贵阳地区下三叠统安顺组遗迹化石及古环境意义
2016-08-15余关美
余关美 时 国
(东华理工大学地球科学学院 南昌 330013)
贵阳地区下三叠统安顺组遗迹化石及古环境意义
余关美时国
(东华理工大学地球科学学院南昌330013)
贵阳地区下三叠统安顺组中的遗迹化石是研究P-T生物大灭绝后古环境、古生态的良好材料,在研究区发现、描述了8个遗迹属,9个遗迹种。安顺组中遗迹化石的垂向分布表明:安顺组一段以结构单一、个体较小的遗迹化石Planolitesmontanus,Palaeophycuscurvatus和Phycodespalmatus为主;二、三段中遗迹化石属种增多、生物扰动增强,除了一段的3个遗迹种外还发现有6个遗迹种:Beaconichnusdarwinum,Chondritesrecurvus,Thalassinoidesparadoxicus,Thalassinoideshorizontalis,Arenicolitesisp.,Rhizocoralliumcommune,在三段出现的复杂遗迹化石Rhizocoralliumcommune和Thalassinoidesparadoxicus代表了本地区海洋环境在早三叠世晚期的重大改善。岩性特征和生物化石特征表明安顺组一段上部为台缘浅滩环境,二段为碳酸盐岩台地边缘斜坡环境,三段为靠近台地边缘的潮间—潮下环境,在一至三段沉积期,海水含氧条件有所改善,研究区处于浅海“宜居带”上。安顺组四段沉积于半局限台地内的潮间—潮上环境,处于“宜居带”之外,在此区域遗迹化石不发育,生物活动相对较弱。早三叠世奥伦尼克期遗迹化石发育特征表明生物复苏活动在碳酸盐岩台地边缘带相对活跃。
遗迹化石安顺组沉积环境宜居带生态复苏
二叠纪末的生物大灭绝事件使海洋生态系统受到极大破坏[1],火山活动导致的温室气体快速释放和全球极热化使生态系快速崩溃[2],其后的生物复苏经历了4~8 Ma的延续,直到中三叠世海洋生态系统才开始恢复到灭绝前的状态[3-5]。近年来越来越多的遗迹化石研究表明在早三叠世生物复苏活动业已开始[5-17]。贵阳地区下三叠统遗迹化石丰富,保存完整[5-6],与该地区中三叠世早期的“青岩生物群”[18]具有沉积上的连续性,为研究P-T事件后的环境变化与生物复苏提供了良好的材料。本文以该地区早三叠世奥伦尼克期安顺组中的遗迹化石为研究材料,结合岩性特征,分析安顺组的沉积环境,并初步讨论了遗迹化石在早三叠世生物复苏过程中的意义。
1 地层和遗迹化石
早三叠世贵阳花溪地区位于扬子台地西南缘(图1a),发育一套浅水碳酸盐岩台地—深水碎屑岩盆地过渡相沉积[9],自下而上为大冶组和安顺组。大冶组主要为浅灰色薄层石灰岩与薄板泥质石灰岩;安顺组发育白云质石灰岩、白云岩以及溶塌角砾岩,覆于大冶组石灰岩之上,整合于中三叠世关岭组白云岩之下,该组主要分布于清镇、贵阳、花溪一带(图1b)。牙形石Neogondolellapingdingshanensis和N.waageni的出现,可将安顺组地质时代界定为奥伦尼克期[18-19],组内已发现的Dienerocerassp.、Pseudowenitesoxynostus等菊石类,以及Eumorphotis-Entoliumdiscites-Unionites、Pteriacf.murchisoni-Bakevellia等瓣鳃类组合也显示了奥伦尼克期的特征[20]。根据岩性组合的差异,安顺组自下而上可以分为四段(图2)[20],各段所发现的遗迹化石特征简单描述见表1。

图1 贵阳地区奥伦尼克期地层分布(a据Lehrmann et al., 2003,修改)Fig.1 Olenekian facies distribution in Guiyang area (a is modified from Lehrmann et al.,2003)
安顺组一段由浅灰色中层状、藻纹层状白云岩夹少量白云质石灰岩构成,缝合线发育,生物化石稀少,偶见瓣鳃类Leptochondriacf.bittneri及腕足类Lingulasp.等[20]。该段遗迹化石发育较少,下部发现Planolitesmontanus(图3A),上部发现Phycodespalmatus(图3C)和Palaeophycuscurvatus(图3I)2个遗迹种。
安顺组二段由灰白色结晶白云岩夹薄层泥岩构成,偶夹薄层细晶白云岩,发育鸟眼构造和交错层理,富含瓣鳃类Unionitescanalensis,Claraiasp.等[20]。该段遗迹化石丰富,除Planolitesmontanus,Palaeophycuscurvatus和Phycodespalmatus之外,还发现了Beaconichnusdarwinum(图3D),Thalassinoidesparadoxicus(图3E),Chondrites recurvus(图3G)和Planolitesbeverleyensis等4个遗迹种。
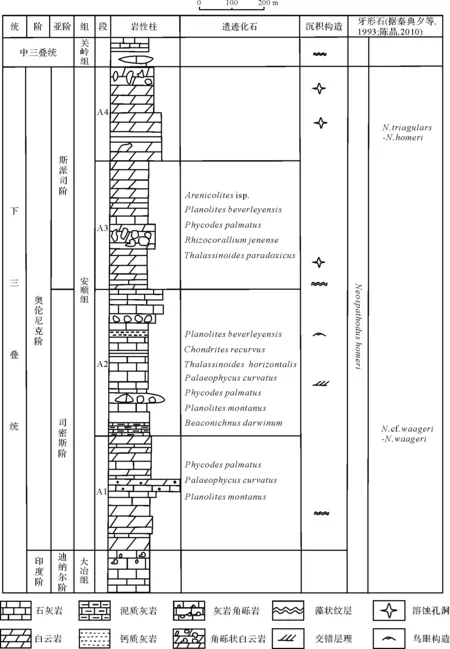
图2 贵阳地区下三叠统安顺组综合柱状图Fig.2 Synthetic column of the Lower Triassic Anshun Formation in Guiyang, Guizhou province

遗迹属简单描述习性类型造迹生物Planolitesmontanus直或略弯曲的水平简单潜穴,不分枝;潜穴深度在2mm以内,表面光滑,潜穴直径2~4mm。觅食迹蠕虫类Palaeophycuscur-vatus圆柱形或亚柱形潜穴,潜穴直或微弯曲,潜穴直径2~4mm,表面光滑,不分枝,潜穴具衬里。觅食迹多毛类、蠕虫类Phycodespalmatus整体呈掌状或扫帚状,管面光滑,每个潜穴管为圆柱状,只发生一次分叉,潜穴管分支直径为3~5mm;分支角度在15o~30o之间。觅食迹蠕虫类环节动物、海鳃动物Chondritesrecurvus洞穴表面上与植物的根相似,它由中心体管构成,从中心体管的基部辐射出众多的细小分枝。觅食迹蠕虫类Beaconichnusdarwi-num光滑简单沟槽状遗迹,由两条窄的沟槽组成,不分支,相互之间交切,遗迹宽度在5mm左右。爬行迹节肢动物Rhizocoralliumcommune具有清楚蹼状构造的U形管,顶部呈半圆形,管壁光滑,管内直径3~4mm,宽约8~9mm,两翼管与层面从斜交30o~45o至近平行于层面。居住觅食迹甲壳动物、多毛类、蠕虫类Thalassinoidespar-adoxicus分枝系统不规则,水平潜穴发达,Y形或垂直分枝,潜穴直径4~8mm,潜穴直径在分枝结合点肿大。居住觅食迹甲壳动物Thalassinoideshor-izontalis三维潜穴系统,在水平面上呈网状分支构造,分支呈Y型或T型,潜穴直径3~7mm,整体上直径变化不大,在分支处稍有膨大现象,分枝间距较为均一。居住觅食迹迹甲壳动物Arenicolitesisp.简单无蹼状构造的U型潜穴,U形管垂直于层面,管径1~2mm,两管间距为5~15mm,被动充填。居住迹蠕虫类环节动物、小型甲壳动物
安顺组三段主要由灰色薄至中层状白云岩夹泥质白云岩构成,藻状纹层和溶蚀孔洞发育,含瓣鳃类Entolium discites、Pteriacf. murchisoni等、菊石Proptychitoidessp.、有孔虫Amonodiscussp.等[20]。发现有Planolites beverleyensis,Phycodes palmatus,Rhizocorallium commune(图3B),Arenicolitesisp.(图3F)和Thalassinoides pradoxicus(图3H)5个遗迹种。
安顺组四段由紫红色中至厚层状白云岩构成,时夹盐溶角砾岩,发育大量次生孔洞和鸟眼构造,含瓣鳃类Myophoriasp.等、有孔虫、腹足类等[20-21]。该段偶见生物扰动构造,未见明显的遗迹化石。
2 遗迹化石发育特征
遗迹化石在地层中的丰度与分异度、生物扰动强度、潜穴大小及其复杂度等特征是研究造迹生物底栖生态环境变化的重要指标[5,10-12,15,22-27],这些特征对推断研究区早三叠世奥伦尼克期遗迹化石的演变及沉积环境的变化有重要作用。
2.1遗迹化石的丰度与分异度
安顺组一段底部未见遗迹化石,上部开始出现少量遗迹化石,该段遗迹化石的丰度和分异度都较低(图4),仅发现有3个遗迹属,反映出造迹生物类型单调[24];安顺组二段相比一段增加了4个遗迹属,迹化石丰度和分异度开始增大,表明造迹生物生态习性由简单向复杂多样化发展[24];安顺组三段遗迹化石丰度和分异度仍然较高,出现了2个新的遗迹种,反应了古生态环境的极大改善;安顺组四段未见明显的遗迹化石。
对单个属而言,Palaeophycus和Planolites 在岩层中的丰度较大,表示这一时期沉积物中的含氧量较低[26]。

图4 贵阳地区安顺组中遗迹化石的垂向分布Fig.4 Vertical distribution of ichnofossils in Anshun Formation in Guiyang area
2.2生物扰动指数
生物扰动指数指整个沉积物受生物扰动程度的等级,据扰动量在整个沉积物中所占的百分比将其划分为7个(0~6级)等级[28-29]。安顺组一段生物扰动指数为1~2,扰动较弱;安顺组二段生物扰动增强,扰动指数为3~4,最高达到5;安顺组三段生物扰动仍然强烈,扰动指数为3~4;安顺组四段生物扰动指数逐渐减小。
2.3潜穴大小和深度
遗迹化石的大小是古环境恢复的重要指示,小型的遗迹化石,通常出现在不含氧、半咸水环境的栖息地或低养分供应的沉积物中[3,30]。以贯穿安顺组一段到三段的Planolites为例来比较遗迹化石潜穴大小的变化,安顺组一段的Planolites潜穴直径3~6.5mm,平均值4.4mm;安顺组二段潜穴直径迅速增大,集中分布在6.5~13.5mm之间,平均值12.5mm;潜穴直径平均值最大值出现在安顺组三段,为11.8mm,潜穴直径最大为16mm。与大冶组相比[5],安顺组一段潜穴直径明显减小,随后潜穴直径快速增加,之后潜穴直径变化幅度变小。
安顺组中遗迹化石造迹生物多为蠕虫类,其潜穴深度一般较低,安顺组二、三段出现了Rhizocorallium和Thalassinoides等相对复杂的遗迹化石,其造迹生物为甲壳纲、多毛类或蠕虫类[31-33],活动能力较强,潜穴深度相对较大。安顺组中的Rhizocorallium潜穴深度主要为12~22mm,平均值达15mm,与大冶组[5]中的Rhizocorallium相比,安顺组中的Rhizocorallium潜穴深度的平均值明显增大,表明了安顺组中遗迹化石较大冶组有更强的生物扰动能力。
2.4复杂度与形态演化
遗迹化石复杂度的变化不仅表现在遗迹属种的分异度上,还表现在遗迹本身的形态结构上[5]。安顺组一段的遗迹化石Planolites 和Palaeophycus小而单一,随着安顺组二段遗迹化石丰度和分异度的增大,其复杂度也开始增加,对比安顺组一段沿层面分布的简单遗迹,具蹼状构造的Rhizocorallium和呈网状的Thalassinoides都表现了遗迹化石复杂度的增加。在造迹生物方面,Planolites 和Palaeophycus等遗迹化石多为多毛类蠕虫在沉积物表面食沉积物造成[5],而Rhizocorallium和Thalassinoides则是由甲壳纲、多毛类或蠕虫类造成,表明生物的高级化有所发展,同时其改造沉积物的能力也增强,形成的遗迹化石形态也更加复杂。
3 讨论
3.1沉积环境演化
研究区安顺组一段整合于大冶组碳酸盐岩台地边缘斜坡相灰岩角砾岩之上,发育藻纹层状白云岩夹少量白云质石灰岩,偶见鲕粒白云岩,缝合线发育,具藻状纹层构造,该段生物稀少,少见遗迹化石,扰动作用较弱,上述特征表明在大冶组碳酸盐岩斜坡沉积之后[34],本区水体进一步变浅,说明安顺组一段应为台缘浅滩相沉积。安顺组二段由微晶白云岩夹薄层泥岩构成,偶夹薄层细晶白云岩,常见交错层理,夹少量杂基支撑的灰岩角砾岩凸镜体,该段遗迹化石丰富且分异度较大,结合岩性、沉积构造等特征分析表明水体进一步加深,该段应属台缘斜坡相沉积。安顺组三段主要由灰色薄至中层状白云岩夹泥质白云岩构成,发育藻状纹层和溶蚀孔洞,含瓣鳃类、菊石、有孔虫等生物化石,遗迹化石丰富,生物扰动强烈,综合岩性、沉积构造、古生物化石等特征分析,表明该段水体逐渐变浅,属于靠近台地边缘的底内生物繁盛的潮间—潮下沉积环境[35]。安顺组四段由紫红色中至厚层状白云岩构成,时夹盐溶角砾岩,发育大量次生孔洞和鸟眼构造,含瓣鳃类、有孔虫、腹足类等[20],未发现遗迹化石,但存在生物扰动,表明该时期环境变化大,水体进一步变浅,应为半局限台地内的潮间—潮上带沉积环境。上述沉积环境的变化表明早三叠晚期研究区水体经历了由深到浅的过程,总体为浅海环境。
Beatty等学者(2008)在研究泛大陆西北部早三叠世遗迹化石多样性时提出了“宜居带”这一概念,是指二叠世末生物大灭绝后早三叠世时期形成的一个适宜海底生物生存的区域(图5),该区域位于风暴浪基面之上,其特征是具有较大生物扰动指数和遗迹化石多样性。在“宜居带”以上的区域,因为波动太强烈,不适宜遗迹化石的保存,在风暴浪基面以下的区域难以获得足够的氧而不适宜生物生存[10]。最易形成“宜居带”的是正常天气浪基面与风暴浪基面间的区域,此区域风浪作用能使海水充氧,利于海底生物生存[10,36]。印度期末全球环境发生的较大变化[5]以及研究区海平面快速下降[34]致使印度期生物复苏活动结束,同时使奥伦尼克期早期环境压力较大。安顺组一段处于台缘浅滩环境,上部遗迹化石开始发育,环境趋于稳定,生态环境得到改善,适宜底栖生物生存,安顺组二段和三段的遗迹化石多样性大,生物扰动强烈,此时期海水环境含氧量有所增加,结合前文的沉积环境分析,说明处于碳酸盐岩台地边缘的“宜居带”环境为遗迹化石在大冶组之后的再次复苏[5]和演化提供了有利条件。研究区四段处于台地内部,沉积于“宜居带”之上的区域,未见大量遗迹化石。

图5 宜居带位置(据Beatty et al.,2008; Pietsch et al.,2014修改)Fig.5 A diagram of the habitable zone (modied from Beatty et al., 2008; Pietsch et al., 2014)
3.2对早三叠世生物复苏的启示
安顺组一段遗迹化石发育较少,且该段生物稀少,偶见双壳类及腕足类,遗迹化石多样性低,以小而单一的水平潜穴Planolites 和Palaeophycus为主,遗迹属习性以觅食迹为主,结合潜穴直径较小和扰动作用微弱等特征,说明研究区奥伦尼克期早期海水含氧量较低[23],反应了P-T危机之后持续较长时间的环境压力。与安顺组一段对比,安顺组二段和三段的遗迹化石多样性、潜穴大小明显增大,表明沉积物中含氧量有所增高,在这期间,实体化石也较丰富,富含瓣鳃类,腹足类、菊石、有孔虫也较常见[20],加之强烈的生物扰动构造,表明这一时期的海洋古生态条件得到了极大的改善,底栖生物多样性增大,安顺组二段顶部和三段底部出现的复杂遗迹化石Rhizocorallium commune和Thalassinoides paradoxicus反映了生态系统恢复到了一个较高的程度[37]。研究区安顺组遗迹化石的特征变化显示出由习性构造较为简单、分异度较低的复苏期进入习性构造复杂、分异度较高的辐射期,其演变模式与大冶组[5-6,8]时期相似,且表现出的环境转变模式与贵州独山晚泥盆世弗拉期一法门期灭绝界线上部遗迹化石所表现的造迹生物的演化特征相似[38],表明在经历了P-T大灭绝之后,遭到破坏的生态系统在早三叠世开始重建。大冶组上部至安顺组下部遗迹化石消失,说明早三叠世早期建立起来的生态系统较脆弱,未能存活下来,故而安顺组时期遗迹化石与生物化石变化特征指示的是早三叠世的第二次复苏。
贵阳地区及全球其他地区奥伦尼克期的遗迹化石及古生物演化特征研究表明,该时期全球海洋古生态条件得到了极大的改善,底栖生物多样性增大。如位于上扬子北缘的四川地区[25-26]奥伦尼克期遗迹化石多样性增加,潜穴直径增大,且有强烈的生物扰动构造,说明此时海底环境有了明显的改善。与此同时,南盘江盆地海百合类逐渐丰富[39],扬子板块下部的海洋爬行类(ichthyopterygia)实体化石也开始出现。巴基斯坦的Salt地区[39],奥伦尼克期双壳类多样性增大;意大利Bulla地区[35]奥伦尼克期早期遗迹化石多样性、生物扰动指数、遗迹大小等相对较小,随后又开始增大。遗迹化石和实体化石的变化特征表明生物在奥伦尼克期已经开始复苏,但在该时期复苏并未完成,生态系统并不完善,为一个稳定的“初级生态系统”。
4 结论
(1) 安顺组发育遗迹化石有:Planolites montanus,Palaeophycus curvatus,Phycodes palmatus,Beaconichnus darwinum,Thalassinoides horizontalis,Arenicolitesisp.,Chondrites recurvus,Rhizocorallium commune,Thalassinoides paradoxicus。
(2) 沉积环境分析表明,研究区安顺组沉积环境自下而上经历了第一段的台缘浅滩,第二段的台缘斜坡,到第三段的潮间—潮下带,再到第四段的潮间—潮上带的演变过程。安顺组的沉积环境演化表明生物复苏活动在碳酸盐岩台地边缘带相对活跃。
(3) 贵阳地区安顺组遗迹化石的特征变化体现出了海洋生态环境的演化特征。安顺组一段仅发现3个遗迹属,分异度较低,说明研究区奥伦尼克期早期沉积物中的含氧量较低,古生态条件处于较高的压力环境下;安顺组二段和三段遗迹属种增多,分异度增大,表明这一时期海水的含氧量有所增加,环境条件由高压力向正常转变,海洋古生态条件有了明显的改善,生态系统恢复到一个较高的程度。
致谢感谢编辑老师和审稿专家对论文提出的宝贵意见。
References)
1戎嘉余,黄冰. 生物大灭绝研究三十年[J]. 中国科学:地球科学,2014,44(3):377-404. [RongJiayu,HuangBing.StudyofMassExtinctionoverthepastthirtyyears:asynopsis[J].ScientiaSinicaTerrae, 2014, 44(3): 377-404. ]
2SunYadong,JoachimskiMM,WignallPB,etal. 2012.LethallyHotTemperaturesduringtheEarlyTriassicGreenhouse[J].Science, 338(6105): 366-370.
3PrussSB,BottjerDJ.EarlyTriassictracefossilsofthewesternUnitedStatesandtheirimplicationsforprolongedenvironmentalstressfromtheend-Permianmassextinction[J].PALAIOS, 2004, 19(6): 551-564.
4童金南,殷鸿福. 早三叠世生物与环境研究进展[J]. 古生物学报,2009,48(3):497-508. [TongJinnan,YinHongfu.AdvanceinthestudyofEarlyTriassiclifeandenvironment[J].AtcaPalaeontologicaSinica, 2009, 48(3): 497-508. ]
5罗茂,时国,龚一鸣. 贵阳花溪早三叠世遗迹化石及其对二叠纪末生物大灭绝事件后生物复苏的启示[J]. 古地理学报,2007,9(5):519-532. [LuoMao,ShiGuo,GongYiming.EarlyTriassictracefossilsinHuaxiregionofGuiyangandtheirimplicationsforbioticrecoveryaftertheend-Permianmassextinction[J].JournalofPalaeogeography, 2007, 9(5): 519-532. ]
6王尚彦. 贵阳市花溪地区下三叠统大冶组中遗迹化石及沉积环境[J]. 贵州地质,1987,4(4):454-460. [WangShangyan.TracefossilsandtheirsedimentaryenvironmentsinDayeFormationLowerTriassicofHuaxiarea,Guiyang[J].GuizhouGeology, 1987, 4(4): 454-460. ]
7陈文一,杨瑞东. 贵州三叠纪遗迹化石及古地理意义[J]. 贵州地质,1990,7(4):382-386. [ChenWenyi,YangRuidong.ThesignificanceofTriassictracefossilsandpalaeographyinGuizhou[J].GuizhouGeology, 1990, 7(4): 382-386. ]
8时国. 贵阳花溪地区下三叠统大冶组遗迹化石及沉积环境研究[D]. 贵阳:贵州大学,2007. [ShiGuo.TracefossilsandtheirdepositionalenvironmentsoftheEarlyTriassicDayeFormationinHuaxiregion,Guiyang[D].Guiyang:GuizhouUniversity, 2007. ]
9时国,喻美艺,罗茂,等. 贵阳花溪地区下三叠统大冶组遗迹化石及沉积环境分析[J]. 沉积学报,2009,27(3):427-434. [ShiGuo,YuMeiyi,LuoMao,etal.TracefossilsandtheirdepositionalenvironmentsoftheEarlyTriassicDayeFormationinHuaxiarea,Guiyang[J].AtcaSedimentologicaSinica, 2009, 27(3): 427-434. ]
10BeattyTW,ZonneveldJP,HendersonCM.AnomalouslydiverseEarlyTriassicichnofossilassemblagesinnorthwestPangea:Acaseforashallow-marinehabitablezone[J].Geology, 2008, 36 (10): 771-774.
11FraiserML,BottjerDJ.OpportunisticbehaviourofinvertebratemarinetracemakersduringtheEarlyTriassicaftermathoftheend-Permianmassextinction[J].AustralianJournalofEarthSciences:AnInternationalGeoscienceJournaloftheGeologicalSocietyofAustralia, 2009, 56(6): 841-857.
12KnaustD.Theend-Permianmassextinctionanditsaftermathonanequatorialcarbonateplatform:insightsfromichnology[J].TerraNova, 2010, 22(3): 195-202.
13MacNaughtonRB,ZonneveldJP.Trace-fossilassemblagesintheLowerTriassicToadFormation,LaBicheRivermaparea,southeasternYukon[J].BulletinofCanadianPetroleumGeology, 2010, 58(2): 100-114.
14ZonneveldJP,GingrasMK,BeattyTW.DiverseichnofossilassemblagesfollowingtheP-Tmassextinction,LowerTriassic,AlbertaandBritishColumbia,Canada:evidenceforshallowmarinerefugiaonthenorthwesterncoastofPangaea[J].PALAIOS, 2010, 25(6): 368-392.
15ChenZhongqiang,TongJinnan,FraiserML.Tracefossilevidenceforrestorationofmarineecosystemsfollowingtheend-PermianmassextinctionintheLowerYangtzeregion,SouthChina[J].Palaeogeography,Palaeoclimatology,Palaeoecology, 2011, 299(3/4): 449-474.
16HofmannR,GoudemandN,WasmerM,etal.Newtracefossilevidenceforanearlyrecoverysignalintheaftermathoftheend-Permianmassextinction[J].Palaeogeography,Palaeoclimatology,Palaeoecology, 2011, 310(3/4): 216-226.
17ChenZhongqiang,BentonMJ.Thetimingandpatternofbioticrecoveryfollowingtheend-Permianmassextinction[J].NatureGeoscience, 2012, 5(6): 375-383.
18陈晶,童金南,牛志军,等. 贵州青岩地区早—中三叠世生物地层[J]. 地球科学,2010,35(1):51-61. [ChenJing,TongJinnan,NiuZhijun,etal.Lower-MiddleTriassicstratainQingyan,Guizhouprovince,SouthChina[J].EarthScience, 2010, 35(1): 51-61. ]
19秦典夕,颜承锡,熊剑飞. 黔中三叠纪牙形类生物地层研究的新进展[J]. 贵州地质,1993,10(2):120-130. [QinDianxi,YanChengxi,XiongJianfei.NewadvancesinthebiostratigraphyofTriassicconodontsincentralGuizhou[J].GuizhouGeology, 1993, 10(2): 120-130. ]
20贵州省地矿局. 贵阳幅1:20万区域地质调查报告[R]. 贵阳:贵州省地质局,1976:163-168. [The108GeologicalTeamofGuizhou. 1: 200000regionalgeologicalsurveyofGuiyang[R].Guiyang:GuizhouGeologicalBureau, 1976: 163-168. ]
21贵州省地矿局.贵州省区域地质志[M]. 北京:地质出版社,1987:450-484. [GuizhouBureauofGeologyandMineralExpioration.RegionalGeologyofGuizhouProvince[M].Beijing:GeologicalPublishingHouse, 1987:450-484.]
22刘泽均,王文彬. 下扬子区下三叠统遗迹化石及其环境意义[J]. 地层学杂志,1990,14(3):203-208. [LiuZejun,WangWenbin.LowerTriassictracefossilsfromtheLowerYangziregionandtheirsignificanceofsedimentaryenvironments[J].JournalofStratigraphy, 1990, 14(3): 203-208. ]
23杨式溥. 遗迹化石的古环境和古地理意义[J]. 古地理学报,1999,1(1):7-19. [YangShipu.Palaeoenvironmentalandpalaeogeographicsignificanceoftracefossils[J].JournalofPalaeogeography, 1999, 1(1): 7-19. ]
24赵小明,童金南. 浙江煤山钻孔二叠—三叠系界线剖面遗迹化石的两幕式变化[J]. 中国科学(D辑):地球科学,2010,40(9):1241-1249. [ZhaoXiaoming,TongJinnan.TwoepisodicchangesoftracefossilsthroughthePermian-TriassictransitionintheMeishancores,Zhejiangprovince[J].ScienceChina(Seri.D):EarthSciences, 2010, 40(9): 1241-1249. ]
25金惠,张俊,马石玉. 四川盆地上三叠统遗迹化石及其沉积环境[J]. 岩性油气藏,2013,25(1):24-28. [JinHui,ZhangJun,MaShiyu.TracefossilsandsedimentaryenvironmentoftheUpperTriassicXujiaheFormationinSichuanBasin[J].LithologicReservoirs, 2013, 25(1): 24-28. ]
26周志澄,杨昊,李罡,等. 四川广安谢家槽早三叠世遗迹化石及其古生态意义[J]. 古生物学报,2014,53(1):52-69. [ZhouZhicheng,YangHao,LiGang,etal.EarlyTriassictracefossilsfromtheXiejiacaoSection,Guang’an,Sichuanandtheirpaleoecologicsignificances[J].ActaPalaeontologicaSinica, 2014, 53(1): 52-69. ]
27龚一鸣. 中国的遗迹化石研究[J]. 矿物岩石,2001,21(3):205-207. [GongYiming.StudyoftracefossilsinChina[J].JournalofMineralogyandPetrology, 2001, 21(3): 205-207. ]
28林世国,施振生,李君,等. 四川盆地上三叠统生物扰动环境分析及与储集性能的关系[J]. 天然气地球科学,2012,23(1):74-80. [LinShiguo,ShiZhensheng,LiJun,etal.EnvironmentalinterpretationofupperTriassicbioturbationstructuresandcorrelationwithpetrophysicalpropertiesofreservoirsinSichuanBasin[J].NaturalGasGeoscience, 2014, 23(1): 74-80. ]
29TaylorA,GoldringR,GowlandS.Analysisandapplicationofichnofabrics[J].Earth-ScienceReviews, 2003, 60(3/4): 227-259.
30SavrdaCE,BottjerDJ.Theexaerobiczone,anewoxygen-deficientmarinebiofacies[J].Nature, 1987, 327(6117): 54-56.
31张立军,龚一鸣,马会珍. 华南泥盆纪遗迹化石及遗迹相[J]. 古地理学报,2011,13(4):397-418. [ZhangLijun,GongYiming,MaHuizhen.TheDevoniantracefossilsandichnofaciesfromSouthChina[J].JournalofPalaeogeography, 2011, 13(4): 397-418. ]
32KnaustD.TheichnogenusRhizocorallium:classification,tracemakers,palaeoenvironmentsandevolution[J].Earth-ScienceReviews, 2013, 126: 1-47.
33李越,袁金良,林天瑞. 安徽淮南下寒武统馒头组的遗迹化石[J]. 古生物学报,1999,38(1):114-124. [LiYue,YuanJinliang,LinTianrui.LowerCambriantracefossilsfromtheMantouFormationofHuainan,Anhui[J].ActaPalaeontologicaSinica, 1999, 38(1): 114-124. ]
34时国,田景春,喻美艺. 贵阳花溪地区下、中三叠统层序地层及其海平面变化[J]. 地层学杂志,2011,35(4):397-403. [ShiGuo,TianJingchun,YuMeiyi.Anoutcropsequence-stratigraphicstudyoftheLowerandMiddleTriassicintheHuaxiarea,Guiyang[J].JournalofStratigraphy, 2011, 35(4): 397-403. ]
35费安玮,张忠涛. 陕西金粟山中奥陶统生物扰动构造与古环境分析[J]. 地球科学,2002,27(6):703-710. [FeiAnwei,ZhangZhongtao.BioturbationstructuresandpaleoenvironmentanalysisoftheMiddleOrdovicianatJinsushan,Shananxi[J].EarthScience, 2002, 27(6): 703-710. ]
36PietschC,BottjerDJ.TheimportanceofoxygenforthedisparaterecoverypatternsofthebenthicmacrofaunaintheEarlyTriassic[J].Earth-ScienceReviews, 2014, 137: 65-84.
J]. 地球科学,2015,40(2):381-396. [ZhangLijun,ZhaoZhao,GongYiming.TraceFossilsasaproxyofthebig5biotic-andenvironmentaleventsinthePhanerozoic[J].EarthScience, 2015, 40(2): 381-396. ]
8王约,王训练,史晓颖. 贵州独山地区晚泥盆世F-F生物灭绝后的先驱生物及其在生态系统重建过程中的意义[J]. 中国科学(D辑):地球科学,2006,36(4):305-315. [WangYue,WangXunlian,ShiXiaoying.PioneerorganismsafterF-FmassextinctioninDushanregion,Guizhouprovince,andtheirsignificanceinestablishingnewecosystem[J].ScienceChina(Seri.D):EarthSciences, 2006, 36(4): 305-315. ]
39PayneJL,LehrmannDJ,WeiJiayong,etal.Thepatternandtimingofbioticrecoveryfromtheend-PermianextinctiononthegreatbankofGuizhou,Guizhouprovince,China[J].Palalos, 2006, 21(1): 63-85.
TraceFossilsandTheirPaleoenvironmentalSignificanceoftheEarlyTriassicAnshunFormationinHuaxiArea,Guiyang
YUGuanMeiSHIGuo
(CollegeofEarthSciences,EastChinaUniversityofTechnology,Nanchang330013,China)
ThetracefossilsinGuiyangareawasagoodproxyforecosystemrecoveryaftertheend-Permianbioticcrisis.Atotalof8ichnogeneraand9ichnospecieswereidentiedintheAnshunFormation(includingfourmembers,A1,A2,A3,A4),theirverticaldistributionandevolutionindicatethat:tracefossilsweredominatedbysmall,simple,horizontalburrowsofPlanolites montanus, Palaeophycus curvatusandPhycodes palmatusinA1;Tracefossilsbecomemoreabundantanddiverse,andbioturbationindexincreasedinA2andA3.Inadditiontothe3ichnospeciesichnogeneranotedinA1, 6otherichnospecieswereabundantinthesetwomembers: Beaconichnus darwinum, Chondrites recurvus, Thalassinoides paradoxicus, Thalassinoides horizontalis, Arenicolitesisp., Rhizocorallium commune.ComplexfaunawithRhizocoralliumandThalassinoidesinA3representedasignificantimprovementinthemarineenvironmentinthelateEarlyTriassic.NotracefossilswerefoundinA4.ThesedimentaryenvironmentfromA1toA3occurinplatformmarginshoaltoplatformslopetotidaltosupratidalzone,respectively.DuringA1toA3,theoxygenconditionsofmarineenvironmenthadameliorated.Thisconditioncouldprovideanoptimalzoneforbenthiccolonization.ThecharacteristicsofthetracefossilsfromupperA1toA3indicatedthatthestudyareacameintothehabitablezoneenvironment.A4wasdepositedinthesemirestrictedplatform,andintheregionabovethehabitablezonethatthebiologicalactivityisrelativelyweak.Thestudyoftracefossilssuggestedmarinebenthicrecoverysystemwasrelativelyactiveinthecarbonateplatformmargin.
tracefossil;AnshunFormation;habitablezone;sedimentaryenvironment;ecosystembioticrecovery
A
1000-0550(2016)04-0626-08
10.14027/j.cnki.cjxb.2016.04.002
2015-08-28; 收修改稿日期: 2015-12-25
国家自然科学基金项目(41002007);江西省博士后科研基金项目(2014KY31)[Foundation: National Natural Science of China, No. 41002007; Post Doctoral Research Foundation of Jiangxi Province, No. 2014KY31]
余关美女1990年出生硕士研究生地质资源与地质工程E-mail:yuguanmei@163.com
时国男讲师E-mail:309362105@qq.com
Q911
