应用或使其负面作用得不到有效规避,或使一些不当的水环境管理政策被推出. 因此尽早对这个问题进行明确解答或形成共识,对我国的水环境保护具有重要的意义.
为了帮助人们解答“鲢、鳙究竟能否控藻”的疑惑,笔者通过对国内外大量相关研究的分析提出并尝试回答以下几个问题:为什么不同的研究者得到的结果会如此迥异甚至完全对立?对现有众多的鲢、鳙控藻研究进行分析总结,能否达成共识?如果现有的证据还不足以形成共识,那么今后应开展怎样的研究才能有助于问题(分歧)的真正解决?
1 鲢、鳙控藻问题的提出及其界定
利用鲢、鳙控制富营养化湖泊中藻类的过度增长,是在湖泊富营养化问题日趋普遍之后才被提出并逐渐受到关注的. 早在1970s-1980s,国际上就已有对鲢、鳙控藻的研究[4,8-10],我国也在1980s开始关注和研究鲢、鳙控藻的问题[11-12]. 藻类的过度增长,特别是蓝藻水华的暴发,常被认为是湖泊富营养化的最恶劣表征,不但严重影响湖泊水质,也会对湖泊生态系统的健康产生影响,因而是湖泊富营养化控制的一个重要方面. 由于鲢、鳙能够滤食藻类,因此能否利用鲢、鳙来控制富营养化湖泊中的藻类过度增长问题自然就成为了国内外研究者极感兴趣的一个议题.
本文所要探讨的鲢、鳙控藻问题,是指在污染源已得到基本控制(即点源污染已得到有效控制、非点源和内源污染将是主要影响因素)的湖泊,能否利用鲢、鳙来控制藻类的过度增长,或预防控制蓝藻水华暴发的问题. 即探讨鲢、鳙能否控藻时,不应包括那些连点源污染都尚未得到有效控制的湖泊,因为在没有控制污染源的湖泊,仅通过鲢、鳙放养来控制其藻类的过度增长是有难度的. 现有的共识是:污染源的控制是富营养化防治的前提. 而在污染源已得到基本控制的湖泊,之所以仍需要借助鲢、鳙来控藻,是因为即使点源污染得到较好的控制,其水质也并非一定能得到及时的改善,相反,这些湖泊仍可能会因为来自流域的大量面源污染或湖泊自身的内源性污染,导致藻类出现异常增长,甚至暴发蓝藻水华,因此借助生物操纵等措施来预防控制藻类的过度增长和改善水质仍可能是非常必要的. 这也是我国湖泊利用鲢、鳙控藻的主要应用场景. 即鲢、鳙控藻应是湖泊环境管理的一种手段,是对外部污染控制的有效补充.
2 鲢、鳙控藻试验研究的典型案例
有关鲢、鳙控藻的研究,国内外都有大量的案例. 谢平的《鲢、鳙与藻类水华控制》[7]已给出了国内外的很多研究案例,有兴趣的读者可以参考. 然而,由于这些案例还不足以消除人们对鲢、鳙能否控藻的意见分歧,故本文将给出更多的研究案例,旨在帮助读者进一步了解鲢、鳙控藻研究的现状和进展,以帮助人们尽早达成对鲢、鳙控藻的共识. 由于鲢、鳙控藻有成败两方面的案例,因此解决对鲢、鳙控藻的分歧,也必须从正反两方面的案例研究入手.
2.1鲢、鳙控藻的失败案例
大连水产学院史为良教授及其同事的研究,是国内最早开展的鲢、鳙控藻试验之一. 该研究于1982年5-7月进行[11],利用水库岸边的8个3 m3(面积约2 m2,深1.5m)的水泥池,分A、B两组同时进行. A组池底加入半寸左右厚度的水库底泥,B组不加. 试验池内注入库水,分别以0、6、18和36尾/m3的密度放养平均全长10.2cm的鲢、鳙鱼种(鲢、鳙之比为2∶1). 同时在水库库区同一网箱养殖区域选定10个相邻网箱,分别以每箱(规格7 m×4 m×1.8m)0、0.6、1.0和1.5万尾的密度放养全长10 cm左右的鲢、鳙鱼种,分C、D两组进行试验对照. 每5 d测定1次水化学和水生生物组成指标. 每次采样时间为15:00-15:30. 网箱试验期间不洗箱,水流交换很弱,微量测流仪已测不出流速. 网箱试验前、后各进行1次水化学和水生生物组成指标的测定. 试验周期(在水泥池和网箱)均为20 d. 研究结果表明,放养鲢、鳙使水体初级产量、浮游植物生物量和P/B系数大幅度上升. 其中,密养池变化幅度大于稀养池,A组强于B组. 同样,鱼种密度较高的网箱内,浮游植物生物量、生产量和P/B系数等均未因鲢、鳙摄食而相应减少,附着藻类却明显比未养鱼的空箱多,且箱内浮游植物生物量、生产量和P/B系数也稍高于箱外. 此外,随着放养密度达到一定量后,水体中的浮游植物明显小型化,优势种也发生了明显的变化. 以A组为例,未养鱼的A1号池,浮游植物的细胞数为140×104cells/mg,而在放养36尾/m3的A2号池,浮游植物的数量则达到2650×104cells/mg. 随着放养密度的增加,浮游动物生物量下降,且也出现小型化. 而鱼类生长随密度增加而变差. 实验结果表明,鲢、鳙不能控制水体中的藻类数量.
此后,中国科学院水生生物研究所的阮景荣研究员及其同事于1994年前后也在实验室水族箱条件下开展了鲢、鳙控藻的研究[13]. 他们选用了12个60 L的水族箱,分3个实验组(Ⅱ组为鲢鳙混养,Ⅲ组为鲢单养,Ⅳ组为鳙单养)和1个对照组(Ⅰ组),每组设3个重复,水族箱添加以活性炭过滤过的自来水配置的一种被称为WC的藻类培养基,水深为31 cm,并接种罗非鱼实验留下的混合藻类培养液,接种的初始密度为0.15×105cells/L,接种的藻类有颤藻(Oscillatoriasp.)、镰形纤维藻(Ankistrodesmusfalcatus)、菱形藻(Nitzschiasp.)、栅藻(Scenedesmussp.)、小球藻(Chlorellasp.)、衣藻(Chlamydomonassp.)等,其中颤藻占80%以上. 藻类接种后第3周,引入大型溞(Daphniamagna),密度为每个水族箱22个成体. 在接种大型溞后第4周再放养规格分别为2.5~3.5cm的鲢、鳙鱼种,放养密度平均为15 g/m3. 其中两组为鲢和鳙单养,两组为鲢和鳙混养,实验共持续27周,其中鲢、鳙放养后实验持续了21周. 为了能使接种藻类生长,每个水族箱都配备了5000~6000 lx的光照,光照时间为12 h/d. 水族箱水温控制在25℃左右,每周采样1次用于水生生物和水化学指标的测定,每周于采样后给各水族箱补充等量的营养物和自来水,其营养物补充量按照WC培养基用量的1%~5%逐渐增加,实验期间的平均磷负荷为0.0061 g/(m3·d). 实验结果表明,鲢、鳙引入微型实验生态系统后,大型溞密度降低,而浮游植物密度增高,其消长幅度以单养鳙组最高,同时,浮游植物的组成亦发生了很大的变化,蓝藻和硅藻所占比重显著下降,而绿藻的相对密度有大幅度增长. 鲢、鳙放养期间,各实验组的总初级生产力以及II、IV组浮游植物初级生产力都显著高于对照组(P<0.001),表明鲢、鳙不能控制藻类生物量的增长.
由于鳙常被认为主要摄食浮游动物,因此放养鳙将导致浮游动物小型化,基于经典生物操纵理论,放养鳙将不但不能控藻,而且可能加速藻类特别是蓝藻的增长. 因此鳙控藻不成功的结果也似乎更容易被人们所接受. 然而,多项“以鲢控藻”的试验,也没有显示出成功的控藻结果.
唐汇娟等[14]在武汉东湖水果湖湾开展了一个围隔试验,围隔采用不透水的聚乙稀塑料布缝合成袋状,底部被封死并固定于钢管结构上,其结构为2.5m×2.5m的正方形,围隔中水深2 m. 用抽水泵向围隔中注入附近的湖水,随后将在附近收集到的东湖底泥加入围隔中,使各个围隔中底泥厚度达到5 cm. 待底泥沉积1夜后,在围隔中放入鲢. 试验用鱼为1龄鱼,平均体长为19.1±3.3cm,试验前已在湖边驯养. 共设4个不同的鱼密度:0、116、176和316 g/m2,即低密度、中密度、高密度和空白对照,每个处理设2个重复. 试验在2006年5-6月间进行,持续了2个月. 试验结果为:(1)氮、磷浓度方面,围隔氨氮浓度低于湖水且各围隔氨氮浓度无显著差异,无鱼围隔正磷酸盐浓度显著高于有鱼围隔和湖水,但3个有鱼围隔,以中密度组的磷浓度最高,且显著高于高密度组围隔. (2)透明度方面,则为围隔高于湖水,无鱼围隔高于有鱼围隔,有鱼围隔水体透明度又随鱼的密度而递增. (3)虽然有鱼围隔中,浮游植物生物量与鱼的密度呈反比,即随鱼密度增加而减少,但有鱼围隔的浮游植物生物量却大于无鱼围隔,表明,以鲢控藻的实验并没有成功. 研究者认为,造成该实验结果与以往围隔能够控藻的实验的差别主要有两方面原因:一是该实验与以往实验的季节不同,其他围隔实验多发生在夏季蓝藻暴发季节,使得无鱼围隔更易暴发蓝藻;二是可能无鱼围隔中存在较多的大型溞(但实验中实际是透明溞(D.hyalina)),在一定程度上抑制了蓝藻水华的发生.
而更近一次显示“鲢不能控藻”的围隔试验是由暨南大学在广东省从化市流溪河水库进行的[15]. 该水库位于热带与亚热带分界线(北回归线)附近,其气候和水文条件可能与长江流域水库有一定的差异. 流溪河水库是一座典型的峡(山)谷型水库,最大水深73 m,平均水深21.3m. 水库面积15.25 km2,库容3.25×108m3,为一贫中营养型水库. 控藻实验在24个围隔中进行,每个围隔的体积(长×宽×深)为4 m×4 m×6 m. 围隔底部封闭,上口敞开. 围隔全部固定在由铁桶和铁架所构成的浮床上,围隔离岸约50 m,为防止围隔内鱼类逃逸和水库内鱼类进入,围隔顶部上方均高出水面约50 cm,围隔内实际水体体积约为85 m3. 该实验设置对照组、添加不同营养盐浓度梯度组(设低浓度营养盐、中浓度营养盐、高浓度营养盐3个梯度)与添加不同营养盐浓度梯度+鱼(低浓度营养盐+鱼、中浓度营养盐+鱼、高浓度营养盐+鱼)等共7个处理组,每个处理组设3个平行. 实验添加的营养盐为NH4NO3和KH2PO4. 3个浓度梯度的N和P浓度分别为:低浓度组0.3、0.03 mg/L;中浓度组0.6、0.06 mg/L;高浓度组0.9、0.09 mg/L. 营养盐添加时先用围隔内的水使其充分溶解,然后将其均匀地泼洒入围隔内、并充分搅拌围隔内的水使其均匀分布于围隔内的水体中. 实验用鲢的规格为1龄鱼种,其平均体长约为16.5cm/尾,体重约为80 g/尾. 围隔中鲢的放养密度为4 g/m3. 实验于2008年4月24日正式开始,至6月7日结束. 实验结果显示,有鱼围隔透明度小于无鱼围隔,总氮(TN)浓度低于无鱼围隔,总磷(TP)浓度则高于无鱼围隔;有鱼围隔的叶绿素a浓度也高于无鱼围隔,且鲢使围隔内的蓝藻、绿藻、硅藻和隐藻生物量升高,鲢还使围隔内藻类小型化,使<10 μm的藻类生物量明显升高. 总之该实验的结果似乎也说明了,放养鲢使围隔藻类生物量不降反升,用鲢控藻是不可行的.
除了这些实验研究外,Wang等[16]还通过对我国长江中下游的35个浅水湖泊和10个池塘的大规模调查,探讨了鲢、鳙与藻类的关系. 调查发现,叶绿素a浓度在鲢、鳙产量超过100 kg/hm2的湖泊显著高于鲢、鳙产量低于100 kg/hm2的湖泊,但TP浓度与叶绿素a浓度或TP浓度与透明度的关系在这两类湖泊中并没有显著差异,据此得出结论:鲢、鳙不能降低叶绿素a浓度或者提高水体透明度,故不宜作为旨在改善水质的生物操纵之用.
对鲢、鳙不能控藻的证明,还来自一些间接证据. 王银平等[17]利用原位实验调查了鲢、鳙粪便中的藻类是否具有光合活性. 其研究方法是:在中国科学院南京地理与湖泊研究所太湖湖泊生态系统研究站设置规格为1 m×1 m×1.5m的敞口式聚乙烯不透水围隔9个(鲢组、鳙组和对照组各3个),加入经200 μm筛网预滤的湖水和鲢、鳙. 所用鲢规格为体重84.8±2.3g,体长17.7±1.2cm,鳙规格为体重76.6±1.7g,体长17.2±1.5cm(均购自中国水产科学研究院淡水渔业研究中心). 实验前对这些鱼类进行7~10 d的微囊藻驯养. 实验开始时,将饥饿72 h的健壮鲢、鳙分别放入微囊藻水华严重的围隔中,每组10条,对照组不放鱼. 待排泄稳定,分别收集围隔内漂浮的条状排泄物,用去离子水缓缓冲洗排泄物表面附着物后转入盛有经0.2μm滤膜预滤湖水的锥形瓶中超声振荡, 打匀,随后等量移取至透析袋中(截流分子量14 kD,半周长150 mm),然后将透析袋分别悬挂在对应围隔中进行原位渗透培养. 对照组透析袋中加入未被摄食的经超声震荡的微囊藻悬浮液. 实验期间,每天定时摇动透析袋4次,每2 d取样测定藻类叶绿素荧光参数、叶绿素a和胞外多糖浓度、藻细胞密度,培养周期13 d. 于实验第1、7、13 d取样进行藻种鉴定,并利用德国Walz公司生产的双通道PAM-100荧光仪,对藻类进行叶绿素荧光参数测定. 研究结果发现,微囊藻经鲢、鳙滤食后,虽然其在排出鲢、鳙消化道后的第一天,其叶绿素荧光参数PSⅡ最大光能转化效率(Fv/Fm)、PSⅡ潜在光合活性(Fv/Fo)、PSⅡ实际光能转化效率(Yield)和光合电子传递速率(ETR)均显著低于对照组(P<0.05),而光化学猝灭(qP)和非光化学猝灭(NPQ)显著高于对照组(P<0.05),但随着培养的推进,其光合活性的参数迅速升高,至实验结束时,鲢、鳙组的Fv/Fm、Fv/Fo、Yield和qP均显著高于对照组,其非光化学猝灭(NPQ)又显著低于对照组(P<0.01). 原位培养期间,鲢、鳙组藻细胞密度和叶绿素a浓度呈增长趋势,且鲢组明显高于鳙组;鳙组藻类游离胞外多糖含量增长幅度高于鲢组. 至实验结束时,鲢、鳙组浮游藻类总生物量分别为对照组的7.78、6.55倍,据此他们认为鲢、鳙单次滤食未对微囊藻造成生理上的致命损伤,而藻类由于超补偿生长,其光合及生长活性在短期恢复并显著增强,有潜在加速水体富营养化的可能,利用鲢、鳙控藻的技术值得商榷.
2.2鲢、鳙控藻的成功案例
2.2.1池塘和池塘围隔中的控藻试验
案例一:美国斑点叉尾鮰养殖池塘中的控藻研究
斑点叉尾鮰是美国最主要的养殖鱼类,年产量约2800 t,年销售收入逾3.5亿美元. 由于传统单养池塘中经常出现微囊藻等蓝藻水华,不但恶化水质,还会使养殖的斑点叉尾鮰出现异味,因此研究其池塘控藻技术具有重要意义.
为了解决斑点叉尾鮰池塘的控藻问题,Mueller等[18]于2000年8月30日至9月30日开展了该项控藻试验. 研究选用了4套池塘养殖系统,每套系统的结构完全相同,均由4部分组成:水车式增氧机、藻类区、控藻区和养殖区. 其中斑点叉尾鮰主要放养在养殖区,4套系统中斑点叉尾鮰的养殖密度完全相同,均为15500 kg/hm2. 池塘废水经水车式增氧机驱动流入藻类区净化. 藻类区面积最大,占整个养殖系统的95%,藻类区藻类吸收养殖废水中的氮、磷得到增殖,经水流驱动,流入控藻区. 控藻区放养滤食生物(鲢Hypophthalmichthysmolitrix或蚌Elliptiocomplanata)滤食藻类,4套养殖系统的主要区别就在于控藻区放养的滤食生物种类不同. 为了了解不同控藻生物的控藻效果,该研究选择了在鲢和蚌之间的不同轮换放养,如养殖系统A中,在8月30日至9月1日期间控藻区放养鲢,鲢的起始密度为3000 kg/hm2,然后从9月2日至30日开始放养蚌,其初始密度为3300 kg/hm2(组织湿重). 各养殖系统中滤食生物的具体设置如表1所示.
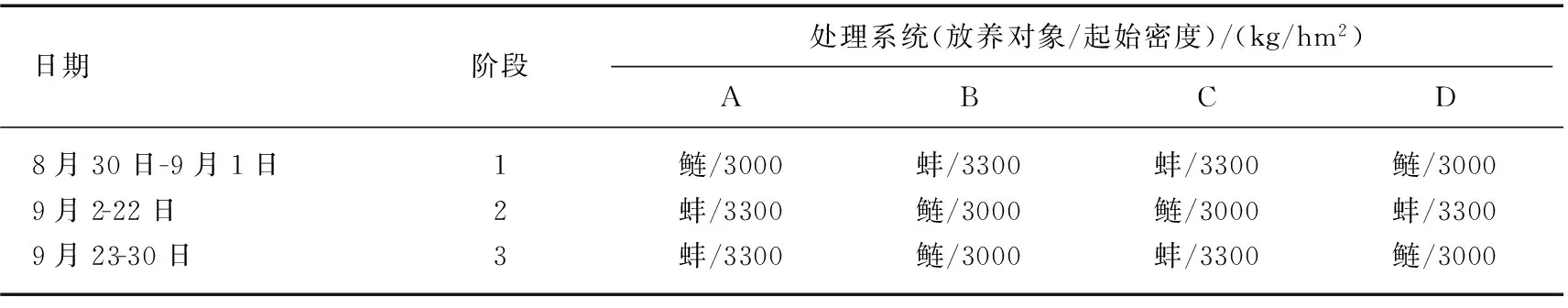
表1 斑点叉尾鮰养殖试验控藻区滤食生物的放养处理设计
试验期间,用蚌和鲢处理的池塘水质监测结果分别列于表2. 从各项水质指标来看,在以鲢或蚌作为滤食生物进行控藻时各项水质参数之间差别不明显.
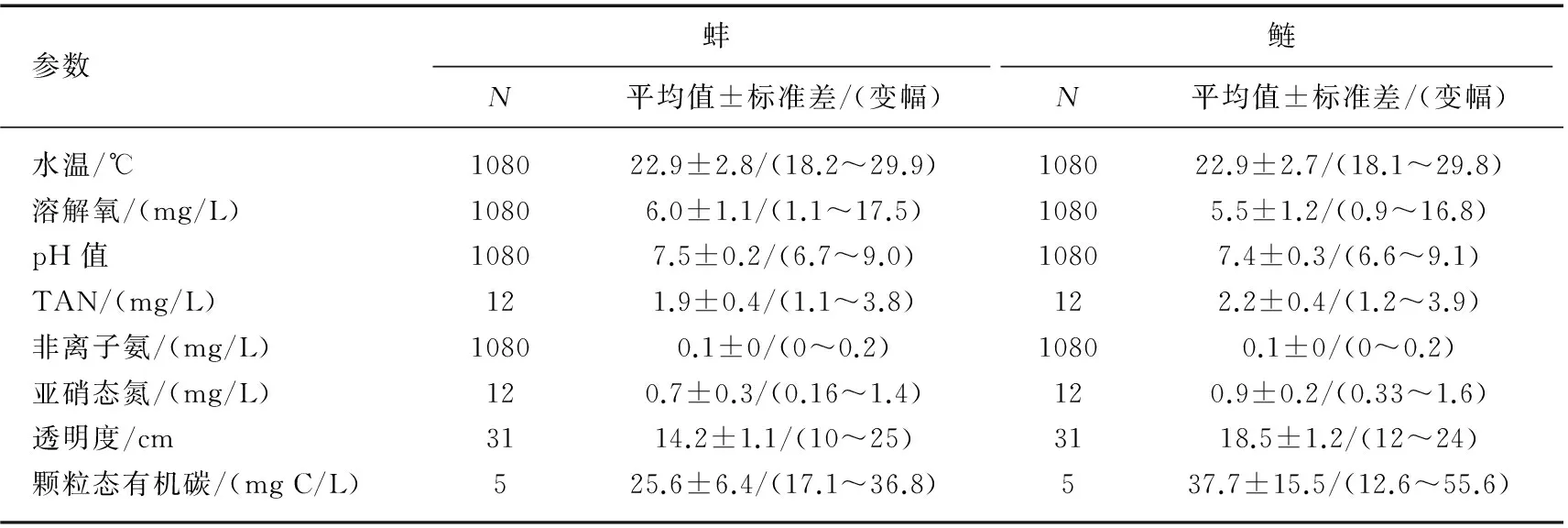
表2 试验期间养蚌和鲢水体的主要水质指标
然而,用蚌或鲢做控藻生物时,各养殖系统中的藻类组成差别显著. 例如,在试验第1阶段,以蚌为滤食生物的B、C两个系统,其藻类组成均以微囊藻为单一优势种,其藻类密度(单位:cells/ml)占61.6%,其生物体积(个体平均体积×生物密度)占比达99.9%. 所形成的微囊藻群体的平均颗粒大小达78.5±25.1μm(以最大轴直径测量,GALD). 而以鲢为滤食生物的A和D系统中,藻类组成仅有绿藻而没有蓝藻,其优势种为栅藻、纤维藻和一种未定名圆盘状藻类. 3种优势藻类的大小分别为10.0±2.4、3.0±0.5和4.0±2.0 μm GALD. 至试验阶段2,在将养殖系统A和D中的鲢替换成蚌后,很快微囊藻便发展成为优势种,微囊藻的生物体积以平均6.4%±11.8% d-1的速度增加,尽管微囊藻的生物密度在此阶段增加,但其增长的斜率与阶段1差异不大. 在这两个系统中由于滤食生物由鲢变为蚌以后,微囊藻群体的大小也从47.9±7.2μm增加到了77.9±15.9μm GALD. 由鲢改为蚌后,栅藻的大小也增加了,与阶段1相比,增加的回归曲线斜率也没有呈现显著差异. 而在此阶段,将B和C系统中的蚌滤食改为鲢滤食后,原先的微囊藻数量出现了显著减少,其生物体积以平均3.9%±6.3% d-1的速度减少. 虽然微囊藻数量减少,但其变化的斜率与阶段1没有显著差异. 微囊藻群体的平均颗粒大小在阶段1和2之间差异显著,阶段1平均为44.5±18.9μm,阶段2平均为33.2±4.1μm GALD. 栅藻的平均大小也减小了,但减小的斜率差异不显著. 在滤食生物从蚌变为鲢后两周,小于6 μm的小型藻类开始出现,颗粒态有机碳也有一定程度的减少. 至阶段3,将D系统的蚌再改为鲢后,其微囊藻密度随之降低,其生物体积比也显著降低(P=0.003),乃至彻底消失. 而在A系统中,由于仍保留了蚌,因此其微囊藻仍是优势种,其颗粒大小也比D系统中的大(A系统中微囊藻颗粒大小为64.9±18.4μm GALD,而D系统中为50.2±13.7μm GALD). 而在系统C中,当将滤食生物由鲢再变回蚌后,微囊藻生物密度增加,其生物体积更是出现了显著的增加,而在系统B中,由于鲢仍是滤食者,微囊藻彻底消失.
该研究清楚地表明,鲢为滤食生物都能导致微囊藻的迅速降低并最终消失,而系统中没有鲢后都会出现微囊藻,说明鲢可以控制池塘中的微囊藻. 同样是滤食,蚌由于摄食的颗粒粒径小,因此不但不能控制蓝藻,反而会促进蓝藻的增长.
案例二:浙江金华养蚌池的围隔控藻试验
笔者实验室于2007年6月19日至12月25日在浙江省金华市汤溪镇浙江金华威望养殖公司养殖基地开展了该项鲢、鳙控藻试验[19],试验在设置于一4 hm2左右的大型养蚌池的9个围隔中进行. 每个围隔面积均为约200 m2(18.5m×11.0 m),水深2.0 m. 试验所用蚌均为l龄的插片蚌,用网袋吊养在离水面40~50 cm处,网袋间隔为1.5m左右,每个网袋吊养4~5只蚌. 所用鲢、鳙也均为1龄鱼种,规格分别为鲢200 g/尾,鳙300 g/尾. 试验期间,围隔的放养情况如表3所示. 所有实验围隔每周施肥一次,每次施肥6.95 g/m2. 所用肥料含氮、磷分别为TN 8.0%,TP 5.0%.
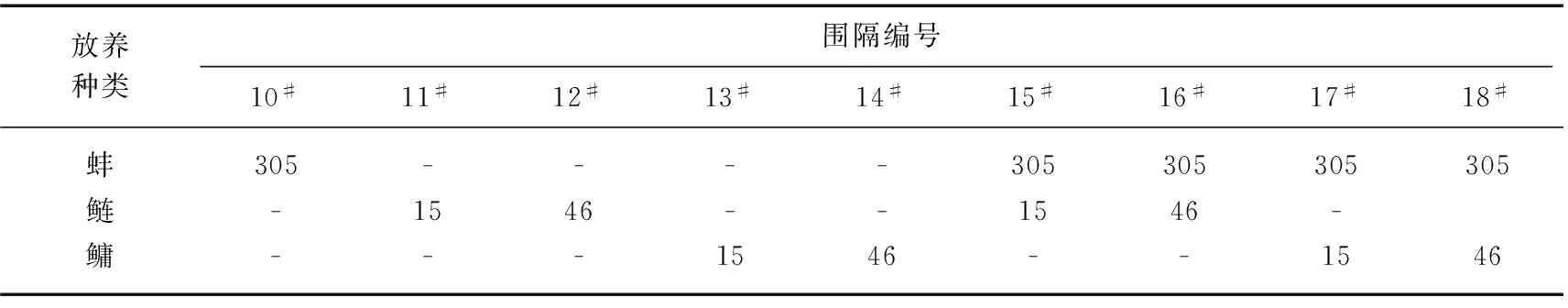
表3 金华池塘控藻围隔实验放养蚌、鱼数量
结果表明,在三角帆蚌、鲢和鳙3种滤食生物均单独养殖的围隔中,浮游植物生物量在蚌围隔最高,在高密度鲢围隔最低,且两者之间差异显著. 总体上,有鲢、鳙的围隔浮游植物生物量均低于有蚌的围隔,表明与蚌相比,鲢、鳙更能降低水体中的浮游植物生物量. 从浮游植物优势种组成来看,养蚌围隔蓝藻生物量显著高于鲢、鳙围隔,而绿藻生物量又明显低于鲢、鳙围隔;且围隔中蓝藻生物量又随鲢密度的增加而进一步降低. 研究显示,在蚌池混养鲢或鳙后,浮游植物生物量也均显著下降,且混养鲢的围隔下降更显著,表明在池塘围隔条件下养殖鲢、鳙也均能使藻类特别是蓝藻生物量下降,特别是鲢对藻类的控制作用更加明显.
该研究再次证明,鲢和鳙的滤食能够降低水体中的蓝藻数量,而蚌的滤食不但不能控制藻类,反而会促进蓝藻的增殖,从而可能诱发蓝藻水华. 这个结果,希望能被那些用蚌开展水环境修复的研究者们所重视.
案例三:天津于桥水库的控藻试验
南开大学于2007年在天津于桥水库北岸的一废弃鱼池经改造后进行了鲢、鳙控藻试验[20]. 该鱼池南北长100 m,东西长30 m,深3 m. 泥底,边坡45°. 试验将该鱼池用防水布分隔成30 m×10 m的10个围隔,于2007年6月4日注水至1.6m. 按照鱼的投放密度设置了高鲢鱼(HS,60 g/m3)、中鲢鱼(MS,30 g/m3)、低鲢鱼(LS,10 g/m3)、低鳙鱼(LB,10 g/m3)和对照(NF,无鱼)共5个处理组,每个处理组设2个重复,共10个围隔. 2007年6月11日按预设密度向各处理组围隔中投放鲢、鳙鱼苗后试验开始(记为第0 d),至2007年7月19日第1阶段试验结束. 试验期间未向围隔中注水,无投放肥料和饵料. 试验用鱼苗购自于桥水库附近渔场,均为同批次的1龄鱼,其中鲢平均尾重252±6.2g,平均体长23.4±0.4cm;鳙鱼平均尾重239.9±3.0 g,平均体长23.8±0.6cm.
试验期间水温在25.0~29.5℃之间波动,统计检验显示各处理间水温均值并无显著差异;放鱼后各处理组pH值以HS、MS、LS、LB、NF组依次递增,HS、LS、NF组相互之间pH值差异显著,而TP、TN浓度和透明度则处理之间均无显著性差异,TP浓度较放鱼前(均值0.025 mg/L)明显上升,TN浓度和透明度则较放鱼前(0.84 mg/L和1.08 m)明显下降;放鱼后,有鱼处理组叶绿素a浓度均较放鱼前(均值4.90 μg/L)有不同程度的上升,而NF处理组则略有下降;LS处理组叶绿素a平均浓度最高,NF组浓度最低,随鲢鱼密度升高,叶绿素a浓度有下降趋势. 经多重比较,NF处理组与MS、LS处理间叶绿素a浓度具显著差异,而HS、LB处理组与其他处理间则无显著差异. 各处理组浮游植物群落结构在放鱼后均发生了明显变化(表4).
放鱼前,绿藻门为各处理组中的绝对优势门类,优势种主要是空星藻(Coelastrumsp.)和盘星藻(Pediastrumsp.);放鱼后,绿藻所占比例有所下降,硅藻比例明显上升,硅藻和绿藻共同成为优势门类. 裸藻和隐藻比例则较放鱼前有所升高,而蓝藻比例仅在对照组中有所上升,甲藻比例在有鱼处理组中明显下降;放鱼后,各有鱼处理组的优势种变为硅藻门的最小曲壳藻(Achnanthesminutissima)和绿藻门的角星鼓藻(Stauras-trumsp.);NF处理组中绿藻门优势种仍为放鱼前的空星藻和盘星藻,但比例有所下降,而蓝藻门的微囊藻(Microcystissp.)和拟鱼腥藻(Anabaenopsissp.)比例不断上升,至试验结束时,NF处理组水面有明显的一层微囊藻颗粒,形成轻度蓝藻水华.

表4 天津于桥水库控藻试验主要浮游植物质量百分比组成(%)
该试验结果显示,投放鲢、鳙后藻类总生物量以及叶绿素a水平并未下降,反而有不同程度的上升,但使大型藻类特别是使微囊藻等群体蓝藻数量下降. 因此他们得出的结论是,鲢、鳙不能控制藻类总生物量,但可以控制蓝藻的数量.
案例四:笔者实验室在2013年6-9月在上海浦东新区老港镇开展了一项罗氏沼虾养殖围隔中利用鲢、鳙控藻的研究. 试验设置了虾单养、虾蚌二元混养、虾鲢鳙三元混养和虾蚌鲢鳙(蚌设不同密度)四元混养等实验组(各设4个重复),试验的详细方法请参阅文献[21]. 研究结果显示,无需镜检,只要到现场就能轻易看出,虾单养、虾蚌混养围隔内均有明显的蓝藻水华,而虾鲢鳙混养和虾蚌鲢鳙混养的围隔均未见蓝藻水华. 试验结果清楚地表明,鲢、鳙可以控制水体中的蓝藻数量.
2.2.2湖泊(水库)中的控藻研究案例除了上述在池塘中开展的控藻研究外,国内也开展了很多在湖泊中的控藻试验研究.
案例一:滆湖网围中的鲢、鳙控藻试验
2011-2013年笔者实验室与江苏省滆湖渔业生产管理委员会办公室合作在江苏滆湖开展了一项新式网围控藻养殖试验[22]. 与传统网围相比,该新式网围结构上更简洁美观,网衣主要靠浮子浮于湖面上,而不是像传统网围那样高出湖面,网围周边用不锈钢框架固定,而不是传统毛竹固定. 最关键的是,网围内放养的鱼种,不是吃食性鱼类,而主要是鲢、鳙,*1亩=0.0666667 hm2.适当搭配少量底层的杂食性或食有机碎屑鱼类(如细鳞鲴或鲫鱼等). 网围内鱼类放养情况见表5. 整个网围养殖不投喂任何人工饲料,鲢、鳙主要滤食湖中的浮游生物,搭配的鱼类主要摄食网衣上的着生藻类和水体中的有机碎屑. 整个网围面积达137 hm2,研究结果表明,尽管网围水体与湖水相通,但网围内监测到的蓝藻数量显著少于网围外湖水,鲢、鳙控藻效果十分明显.
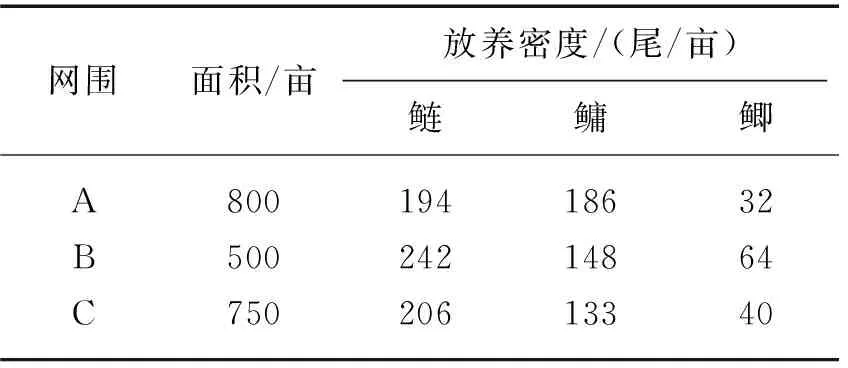
表5 新式网围控藻试验鱼种放养情况*
案例二:太湖贡湖湾的控藻研究
贡湖是太湖东北部靠近无锡市的一个湖湾,是无锡市的主要饮用水源地,其面积为150 km,水深为1.8~2.5m. 2007年贡湖湾暴发了严重的蓝藻水华,引发了无锡市的饮水危机,因此Guo等在该水域开展了旨在控制蓝藻水华的控藻试验[23]. 研究者们于2008年底在贡湖设置了一个巨型围隔(320 m×250 m),围隔内外没有水交换. 用于控藻的鲢、鳙于2009年3月投放到围隔内,规格分别为120~170 g/尾和80~120 g/尾,鲢和鳙放养密度分别为7.5和1.1g/m3,共有1200 kg鲢和180 kg鳙鱼种被投放到该围隔内,然后监测了围隔内外的水质和浮游动植物组成及其变动格局,结果显示,围隔内外浮游植物的季节变动格局基本类似:即围隔内外均呈现出1-6月以蓝藻(主要是微囊藻)和硅藻(主要是小环藻)为主、7-11月以蓝藻(微囊藻)为主、12月蓝藻迅速下降,隐藻和硅藻成为优势类群的总体格局. 1-6月围隔内藻类生物量大于围隔外的湖水,但在暴发藻华的7-9月,围隔内的藻类生物量小于围隔周边湖水. 围隔外湖水中蓝藻占了90%以上,但围隔内蓝藻仅占40%~80%. 在1-6月间围隔内微囊藻生物量较低,7月微囊藻生物量迅速上升达到峰值,其在湖水和围隔内分别达到60.6和3.6mg/L. 与湖水相比,水华期间围隔内的微囊藻生物量下降了94.1%. 围隔内、外微囊藻的年平均生物量分别为0.61和7.62 mg/L. 表明,利用鲢、鳙控制富营养化水体中的微囊藻是可行的.
很多鲢、鳙控藻的研究也发现,虽然蓝藻数量能得到控制,但叶绿素a浓度可能不降反升. 从贡湖的研究来看,虽然1-6月围隔内叶绿素a浓度显著高于湖水,但试验进行至7月藻类达到高峰时,湖水中的叶绿素a浓度又显著高于围隔,表明利用鲢、鳙控制藻类总生物量的目的可以达到,特别是利用鲢、鳙削减藻类增长高峰时的藻类生物量是可以达到预期的,此后随着水温的降低,内外差异缩小,两者差异也变得不显著.
3 对鲢、鳙控藻试验成败的探析
从对上面各个研究案例的介绍中不难看出:无论鲢、鳙成功控藻或不能控藻的试验结果,都可重复实现,即试验出现其中任何一个结果皆非偶然. 那么为什么鲢、鳙控藻会有成与败两种结果,是什么因素导致鲢、鳙控藻的成与败呢?
虽然要明确回答这些问题,可能仍需要开展一些验证性的研究来加以证实,但我们通过对上述研究案例的仔细分析仍可看出其中的一些端倪:即这两类研究在试验方法上存在着显著的差异. 在鲢、鳙不能控藻的试验中,研究者几乎不约而同地都使用了有底的围隔(个别使用了水族缸,相当于有底围隔),甚至有些还人为地添加了仅有数厘米厚的淤泥. 在有底围隔实验条件下,鱼类牧食藻类形成的粪便及排泄出来的氮、磷是很难被其他因素转化的,相反鲢、鳙牧食排泄形成了所谓的营养短路,加快了氮、磷的周转,为藻类重复利用,即使在人工添加了底泥的情况下,由于这些底泥不能固定在围隔底部,在试验过程中会被搅动、甚至被搅起,从而不但不能起到对有机质和氮、磷的吸附作用,反而会在不断地搅动中释放出更多的氮、磷. 因此不管有无添加底泥,有底围隔的研究相当于氮、磷输入没有得到较好的控制,故无一例外地在试验期间使叶绿素a浓度增加了;相反,在所有鲢、鳙成功控藻的试验中,所用围隔都是无底的(即自然底质),有的更是直接在自然水体(如池塘或湖泊)中进行,试验过程中这种底泥通常不会被扰动,能较好地起到对有机质(如鱼类粪便)和氮、磷的吸附,有利于降低水体营养盐的可获性,从而能起到降低总叶绿素和控制藻类的目的. 有的研究在深水湖泊(水库)中进行,有机质沉降后也不会被再悬浮,因此在这样的实验系统中,鲢、鳙对藻类摄食后能有效地降低其数量,达到鲢、鳙控藻目的.
也许这样的解释还不足以让你信服,那不妨再来看看下面的这个案例:利用隔离水界研究池塘施磷肥的效果和鲢、鳙鱼控制水域富营养化的作用[24].
该试验在上海市青浦县淀山湖联营场的一口面积为0.5hm2、平均水深2.5m的池塘中开展,该池塘以放养鲢、鳙和草鱼为主,放养量为3255尾/hm2. 研究者用纯橡胶做成3个隔离水界(即围隔),在水中浸泡7 d 后于1990年6月20日安装在池塘东北角离塘埂各5 m处. 隔离水界的长宽均为3 m,上部有0.5m左右露出水面,水深为2.2m;其中有两个是无底隔离水界,水界中的水能与底泥接触,有一个是有底水界,水与底泥隔离,不能接触. 6月28日在一个无底隔离水界和一个有底隔离水界中各放养鲢、鳙鱼种,8月30日起捕,放养和起捕情况见表6.
研究者在3个隔离水界和池塘中分别采集水样,采样时间都在上午7:00(当时采用夏令时,采样时间为夏令时8:00),采样点在水面下0.3m和1.5m处. 把两个水层的水样混合后再取样测试. 每隔1~2 d测定有效磷和叶绿素a浓度. 在7月17日-8月27日每隔1~2 d进行浮游动植物的定性和定量分析,每隔半个月测定COD(碱性高锰酸钾法),初级生产力用黑白瓶法,白天每隔4 h、晚上每隔6 h测定溶解氧,在7月18日和8月6日中午12:00时(晴天)分别向3个隔离水界施50.0 g Na2HPO4·12H2O+17.5g NaH2PO4·2H2O作为磷肥. 在8月17和25日中午12:00时(晴天)分别用谷耙翻动无底隔离水界的底泥. 试验期间隔离水界和池塘的水温为26.4~34.0℃,COD为10.02~18.26 mg/L,pH值为7.8~9.1,透明度为0.25~0.60 m.

表6 隔离水界鱼种放养和起捕情况
研究结果显示,围隔施磷肥后,单位面积磷的日减少量为:无底有鱼围隔>无底无鱼围隔>有底有鱼围隔. 测定的初级生产力则为:无底有鱼围隔>有底有鱼围隔>无底无鱼围隔,因此藻类通过光合作用吸收的有效磷也一定是:无底有鱼围隔>有底有鱼围隔>无底无鱼围隔. 两相比较可知,有无底泥对磷的吸附,是导致不同围隔中磷减少的主要原因,底泥吸附对磷的减少作用远大于鱼类的作用.
该研究还有一个值得关注点是,翻动底泥对围隔磷的影响. 研究结果显示,施肥前,无底围隔和池塘中的有效磷含量与有底围隔的有效磷含量无显著差别,说明没有外界干扰时底泥对磷的释放作用很小. 而翻动底泥的影响则视水体磷本底而定,若水体磷浓度已较高,翻动底泥对磷释放作用不明显,而若水体磷浓度较低,翻动底泥对磷释放作用非常大.
该研究结果还显示,放鱼围隔的有效磷和叶绿素a浓度都比不放鱼围隔低,表明放养鲢、鳙能降低TP浓度和藻类数量.
这个实验对我们的启示是,水体中藻类(或叶绿素a)的增长,直接取决于水体中磷的可获性. 在有底围隔中,由于缺少了底泥对磷的吸附,甚至相反由于鱼类对底部的扰动加速了磷的释放,导致磷不降反升,从而也促进了叶绿素a浓度的增加;而无底围隔,则由于磷被底泥吸附,有效地降低了磷的可获性,使得藻类数量下降. 因此,鲢、鳙对藻类的控制,除了直接的牧食作用外,更主要的是对水体中磷可获性的改变. 而对磷可获性最直接的影响,就是围隔的结构:有底和无底.
综合上述分析,我们认为,围隔实验条件的差异,是目前造成鲢、鳙控藻成败不同结果的主要原因. 有底围隔实验条件下得出的都是鲢、鳙不能控藻的结果,而在无底围隔条件下大多能得到鲢、鳙可以控藻的研究结果.
那么是否在有底围隔条件下鲢、鳙就一定不能控制藻类呢?因为目前我们还缺少相关的实验研究证据,所以也不能对此轻易作出肯定或否定的回答. 尽管如此,我们仍可通过既有的一些文献对此作出推断. 首先,之所以多数有底围隔鲢、鳙不能控藻,其原因可能是:由于在围隔有底的情况下,鲢、鳙摄食藻类后排泄的氮、磷不能被有效削减,增加了氮、磷的周转率,相当于湖泊污染源没有得到很好的控制,因此原先设定的鲢、鳙密度便不能达到控藻的目的,此时若能进一步加大试验所用鲢的密度,藻类数量也仍有可能被控制住. 如在唐汇娟、谢平的研究案例[14]中,刘恩生[27]就进行了进一步的推算,结果发现,该实验系统中,藻类密度(y)与鲢(x)之间存在着这样的关系:
y=-0.0002x2+0.0869x+1.5007(R2=0.9696)
由该关系可知,只要鲢的密度增加到一定阈值,围隔中的藻类数量仍是可以被控制住的. 在实践中是否如此,仍有待研究去证实.
4 影响鲢、鳙控藻的其他因素分析
除了上述围隔结构类型会影响鲢、鳙控藻的效果外,在实际应用中还有一些其他因素也会影响到鲢、鳙的控藻效果.
首先,鲢、鳙的密度大小会影响其效果. 由于湖泊中藻类数量的增长主要取决于其增长与死亡两种影响因素的对比. 通常富营养化条件下氮、磷等营养物丰富,有利于藻类繁殖,促进其数量增长,这种效应通常被称为上行效应;另一方面,鲢、鳙和浮游动物对藻类的牧食,是导致其数量下降的最主要因素,该因素则被称为下行效应(或下行控制). 如果水体中的鲢、鳙数量低,其下行效应小于上行效应,就会导致藻类数量的增长,这也是千岛湖在中营养条件下也会暴发蓝藻水华的重要原因[25]. 鲢、鳙密度(或生物量)对藻类的这种影响虽然不难被理解,然而在实践中却常常容易被误解. 即有时候虽然湖泊中放养了鲢、鳙,但由于其确凿的数量常缺少简便有效的方法加以评估,使得湖中鲢、鳙数量是否足够不能轻易地被判断出来. 此时,如果鲢、鳙数量仍不足于控制上行效应时,藻类数量仍将表现出一定的增长,即出现了鲢、鳙放养不能控制藻类增长的表象,从而可能使鲢、鳙的实际控藻作用被否定.
其次,有无其他藻食生物参与控藻也会影响到鲢、鳙的控藻效果. 即在鲢、鳙控藻时有无浮游动物参与,其结果也会有所不同. 在自然水体中,浮游动物也会与鲢、鳙起协同控藻的作用. 尽管鲢、鳙的放养会对浮游动物种群起到一定的压制,使其数量有一定的下降,然而,与实验围隔不同的是,在自然水体中,鲢、鳙从来都不会把浮游动物彻底消灭掉,如据我们在千岛湖的常年监测,千岛湖实施保水渔业(鲢、鳙控藻)后,水体中的大型浮游动物(如透明溞)仍保持着一定的数量. 而且根据我们的野外采样观察,浮游动物在水下10~20 m均有大量分布,即其与鲢、鳙形成了明显的生态位分化. 浮游动物之所以在深水层分布,除了躲避鲢、鳙捕食的需要外,也可能与该水层的食物资源丰富有关. 大量研究表明,鲢、鳙放养会导致藻类小型化,而藻类小型化恰恰有利于浮游动物的摄食. 通常小型浮游植物,特别是硅藻类,其分布水层较深. 同时鲢、鳙粪便等形成的有机碎屑,也会在沉降至中下水层时而成为其食物资源. 因此在自然湖库生态系统中,一方面,鲢、鳙摄食大型浮游植物后,使水体中的营养物更有利于为大量小型或超微浮游植物利用,从而为浮游动物提供更多的食物,同时鲢、鳙摄食形成大量的有机碎屑在沉降过程中也会为分布于深水层的浮游动物提供食物;这些因素都会促进浮游动物数量的增长. 另一方面,尽管鲢、鳙会牧食掉水体中的部分浮游动物,但在这些大中型的自然湖库生态系统中,鲢、鳙与浮游动物均有各自的生态位,当鲢、鳙在某个局部水域觅食时,其他水域的浮游动物就得到了繁衍生息,因此鲢、鳙永远也不可能彻底食尽水中的浮游动物,从而使鲢、鳙和浮游动物共存成为可能,发挥鲢、鳙与浮游动物之间的协同控藻作用. 即使在浅水湖泊如武汉东湖,虽然浮游动物不可能像千岛湖那样分布在深水层以躲避捕食,但也因为鲢、鳙并非多到随处都是,因此也会在水平方向形成生态位,即有大量没有鲢、鳙的水域可供其栖息繁衍,因此在这些自然湖库中,鲢、鳙和浮游动物总能共存和起到协同控藻的作用. 因此在自然湖库条件下放养鲢、鳙,其对藻类的控制作用是两者共同作用的结果. 鲢、鳙与浮游动物协同控藻会使控藻效果更佳,也能一定程度上减缓藻类小型化的速度.
第三,水体大小也会影响鲢、鳙控藻的效果. 水体大小不同除了会影响到浮游动物的数量,从而影响到鲢、鳙能否与其协同控藻外,水体大小对鲢、鳙控藻的影响还表现在鲢、鳙活动对水体的物理搅动作用的大小及其对生态影响上. 在自然湖库生态系统中,因人工放养的鲢、鳙等鱼类的游动对水体的物理扰动作用通常都远小于风浪、湖流等自然因素,因此鲢、鳙活动这种作用的影响可以是忽略的,然而,在微型或小型实验系统中,鱼类的扰动则足以大到搅动整个水体及人工添加的底泥,从而促进底泥中的营养物释放,进而使其与无鱼系统产生显著差异. 其次,在大型自然湖库生态系统中,鲢、鳙的排泄对整个水体的氮、磷输入贡献不大,而在小型封闭式围隔中,鲢、鳙排泄对氮、磷的输入起着显著的作用,从而对藻类增殖起到显著影响.
此外,实验时间的长短也会对实验结果产生很大的影响. 在鲢、鳙控藻实验系统中,藻类数量的变动取决于鲢、鳙等对藻类牧食引起的数量减少与藻类繁殖而使数量增长之间的净效应. 由于藻类繁殖速度快,而鲢、鳙在实验系统中还需要有一定的适应时间等因素的存在,即使在很高的鲢、鳙放养密度系统中,都可能会出现短期内藻类数量的增长. 因此为了观察到鲢、鳙对藻类的控制作用,往往需要有足够长的相互作用时间. 通常实验时间越长,其结果的可靠性也越大.
还有一个因素也许也值得关注:水体中的微生物组成. 在自然系统中,通常会有各种微生物,特别是各种硝化细菌等,可以起到对氮的转化作用,从而影响水体中的藻类;但在实验系统中,很少有研究考虑微生物的参与,通常只有循环水养殖系统会考虑给以一定的微生物“熟化”培养时间,而在实验控藻系统中很少有研究会考虑微生物的培养(硝化细菌等微生物为自养生物,其培养时间要在4~7周,因此那些仅有1~2个月的实验,微生物的作用被完全忽略了),因此实验系统的食物网结构远比湖库鲢、鳙控藻系统简单. 通常系统越小,其实验结果与真实的偏差就会越大. 而我们最近的研究更是显示了有无鲢、鳙的存在,其微生物组成差异显著[26]. 当然微生物组成的差异与控藻的关系仍需要更多的研究加以证实.
在文献中,我们有时也能看到这样的结论:认为鲢、鳙放养后形成了所谓的营养短路,从而加速了水体的富营养化和藻类的增长. 实际上,得出这样的结论是很可笑的,那等于承认了“永动机的存在”:即水体放养鲢、鳙后,可以不断形成所谓的营养短路,变成藻类,不断地为鲢、鳙提供食物. 显然这与事实不符. 我国的养殖户早就在实践中发现了这样的事实:要使鲢、鳙鱼类不断生长,就得不断地向水体中追加肥料来肥水. 从生态学原理上讲,这就是简单的上行效应.
同样的道理,在湖库中,鲢、鳙的产量主要取决于其食物资源的丰度,即浮游生物越丰富,鲢、鳙的产量越高,反之亦然,这是水体中上行效应的必然结果,我国百姓利用上行效应开展施肥养鱼已有近千年的历史. 如果仅仅根据鲢、鳙产量达到某一产量时的叶绿素a浓度高低来判断鲢、鳙对藻类是否存在控制作用显然是有悖常理的,即根据调查鲢、鳙产量大于100 kg/hm2的湖泊,其叶绿素a浓度高来判断鲢、鳙不能控藻,显然是不合理的,这恰恰是颠倒了因果关系的缘故. 要从统计角度去判断鲢、鳙能否控藻,需要在相同的营养条件下考察不同鲢、鳙生物量与叶绿素a浓度的关系,方能说明鲢、鳙与藻类之间的关系.
还有根据鲢、鳙肠道粪便中分离的藻类是否具有光合活性来判断其是否消化吸收蓝藻也是不尽科学的. 这是因为,首先,鲢、鳙在高密度藻类存在时为了保持呼吸顺畅,会产生假粪,从假粪分离的藻类,当然会具有光合活性;其次,即使没有假粪,从其粪便中分离的藻类具有光合活性,也不代表其没有消化吸收蓝藻. 只是代表鲢、鳙没有100%地消化藻类而已. 重要的不是其是否能够消化蓝藻,而是看其对蓝藻消化的数量是否能超过其繁殖的量,从而通过不断地牧食,最终达到控制蓝藻数量和遏制水华发生的目的.
5 对鲢、鳙控藻作用的共识
通过上述分析,我们似乎可以对鲢、鳙控藻形成这样的共识:鲢、鳙可以控制蓝藻,之所以在某些条件下鲢、鳙没有成功地降低蓝藻数量有两方面原因:一是因为实验条件的限制,即研究者采用了有底围隔,使实验系统结构太过简单. 如果将实验条件改为有自然的底质,且实验时间延续得足够长,实验结果应可以出现预期的结果:鲢、鳙能够降低水体中蓝藻或藻类总数量. 二是在某些条件下,由于水体中鲢、鳙密度太低,从而使鲢、鳙产生的下行控制,不能有效制衡富营养化产生的上行效应. 如果增加鲢、鳙数量,也是可以预期到鲢、鳙对藻类的遏制. 无论是基于稳定性同位素或PCR-DGGE食性分析,还是通过合理的围隔实验,以及大量的养殖实践,都一再证实了鲢、鳙控藻的基本事实.
6 对鲢、鳙控藻今后应开展的研究
我们的上述分析,可能还不足以使人们彻底消除对鲢、鳙控藻的意见分歧. 这也意味着,未来关于鲢、鳙控藻的研究仍有继续的必要;然而值得指出的是,未来的研究不应是对相关问题的简单重复,而应围绕一些关键问题开展更深入的研究,才会使鲢、鳙控藻的相关研究更具价值.
首先,有必要开展对本文提出的导致鲢、鳙控藻成败关键原因的验证性研究,即利用有底和无底围隔进行一些验证性的研究,看看其结果是否正如我们分析的那样可以预期,以便使导致鲢、鳙控藻成败的真正原因得到广泛的认可.
其次,要通过未来的研究,还要能找出鲢、鳙在不同营养程度时成功控藻的临界密度,以便更好地指导实践.
第三,要研究鲢、鳙控藻最佳效率时的水体营养条件. 即由于水体富营养化程度太高时,藻类密度会过高,鲢、鳙为了保护自身呼吸和其他生理需要,会以假粪形式摄食藻类,从而不能有效发挥控藻、遏藻作用,因此未来就需要研究在何种水体营养条件下采取鲢、鳙控藻,以致使水体中藻类不会达到如此高密度,从而发挥鲢、鳙控藻的最佳效果.
实际上,有关鲢、鳙控藻的研究还可以包括很多,如鲢、鳙的最佳比例,鲢、鳙对不同藻类的可消化率,鲢、鳙食性的可塑性及其影响因素,鲢、鳙在水体中的活动规律,鲢、鳙放养对其他鱼类乃至整个生态系统的影响等等. 所有这些研究,都将有助于增进我们对鲢、鳙控藻的深入了解和更好应用,同时能有效规避鲢、鳙放养可能对生态系统产生的不利影响. 但不管开展怎样的研究,未来的研究都不应是那些单一系统中鲢、鳙对藻类影响的重复研究.
7 结语
鲢、鳙控藻的研究有成功和不成功两种实验结果,两种结果都能被重复验证. 出现两种研究结果的原因,可能主要是两种研究的实验条件不同所致:在有底围隔或水族箱条件下,由于系统结构简单,鱼类排泄的氮、磷缺少迁移转化的途径,以致能被实验系统中的藻类反复利用,导致鲢、鳙控藻的失败;而在无底围隔或自然系统中,鲢、鳙的控藻作用不仅仅是鲢、鳙对藻类的直接控制作用,而是鲢、鳙介导的综合控藻作用,即除了鲢、鳙对藻类的直接牧食外,还包括鲢、鳙牧食后加快了对颗粒磷的沉降(鲢、鳙粪便大于浮游动物的粪便,即如果没有鲢、鳙对藻类的摄食,这些浮游植物就会被浮游动物摄食,因此相当于鲢、鳙粪便与浮游动物粪便的比较)和底质对这些颗粒磷和有机碎屑的吸附,氮、磷特别是磷的迁移转化影响了藻类以及浮游动物对藻类的协同控藻作用等,因此通常都能取得理想的控藻效果.
此外,影响鲢、鳙控藻效果的因素,还包括水体中鲢、鳙的实际密度,水体的大小,水体的营养程度,实验持续的时间以及实验系统的食物网结构等. 总之,利用有底围隔得出的结果,由于与实际湖泊生态系统的条件有太多的差异,因此并不能据此得出鲢、鳙在自然生态系统中不能控藻的结论,相反,已有大量的研究和实践证明,在点源污染得到有效控制的水体中,利用鲢、鳙控制富营养化水体的藻类过度增长是可行的.
[1]FAO. The state of world fisheries and aquaculture 2012. Rome, 2012.
[2]刘建康. 东湖生态学研究(一). 北京: 科学出版社, 1995.
[3]Liu Q, Chen Y, Li Jetal. The food web structure and ecosystem properties in a filter-feeding carps dominated deep reservoir ecosystem.EcologicalModeling, 2007, 203(3/4): 279-289.
[4]Smith DW. Biological control of excessive phytoplankton growth and the enhancement of aquacultural production.CanadianJournalofFisheriesandAquaticSciences, 1985, 42: 1940-1945.
[5]李琪, 李德尚, 熊邦喜等. 放养鲢鱼(HypophthalmichysmolitrixC et V)对水库围隔浮游生物群落的影响. 生态学报, 1993, 13(1): 30-37.
[6]Domaizon I, Dévaux J. Experimental study of the impacts of silver carp on plankton communities of eutrophic Villerest reservoir (France).AquaticEcology, 1999, 33: 193-204.
[7]谢平. 鲢、鳙与藻类水华控制. 北京: 科学出版社, 2003.
[8]Kajak Z, Rybak J, Spodniewska Ietal. Influence of the planktivorous fish,Hypophthalmichthysmolitrix(Val.) on the plankton and benthos of the eutrophic lake.PolskieArchiwumHydrobiologii, 1975, 22: 301-310.
[9]Burke JS, Bayne DR, Rea H. Impact of silver and bighead carp on plankton communities of channel catfish pond.Aquaculture, 1986, 55: 59-68.
[10]Spataru P, Gophen M. Feeding behaviour of silver carpHypophthalmichthysmolitrixVal. and its impact on the food web in Lake Kinneret, Israel.Hydrobiologia, 1985, 120(1): 53-61.
[11]史为良, 金文洪, 王东强等. 放养鲢鳙对水体富营养化的影响. 大连水产学院学报, 1989, 4(3): 11-20.
[12]刘建康, 谢平. 用鲢鳙直接控制微囊藻水华的围隔试验和湖泊实践. 生态科学, 2003, 22(3): 193-198.
[13]阮景荣, 戎克文, 王少梅.微型生态系统中鲢、鳙下行影响的实验研究——1.浮游生物群落和初级生产力. 湖泊科学, 1995, 7(3):226-234. DOI 10.18307/1995.0304.
[14]唐汇娟, 谢平. 围隔中不同密度鲢对浮游植物的影响. 华中农业大学学报, 2006, 25(3): 277-280.
[15]孙育平. 营养盐加富、滤食性鱼类和浮游动物对水库浮游植物群落结构的影响[学位论文]. 广东: 暨南大学, 2010.
[16]Wang H, Liang X, Jiang Petal. TN∶TP ratio and planktivorous fish do not affect nutrient-chlorophyll relationships in shallow lakes.FreshwaterBiology, 2008, 53: 935-944.
[17]王银平, 谷孝鸿, 曾庆飞等. 控(微囊)藻鲢、鳙排泄物光能与生长活性. 生态学报, 2014, 34(7): 1707-1715.
[18]Mueller CR, Eversole AG. Effect of silver carpHypophthalmichthysmolitrixand freshwater musselElliptiocomplanatafiltration on the phytoplankton community of partitioned aquaculture system units.JournaloftheWorldAquacultureSociety, 2004, 35(3): 372-382.
[19]周小玉, 张根芳, 刘其根等. 鲢、鳙对三角帆蚌池塘藻类影响的围隔实验. 水产学报, 2011, 35(5): 35-39.
[20]王嵩, 王启山, 张丽斌等. 水库大型围隔放养鲢鱼、鳙鱼控藻的研究. 中国环境科学, 2009, 29(11): 1190-1195.
[21]杨洋. 鲢、鳙和三角帆蚌与罗氏沼虾不同混养系统的结构优化与氮磷收支研究[学位论文]. 上海: 上海海洋大学, 2014.
[22]贾佩峤. 滆湖ecopath模型构建及围网放养鲢鳙的生态效应[学位论文]. 上海: 上海海洋大学, 2012.
[23]Guo L, Wang Q, Xie Petal. A non-classical biomanipulation experiment in Gonghu Bay of Lake Taihu: Control ofMicrocystisblooms using silver and bighead carps.AquacultureResearch, 2015, 46: 2211-2224.
[24]谭玉钧, 李家乐, 康春晓. 利用隔离水界研究池塘施磷肥的效果和鲢、鳙鱼控制水域富营养化的作用. 见: 朱学宝, 施正峰编. 中国鱼池生态学研究. 上海: 上海科学技术出版社, 1995.
[25]刘其根, 陈立侨, 陈勇. 千岛湖水华发生与主要环境因子相关性分析. 海洋湖沼通报, 2007,(1): 117-124.
[26]唐永涛, 赵良杰, 刘其根等. 基于16S rDNA比较研究混养三角帆蚌和鲢鳙对池塘养殖水体微生物群落结构的影响. 淡水渔业, 2015, (3): 70-77.
[27]刘恩生. 生物操纵与非经典生物操纵的应用分析及对策探讨. 湖泊科学, 2010, 22(3): 307-314. DOI 10.18307/2010.0301.
Controlling the nuisance algae by silver and bighead carps in eutrophic lakes: disputes and consensus
LIU Qigen & ZHANG Zhen
(CollegeofFisheriesandLifeScience,ShanghaiOceanUniversity,Shanghai201306,P.R.China)
Treatment of lake eutrophication is shifting from pollution control towards restoration and management of lake ecosystems in China, so it has drawn much attention of the non-traditional biomanipulation based on the role of silver carp and bighead carp in lakes. However, there are both successful and unsuccessful cases over the use of silver carp and bighead carp in controlling nuisance algae, which has led to the discrepancy over the feasibility of its usage. In this paper, we analyzed many studies that tested the effects of silver carp and bighead carp on algal production, pinpointed out the possible causes leading to the different results in these case studies, and tried to reach a consensus over the use of silver carp and bighead carp to control algal over growing in eutrophic lakes. We hope this effort could play a role in helping the environmental management and protection of lakes in China.
Lakes; eutrophication; silver carp; bighead carp; cyanobacterial bloom; control of nuisance algae
J.LakeSci.(湖泊科学), 2016, 28(3): 463-475
10.18307/2016.0301
©2016 byJournalofLakeSciences
*公益性行业(农业)专项(201303056)、国家水体污染控制与治理科技重大专项(2012ZX07101-007)、国家自然科学
31072218)和上海市高校知识服务平台项目(ZF1206)联合资助. 2016-01-31收稿;2016-02-02收修改稿.
刘其根(1965~),男,博士,教授;E-mail:qgliu@shou.edu.cn.
