林窗大小对马尾松人工林凋落叶可溶性有机碳损失的影响1)
2016-08-17李勋刘洋张健杨万勤张艳邓长春张明锦刘华
李勋 刘洋 张健 杨万勤 张艳 邓长春 张明锦 刘华
(四川农业大学,成都,611130)
林窗大小对马尾松人工林凋落叶可溶性有机碳损失的影响1)
李勋刘洋张健杨万勤张艳邓长春张明锦刘华
(四川农业大学,成都,611130)
基于人工林近自然改造,调控低山丘陵区马尾松低效林林分结构,探明马尾松人工林不同面积林窗下,凋落叶可溶性有机碳(DOC)释放规律。以长江上游低山丘陵区人为采伐形成的马尾松人工林7种林窗(100、225、400、625、900、1 225、1 600 m2)为研究对象,研究不同面积林窗的马尾松(Pinusmassoniana)、红椿(Toonaciliata)、樟(Cinnamomumcamphora)、桢楠(Phoebezhennan)等凋落叶可溶性有机碳(DOC)的损失率。结果表明:(1)林窗面积对凋落叶DOC损失率有显著影响。中小面积林窗(225~625 m2)内凋落物的DOC损失率高于其它面积林窗。(2)经过1 a的分解,4种凋落叶的DOC损失率为61.55%~90.06%,其中:DOC损失率红椿最高,马尾松最低。(3)凋落叶的DOC损失主要集中在分解前期(0~90 d),此后凋落叶的DOC损失速率不断降低,且不同凋落物之间DOC损失速率的差异不断减小。因此,在马尾松低效林改造过程中,可利用中小面积林窗这一显著作用,引入乡土阔叶树种红椿、樟、桢楠,加速马尾松人工林的物质循环,维持林地肥力。
可溶性有机碳;林窗;马尾松人工林;马尾松凋落叶
We adjusted the structure of forest and explored the regularity of litter dissolved organic carbon (DOC) release in seven gap sizes of aPinusmassonianaplantation in a hilly area of the upper reaches of Yangtze River. The experiment was set up inPinusmassonianaplantation with seven different gap sizes (100, 225, 400, 625, 900, 1 225, and 1 600 m2) established by artificial thinning. The contents of DOC forPinusmassoniana(Pm),Toonaciliate(Tc),Cinnamomumcamphora(Cc) andPhoebezhennan(Pz) were evaluated in different sized forest gaps, with closed canopy as the control. Litterbags were collected from the gap center and the closed canopy in December of 2013, February, May, August and November of 2014. The forest gap size had significant effects on DOC loss rate small and medium sized gaps (225-900 m2) had greater effect on DOC loss rate forPm,Tc,Cc, andPzleaf litter. Over one-year decomposition, variation trend of DOC loss rate of descending order for four species wasTc(90.06%),Cc(85.17%),Pz(78.74%) andPm(61.55%). The DOC loss rate for four species mainly was concentrated in the early decomposition stages (first 90 days), and then was decreased quickly, and the difference of DOC loss rate (per 30 days) among four species diminished gradually. Therefore, when we reconstruct the low-function forest, we can take advantage of small and medium sized gaps, introduce native broad-leaf tree speciesTc,Cc,Pzto accelerate nutrient cycling, and maintain the soil fertility ofPinusmassonianaplantation.
凋落物分解过程中可溶性有机碳(DOC)的释放不仅是森林生态系统养分循环的重要环节,也对碳循环起着极其重要的作用[1-2]。DOC是森林生态系统中一类重要的、十分活跃的化学组分,也是微生物生长和生物分解过程中的重要能量来源。DOC对森林生态系统营养物质的有效性和流动性、污染物质的毒性及其迁移特性、pH有直接影响[3-4]。DOC也会影响矿物风化,与生态系统物质循环和能量流动具有密切的关系[5-7]。有关研究表明[8-10]:凋落物、土壤腐殖质、土壤微生物和根系分泌物等是森林生态系统DOC的主要来源;森林枯枝落叶层不仅通过分解作用向大气释放CO2,通过淋溶作用把DOC迁移到矿质土壤中,构成森林碳平衡的重要组成部分。
马尾松(Pinusmassoniana)是我国南方地区重要的造林树种之一,适应性强、耐干旱与瘠薄[11]。据全国第六次森林资源清查结果,马尾松人工林现有面积583.27万hm2,居人工林第2位[12]。近年来,由于马尾松人工林组成树种单一、层次结构简单、林分密度和郁闭度过大,导致其发展到中龄林阶段后,出现了林木长势普遍衰弱、地表植被盖度小、物种多样性低、生物多样性贫乏、土壤供肥能力不足、病虫害发生风险高、林分生产力低下等问题[13]。林窗作为森林群落中经常发生的小规模干扰,它的形成能改变林下微环境以及土壤养分的有效性等,为引入乡土阔叶树种进行低效林分改造提供了必要的环境条件。林窗大小是森林的重要空间特征,直接影响着林内的环境因子及其组合状况[14-15]。目前,国内外有关林窗和DOC的研究主要涉及林窗的形成、林窗对森林群落的组成和动态的影响[16-18],天然林或次生林林窗特征[19-20]及环境因子特征[21-23],土壤DOC的变化[24-27]等。林窗面积大小与凋落物的DOC损失率的关系,尚无明确的研究结论,因此,以四川宜宾来复镇马尾松人工林7个不同面积林窗为研究对象,通过引种乡土阔叶树种红椿、樟、桢楠,形成斑块状混交人工林,研究不同大小林窗下针叶(马尾松)和阔叶(红椿、樟、桢楠)凋落叶1 a分解过程中的DOC释放,为低山丘陵区马尾松人工林物质循环过程的合理调控、优化林分结构提供理论依据。
1 研究区概况
研究区位于四川盆地南缘,高县北部(104°21′~104°48′E,28°11′~28°47N)。气候类型属中亚热带湿润季风气候,年均气温18.1 ℃,月均气温大于10 ℃有278 d,年均降水量1 070.4 mm。气候温和,雨量充沛,雨热同季。植被绝大部分为次生植被与人工植被,境内亚热带次生常绿针叶林植被分布较广,该研究样地选自于高县来复镇毛顛坳的41年生人工马尾松林,海拔高度在400~550 m。马尾松人工林林分结构简单,乔木以马尾松为主林,主要灌木有:枹栎(Quercusserrata)、黄荆(Vitexnegundo)、黄牛奶(Symplocoslaurina)、梨叶悬钩子(Rubuspirifolius)、野梧桐(Mallotusjaponicus)、展毛野牡丹(Melastomanormale)等;主要草本有淡竹叶(Lophatherumgracile)、蕨(Pteridiumaquilinum)、马唐(Digitariasanguinalis)、芒(Miscanthussinensis)、芒萁(Dicranopterispedata)、皱叶狗尾草(Setariaplicata)等。
2 研究方法
2.1样地设置
2011年8月,对试验区域内的植物多样性、土壤理化性质、林分结构等进行初步调查。根据样地调查结果,于2011年10月在试验区选择林地质量相近、林分密度相同的42年生马尾松人工纯林进行采伐,形成7种(G1、G2、G3、G4、G5、G6、G7)大小不等,面积分别为:100、225、400、625、900、1 225、1 600 m2,形状近似正方形的林窗各3个,林窗边界间距大于10 m。未砍伐的纯林作为林下对照(CK),林下、林窗都设置3个重复。样地基本情况见表1。

表1 样地基本概况
注:表中pH、全碳质量分数、全氮质量分数、土壤密度、土壤最大持水量数据为“平均值±标准差”。
2.2试验设计与样品处理
2013年10月,在研究地的马尾松林群落内,收集新鲜的马尾松、红椿(Toonaciliata)、樟(Cinnamomumcamphora)、桢楠(Phoebezhennan)凋落叶,将样品带回实验室风干,然后分别称取10克叶片,将叶片装入上层面0.5 mm、下层0.04 mm孔径的尼龙袋(20 cm×20 cm)进行试验。另外,分别取相同质量(10克)风干的4种凋落叶9袋在65 ℃下烘干至恒质量,由其推算出网袋内样品的初始干质量。2013年11月17日,分别在人为采伐形成的马尾松人工林7种不同大小林窗以及林下放置红椿、樟、马尾松、桢楠凋落叶分解袋各120袋,总袋数=4物种×3重复×(7林型+1对照)×5次取样=480袋。且在对照和每个林窗下5 cm深土壤中分别埋设1个纽扣温度记录器,每个面积林窗设置3个重复,每2 h自动记录一次温度。分别于2013年12月17日、2014年2月17日、2014年5月17日、2014年8月17日、2014年11月17日收集凋落物袋,每次从每个样点内取回红椿、樟、马尾松、桢楠分解袋各3袋。清除网袋表面泥土和其他腐殖质,标记后装入无菌塑料袋中,带回实验室。
2.3样品分析与测定
将样品中其他凋落物和石子剔除后,在65 ℃下烘干至恒质量,称取质量后,磨碎后过2 mm筛,装入密封塑料袋保存,用于测定DOC。DOC的测定采用[28-30]:精确称取磨碎后过筛样品0.05 g于20 mL离心管中,加入10 mL蒸馏水,往复振荡机上振荡0.5 h;然后在温度4 ℃的条件下,用4 000 r·min-1的离心机离心15 min;取上清液,在-0.08 MPa压力下,经孔径为0.45 μm滤膜(上海市新亚净化器件厂生产,使用前首先要用蒸馏水淋洗滤膜,以淋洗滤膜可能残留的有机组分)过滤,即获得植物凋落物DOC提取液,在4 ℃温度下保存待测液。TOC分析仪(multi N/C 2100,AnalytikJena)测定待测液中DOC质量分数。土壤含水量采用铝合烘干法测定。全碳采用重铬酸钾加热法(GB 7657-87);全氮采用凯氏定氮法(LY/T 1269-1999);木质素和纤维素采用范氏(Van Soest)洗涤纤维法[31]。
2.4数据处理与统计分析
差量法计算凋落物质量损失率[32]:
L=((M0-Mt-1)/M0)×100%。
凋落物DOC损失率Rt和30 d DOC损失速率Vt[33]:
Vt=((Rt-1-Rt)/Δt)×30;
Rt=((M0C0-MtCt)/M0C0)×100%。
式中:L为凋落物的质量损失率;M0为凋落物初始质量;Mt-1为前一次凋落物残留量;Mt为当次凋落物残留量;C0为DOC初始质量分数;Ct为当次DOC质量分数;Rt为当次DOC损失率;Rt-1上一次DOC损失率;Δt为相邻采样间隔时间;t为采样时间。
采用SPSS 20.0软件进行统计分析,运用单因素方差分析方法和最小显著差异法(LSD)检验凋落物质量损失率和DOC损失率的差异。凋落物质量损失率和DOC损失率的关系用origin8.5进行拟合。数据整理、计算采用Microsoft Excel 2003软件进行,作图采用Origin8.5软件进行。
3 结果与分析
3.1土壤温度和含水率
由表2可知,各林窗土壤日均温变化趋势一致。2014年5—8月份最高,2013年12月—2014年2月最低,各林窗间土壤日均温度总体上随林窗增大而升高;2013年11—12月份、2014年2—5月份、5—8月份、8—11月份最高温分别出现在G7(14.06 ℃)、G7(11.22 ℃)、G6(19.07 ℃)、G7(25.68 ℃)、G7(20.97 ℃)。2013年11—12月份少数林窗土壤月均温低于林下。
由表3可知。土壤含水率受不同季节降雨量影响。即2013年11月—2014年2月最低,2014年3月开始升高。总体上看,土壤含水率随林窗面积增大,先升高后降低。

表2 马尾松人工林不同时期土壤日平均温度
注:表中数值为“平均值±标准差”,同列不同大写字母表示不同林窗之间差异显著(P<0.05),同行不同小写字母表示不同分解阶段差异显著(P<0.05)。

表3 不同分解阶段各林窗的土壤含水率
注:表中数值为“平均值±标准差”,同列不同大写字母表示不同林窗之间差异显著(P<0.05),同行不同小写字母表示不同分解阶段差异显著(P<0.05)。
3.2凋落叶初始基质状况
由表4可知,凋落叶可溶性有机碳质量分数为15.22~57.16 g·kg-1,树种间差异显著(P<0.05)。其中,红椿凋落叶DOC质量分数最高(57.16 g·kg-1),显著高于其他物种;而马尾松凋落叶DOC质量分数最低(15.22 g·kg-1)。各物种凋落物全碳质量分数为455~674 g·kg-1,m(DOC)∶m(C)为3.34%~8.48%。4个树种凋落叶在DOC、C、m(DOC)∶m(C)等初始值的变化上都是红椿>樟>桢楠>马尾松。N质量分数为红椿>桢楠>樟>马尾松。凋落物m(C)∶m(N)为马尾松>桢楠>樟>红椿,难降解物质质量分数(木质素和纤维素)红椿和樟的都显著低于马尾松和桢楠凋落叶。可见,3个阔叶树种的DOC、C、m(DOC)∶m(C)的初始值都高于针叶凋落物(马尾松),但马尾松凋落叶的m(C)∶m(N)则显著高于其他3种凋落叶。
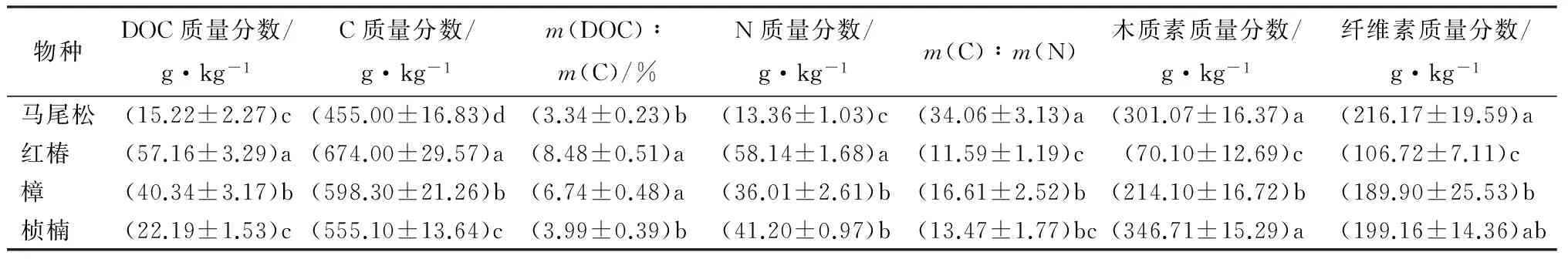
表4 4个树种凋落叶的初始基质状况
注:表中数值为“平均值±标准差”;同列不同的小写字母代表不同树种之间差异显著(P<0.05)。
3.3林窗面积对凋落物质量和DOC损失率的影响
林窗面积对各树种凋落物的质量损失有显著影响(P<0.05)。其中,G2、G3林窗的红椿凋落叶,G1、G4、G6林窗的樟凋落叶,G1、G3林窗的马尾松凋落叶,G2、G3、G4的桢楠凋落叶的质量损失率都显著高于其他林窗(见表5)。G1、G2、G3、G4林窗的红椿凋落叶DOC损失率显著高于其它林窗和林下,G2、G3林窗的樟凋落叶DOC损失率显著高于其它林窗和林下,G2、G3、G4林窗的马尾松凋落叶DOC损失率显著高于其它林窗和林下,G2、G4、G5林窗的桢楠凋落叶的DOC损失率显著高于其他林窗和林下。可见,相对于大面积(G6、G7)林窗,中小面积(G2、G3、G4)林窗内凋落物的DOC损失率更高,尤其是G2林窗,4种凋落叶的DOC损失率都高于其他林窗(见表6)。

表5 不同面积林窗内凋落叶的质量损失率
注:表中数值为“平均值±标准差”;同列不同大写字母表示不同面积林窗之间差异显著,同行不同小写字母表示同一面积林窗下不同凋落叶之间的差异显著(P<0.05)。
凋落物经过1 a的分解,质量损失率平均值为红椿(81.77%)、樟(48.99%)、桢楠(40.80%)、马尾松(38.07%)(见表5)。DOC损失率平均值分别为红椿(88.92%)、樟(83.04%)、桢楠(77.49%)、马尾松(59.30%)(见表6)。4个树种凋落物的质量损失率和DOC损失率均为红椿>樟>桢楠>马尾松。
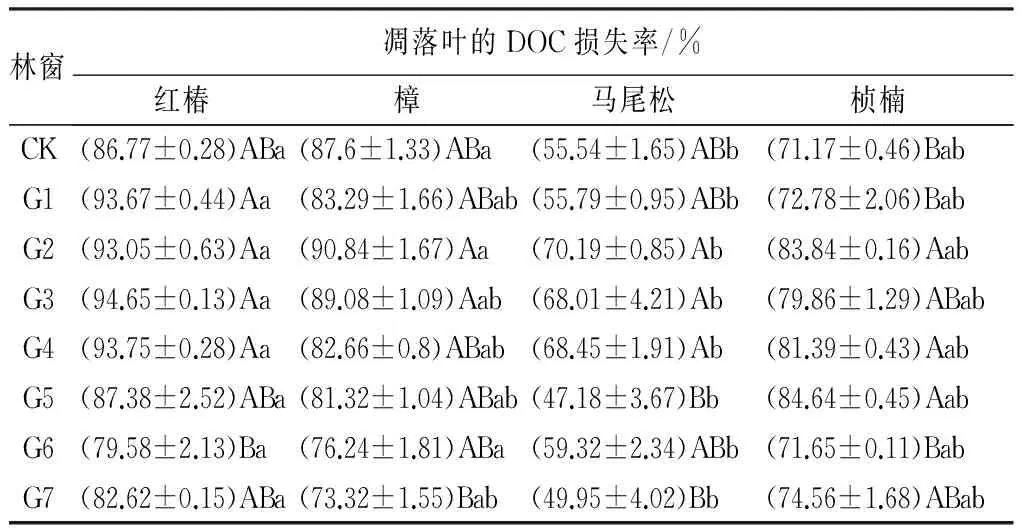
表6 不同面积林窗内凋落叶的DOC损失率
注:表中数值为“平均值±标准差”;同列不同大写字母表示不同面积林窗之间差异显著,同行不同小写字母表示同一面积林窗下不同凋落叶之间的显著差异(P<0.05)。
此外,从4个物种凋落物质量损失率和DOC损失率的平均值来看,不同面积林窗内,4种凋落叶DOC损失率与凋落叶质量损失率呈显著正相关(见图1),说明凋落物质量损失率越高其DOC损失率也相对更高。
3.44种凋落叶DOC损失率的差异
由表7可知,总体上看,分解初期(0~30 d)DOC损失率显著高于分解后期,后4个分解时期(>30~360 d),红椿凋落叶的DOC损失率高于其他3个树种凋落叶,而马尾松凋落叶的DOC损失率最低,樟和桢楠凋落叶的DOC损失率没有显著差异。分解初期(0~30 d),4种凋落叶DOC损失率都超过了10%,但树种之间无显著差异。分解360 d后,4种凋落叶之间的DOC总损失率,从大到小依次为红椿、樟、桢楠、马尾松。

图1 凋落叶DOC损失率与质量损失率的相关关系(n=120)

续(表7)
注:表中数值为“平均值±标准差”;同列不同大写字母表示同一分解时间、不同树种之间差异显著(P<0.05);同行不同小写字母表示同一树种、不同分解时间之间差异显著(P<0.05)。
3.5凋落叶DOC损失率的动态变化
由表8可知,不同树种凋落叶在不同分解阶段的DOC损失速率有所差异。马尾松、樟和桢楠凋落叶分解初期(0~30 d)DOC损失速率最高,而红椿凋落叶DOC损失速率最高值出现在0~90 d。之后,4个树种凋落叶的DOC损失速率均显著降低,且最后两个分解阶段(180~270 d与270~360 d)之间DOC损失速率无显著差异。凋落叶的DOC释放主要集中在分解初期(0~30 d)和分解前期(>30~90 d)。分解前期(>30~90 d)红椿凋落叶、分解中期(>90~180 d)马尾松凋落叶、分解中后期(>180~270 d)樟凋落叶的DOC损失率都显著高于同一分解阶段其他物种的DOC损失率。分解末期(>270~360 d)4种凋落叶的DOC损失速率无显著差异。说明随着分解时间的延长,凋落物的DOC损失速率不断降低,且不同凋落物之间的DOC损失速率的差异也不断减小。

表8 不同分解阶段4个树种凋落叶的DOC损失速率
注:表中数值为“平均值±标准差”;同列不同大写字母表示不同分解阶段之间差异显著(P<0.05),同行不用小写字母表示不同树种凋落叶之间的显著差异(P<0.05)。
4 结论与讨论
4.1林窗面积对凋落叶DOC损失率的影响
凋落叶作为森林凋落物最主要的组成部分,其可溶性有机碳的释放成为该地区土壤DOC来源及养分循环不可忽视的一个部分。众所周知,凋落物DOC质量分数和m(DOC)∶m(C)值可以反映凋落物DOC对土壤C库的贡献[34]。凋落物DOC释放主要决定于凋落物分解的温度、含水量和凋落物的化学性质[35]。本研究发现,林窗面积对凋落叶DOC损失率产生了显著影响,相对于大面积林窗,中小面积林窗内凋落物的DOC损失率更高。采样期间大面积林窗(G6、G7)的土壤温度都高于其他林窗,这与已有的研究结论“林窗面积越大,温度越高”一致[34]。这是因为相对于其他林窗下,大面积林窗内光照更强。研究发现,温度较低的中小面积林窗,DOC损失率更高。这与邓长春等[32]的研究结论一致。凋落叶的DOC损失率和含水率为正相关,凋落物中的DOC容易被水溶解出来。研究区域属中亚热带湿润季风气候,年均降水量1 070.4 mm,雨量充沛,有利于DOC的释放[36]。土壤含水率随林窗面积增大,先升高后降低。这是因为研究区域内马尾松平均株高大于16 m,阻挡了降雨,对小林窗以及林下(CK)的土壤湿度等条件的改善有所抑制[37]。而大面积林窗,由于增强了林窗内的光照强,增大了蒸发,林窗内土壤含水率降低[38]。因此,相对于其他林窗和林下,含水率较高的中小面积林窗更有利于凋落叶DOC的释放。可溶性有机碳主要包括糖类、氨基酸类淀粉类及低分子量的有机物等物质,这些物质极容易被微生物分解和利用[39]。欧江等[37]研究发现,微生物生物量总体上随林窗增大先升后降,且中小面积林窗土壤有机碳质量分数的季节差异更小,能够更长时间维持分解者的生存。Q. ZHANG et al.[40]也指出,直径大于30 m的大林窗土壤微生物生物量显著减少。ARUNACHALAM et al.[41]的研究表明,土壤细菌数量在面积为306.9~981.8 m2的大林窗较高。另外,土壤微生物的种类、数量、分布与地上植被群落的结构有着密切的关系[42]。林窗形成后草灌层先锋植物(如商陆、野桐等)迅速萌发生长,且与大面积林窗相比,较小林窗更利于芒其、蕨类等植物生存。植物盖度、种类等较高的林窗,将为秋季的微生物提供更丰富的凋落物等碳、氮、磷的来源[37]。MUSCOLO et al.[43]指出因更高的养分有效性和更多的腐殖质,微生物量小林窗(380 m2)>中林窗(855 m2)>大林窗(1 520 m2)。由此可见,微生物量更高的中小面积林窗,更能促进凋落叶中DOC的释放。
4.2不同物种凋落叶DOC损失率的差异
本研究中,4种植物凋落物中可溶性有机碳的质量分数(15.22~57.16 g/kg)及其占凋落物全碳的比例(3.34%~8.48%)差异较大。其中,3种阔叶树种(红椿、樟、桢楠)的DOC质量分数和m(DOC)∶m(C)比都高于针叶树种(马尾松)。经过1 a的分解,红椿、樟、桢楠、马尾松不同凋落叶的质量损失率分别为81.88%,49.37%,41.41%,39.04%,红椿、樟、桢楠、马尾松的DOC损失率分别为:90.06%,85.17%,78.74%,61.55%。说明凋落物分解过程中,可溶性有机碳释放与质量损失密切相关[44]。4种凋落叶中,红椿凋落叶的DOC损失率最高,马尾松最低。这可能是因为不同凋落物之间m(C)∶m(N)的差异造成的。4种凋落叶的初始m(C)∶m(N)比为红椿(11.14)、桢楠(13.47)、樟(17.19)、马尾松(34.06)。凋落物分解需要一个适宜的m(C)∶m(N)比(一般低于25)[45],m(C)∶m(N)比越低或含N越高,越有利于其分解[46-47]。而马尾松的m(C)∶m(N)比(34.06)显著高于其他3个阔叶树种。其次,凋落叶的DOC释放与其本身的质地和结构有关。由于马尾松和桢楠凋落叶较红椿和樟凋落叶质地更硬,土壤动物、微生物以及淋溶作用对其影响相对减弱,而红椿、樟则因为质地较柔软,容易受土壤动物、微生物的影响和降雨的淋溶作用使其DOC释放得更快[48]。
4.3凋落叶DOC损失率的动态变化
4种凋落物的DOC损失速率都呈先快后慢的变化趋势,这与李云[44]、TIETEMA et al.[49]的研究结果一致。由于研究区域属中亚热带湿润季风气候,年均降水量1 070.4 mm左右,雨量充沛[50],在凋落叶分解的过程中,初期DOC流失较快,使得凋落物中DOC质量分数越来越低,导致分解后期淋溶作用对凋落物的作用减弱,DOC淋失量降低[51]。同时,随着分解时间的延长,木质素、纤维素等难降解物质的相对质量分数上升,影响土壤动物、土壤微生物对凋落叶的分解,也会影响DOC的释放[52]。凋落物在分解初期损失大量的有机碳,大部分是以DOC的形式流失。随着分解时间的变化,凋落物不断分解增加了土壤中DOC的质量分数,减小了凋落物和土壤的DOC的质量分数,导致凋落物的DOC损失率降低[53]。随着气温回升,林窗内的土壤动物、微生物开始活跃,加快对可溶性物质的分解。丘清燕等[54]、张剑等[55]研究发现,DOC化学组成不同,导致微生物对其降解速率也存在差异,微生物可快速降解分解初期新鲜凋落物释放的DOC,但是对凋落物分解后期释放的DOC降解能力减弱。
综上所述,人工干扰对凋落叶的分解过程中DOC释放有显著影响,中小面积林窗较大面积林窗能显著提高凋落叶DOC释放。4种凋落叶的DOC释放都遵循先快(0~90 d)后慢的变化规律,且3个阔叶树种(红椿、樟、桢楠)的DOC损失率高于针叶树种(马尾松)凋落叶。因此,在中小面积林窗加快凋落物DOC释放的基础上,引入更易分解的乡土阔叶树种(红椿、樟、桢楠)可促进森林生态系统中凋落叶DOC释放,从而更有利于马尾松人工林有机碳的循环利用。这为研究林分干扰调节森林生态系统凋落物养分释放提供参考,也为深入理解长江上游低山丘陵区人工林生态系统物质循环过程提供了一定的基础数据。
致谢:感谢四川农业大学的张丹桔、高顺、陈良华等老师和宋小艳、王珂等同学在实验方面对本文作者提供的帮助与指导。
[1]ROSS D J, TATE K R, NEWTON P C D, et al. Carbon mineralization in an organic soil, with and without added grass litter, from a high-CO2environment at a carbon dioxide spring[J]. Soil Biology and Biochemistry,2003,35(12):1705-1709.
[2]KELLIHER F M, ROSS D J, LAW B E, et al. Limitations to carbon mineralization in litter and mineral soil of young and old ponderosa pine forests[J]. Forest Ecology and Management,2004,191(1):201-213.
[3]MAGILL A H, ABER J D. Variation in soil net mineralization rates with dissolved organic carbon additions[J]. Soil Biology and Biochemistry,2000,32(5):597-601.
[4]杨玉盛,郭剑芬,陈光水,等.森林生态系统DOM的来源、特性及流动[J].生态学报,2003,23(3):547-558.
[5]陶澍,陈静生,邓宝山,等.中国东部主要河流河水腐殖酸的起源、含量及地域分异规律[J].环境科学学报,1998,8(3):286-294.
[6]陶澍,崔军,张朝生.水生腐殖酸的可见—紫外光谱特征[J].地理学报,1990,45(4):484-489.
[7]CURRIE W S, ABER J D. Modeling leaching as a decomposition process in humid mountain forest[J]. Ecology,1997,78(6):1844-1860.
[8]DALVA M, MOORE T R. Sources and sinks for dissolved organic carbon in a forested swamp catchment[J]. Biogeochemistry,1991,15(1):1-19.
[9]ANDERSSON S, NILSSON S I, SSETRE P. Leaching of dissolved organic carbon and dissolved organic nitrogen in mot humus as affected by temperature and pH[J]. Soil Biology and Biochemistry,2000,32(1):1-10.
[10]STROBEL B W, HANSEN H C B, BORGGAARD O K. Composition and reactivity of DOC in forest floor soil solutions in relation to tree species and soil type[J]. Biogeochemistry,2001,56(1):1-26.
[11]田大伦,宁晓波.不同龄组马尾松林凋落物量及养分归还量研究[J].中南林学院学报,1995,15(2):163-169.
[12]雷加富.中国森林资源[M].北京:中国林业出版社,2005.
[13]庞学勇,刘庆,刘世全,等.川西亚高山云杉人工林土壤质量性状演变[J].生态学报,2004,24(2):261-267.
[14]胡蓉,林波,刘庆.林窗与凋落物对人工云杉林早期更新的影响[J].林业科学,2011,47(6):23-29.
[15]陈梅,张愈佳.不同大小林窗生长季初期小气候特征比较:以辽东山区次生林生态系统林窗为例[J].内蒙古农业科技,2011(3):78-82.
[16]GRAY A N, SPIES T A. Gaps size, within-gap position and canopy structure effects on conifer seeding establishment[J]. Journal of Ecology,1996,84(5):635-645.
[17]KENNETH P L. Pattern of gap-phase replacement in a subalpine, old growth forest[J]. Ecology,1992,73(2):657-669.
[18]SHUGART H H. A theory of forest dynamics: the ecological implications of forest succession models[M]. New York: Springer-Verlag,1984.
[19]SCHWARTZ G, LOPES J C A, MOHREN G M J, et al. Post-harvesting silvicultural treatments in logging gaps: A comparison between enrichment planting and tending of natural regeneration[J]. Forest Ecology and Management,2013,293(2):57-64.
[20]WHITMORE T C. Canopy gaps and the two major groups of forest trees[J]. Ecology,1989,70(3):536-538.
[21]OSADA N, TAKEDA H K, PEARCY R W. Functional correlates of leaf demographic response to gap release in saplings of a shade-tolerant tree,Elateriospermumtapos[J]. Ecophysiology Oecologia,2003,137(2):181-187.
[22]RENTCH J S, SCHULER T M, NOWACKI G J, et al. Canopy gap dynamics of second-growth red spruce-northern hardwood stands in west virginia[J]. Forest Ecology and Management,2010,260(10):1921-1929.
[23]KERN C C, REICH PB, MONTGOMERY R A, et al. Do deer and shrubs override canopy gap size effects on growth and survival of yellow birch, northern red oak, eastern white pine, and eastern hemlock seedlings?[J]. Forest Ecology and Management,2012,267(3):134-143.
[24] 徐李亚,杨万勤,李晗,等.长江上游高山森林林窗对凋落物分解过程中可溶性碳的影响[J].长江流域资源与环境,2015,24(5):882-891.
[25]KALBITZ K, SOLINGER S, PARK J H, et al. Control on the dynamics of dissolved organic matter in soils: A review[J]. Soil Science,2000,165(4):277-304.
[26]杨玉盛,林瑞余,李庭波,等.森林凋落物淋溶中的溶解有机物与紫外:可见光谱特征[J].热带亚热带植物学报,2004,12(2):124-128.
[27]ZSOLNAY A. Dissolved organic matter: Artifacts, definitions, and functions[J]. Geoderma,2003,113(3/4):187-209.
[28]BOYER J N, GROFFMAN P M. Bioavailability of water extractable organic carbon fractions in forest and agricultural soil profiles[J]. Soil Biology and Biochemistry,1996,28(6):783-790.
[29]MURPHY D V, MACDONALD A J, STOCKDALE E A, et al. Soluble organic nitrogen in agricultural soils[J]. Biology and Fertility of Soil,2000,30(5):374-387.
[30]王春阳,周建斌,王祥,等.黄土高原区不同植物凋落物可溶性有机碳的含量及生物降解特性[J].环境科学,2011,32(4):1139-1145.
[31]SWAN C M. Methods to study litter decomposition: a practical guide[J]. Journal of the North American Benthological Society,2007,26(2):363-364.
[32]邓长春,陈亚梅,和润莲,等.雪被期川西高山林线交错带凋落物可溶性有机碳的淋失特征[J].生态学杂志,2014,33(11):2921-2929.
[33]李正才,徐德应,杨校生,等.北亚热带6种森林类型凋落物分解过程中有机碳动态变化[J].林业科学研究,2008,21(5):675-680.
[34]ZHU J J, TAN H, LI F Q, et al. Microclimate regimes following gap formation in a montane secondary forest of eastern liaoning province, china[J]. Journal of Forestry Research,2007,18(3):167-173.
[35]MORETTO A S, DISTEL R A, DIDON N G. Decomposition and nutrient dynamic of leaf litter and roots from palatable and unpalatable grasses in a semi-arid grassland[J]. Applied Soil Ecology,2001,18(1):31-37.
[36]窦荣鹏,江洪,余树全,等.柳杉凋落物在中国亚热带和热带的分解[J].生态学报,2010,30(7):1758-1763.
[37]欧江,张捷,崔宁洁,等.采伐林窗对马尾松人工林土壤微生物生物量的初期影响[J].自然资源学报,2014,29(12):2036-2047.
[39]吴庆标,王效科,欧阳志云.活性有机碳含量在凋落物分解过程中的作用[J].生态环境,2006,15(6):1295-1299.
[40]ZHANG Q, ZAK J C. Effects of gap size on litter decomposition and microbial activity in a subtropical forest[J]. Ecology,1995,76(7):2196-2204.
[41]ARUNACHALAM A, ARUNACHALAM K. Influence of gap size and soil properties on microbial biomass in a subtropical humid forest of north-east India[J]. Plant & Soil,2000,223(1):187-195.
[42]孟好军,刘贤德,金铭,等.祁连山不同森林植被类型对土壤微生物影响的研究[J].土壤通报,2007,38(6):1127-1130.
[43]MUSCOLO A, SIDARI M, MERCURIO R. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinuslaricio,Poiret) stands[J]. Forest Ecology and Management,2007,242(2/3):412-418.
[44]李云,周建斌,董燕捷,等.黄土高原不同植物凋落物的分解特性[J].应用生态学报,2012,23(12):3309-3316.
[45]田茂洁.川中人工纯柏林凋落物分解动态研究[J].生态学杂志,2005,24(10):1147-1150.
[46]王瑾,黄建辉.暖温带地区主要树种叶片凋落物分解过程中主要元素释放过程的比较[J].植物生态学报,2001,25(3):375-380.
[47]SCHIMEL J P, WEINTRAUB M N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: a theoretical model[?]. Soil Biology & Biochemistry,2003,35(4):549-563.
[48]王文君,杨万勤,谭波,等.四川盆地亚热带常绿阔叶林土壤动物对几种典型凋落物分解的影响[J].生态环境学报,2013,22(9):1488-1495.
[49]TIETEMA A, WESSEL W W. Microbial activity and leaching during initial oak leaf litter decomposition[J]. Biology and Fertility of Soils,1994,18(1):49-54.
[50]崔宁洁,刘洋,张健,等.林窗对马尾松人工林植物多样性的影响[J].应用与环境生物学报,2014,20(1):8-14.
[51]康根丽,杨玉盛,司友涛,等.马尾松与芒萁鲜叶及凋落物水溶性有机物的溶解特征和光谱学特征[J].热带亚热带植物学报,2014,22(4):357-366.
[52]胡肄慧,陈灵芝,孔繁志,等.油松和栓皮栋枯叶分解作用的研究[J].植物学报,1986,(28):102-110.
[53]雷秋霜,杨秀虹,方志文,等.森林新近凋落叶溶出DOM的性质及其对菲增溶作用的影响[J].生态环境学报,2014,23(1):170-177.
[54]丘清燕,梁国华,黄德卫,等.森林土壤可溶性有机碳研究进展[J].西南林业大学学报,2013,33(1):86-96.
[55]张剑,汪思龙,王清奎,等.不同森林植被下土壤活性有机碳含量及其季节变化[J].中国生态农业学报,2009,17(1):41-47.
Effect of Forest Gap Size on Loss of Dissolved Organic Carbon (DOC) for Four Leaf Litters//
Li Xun, Liu Yang, Zhang Jian, Yang Wanqin, Zhang Yan, Deng Changchun, Zhang Mingjin, Liu Hua
(Sichuan Agricultural University, Chengdu 611130, P. R. China)//Journal of Northeast Forestry University,2016,44(1):23-30.
Dissolved organic carbon (DOC); Forest gap size;Pinusmassonianaplantation;Pinusmassonianaleaf litters
李勋,男,1990年2月生,四川农业大学林学院,硕士研究生。E-mail:502780405@qq.com。
张健,四川农业大学林学院,研究员。E-mail:sicauzhangjian@163.com。
2015年7月24日。
S725.79
1)国家自然科学基金项目(31370628);国家科技支撑计划项目(2011BAC09B05);四川省科技支撑计划项目(12ZC0017);四川省科技应用基础项目(2012JY0047);四川省教育厅科技创新团队计划项目(11TD006) 。
责任编辑:王广建。
