北京市灵山亚高山草甸植被群落特征
2016-07-26赵方莹唐晓芬史振华
赵方莹, 刘 飞, 程 婕, 唐晓芬, 史振华
(1.北京圣海林生态环境科技股份有限公司, 北京 100083; 2.北京市门头沟区水务局, 北京 102300)
北京市灵山亚高山草甸植被群落特征
赵方莹1, 刘 飞1, 程 婕2, 唐晓芬1, 史振华1
(1.北京圣海林生态环境科技股份有限公司, 北京 100083; 2.北京市门头沟区水务局, 北京 102300)
摘要:[目的] 计算分析北京市灵山亚高山草甸区内自然坡面和侵蚀沟道两种立地类型上的植物群落特征差异,为侵蚀沟道植被人工修复提供理论依据和技术支撑。[方法] 采用野外调查和统计分析相结合的方法,对不同立地类型上的植被群落特征进行了研究。[结果] (1) Shannon-Wiener指数、Margalef指数均表现为:侵蚀沟道<自然坡面,而Simpson指数、Pielou指数表现为:侵蚀沟道>自然坡面; (2) 侵蚀沟道与自然坡面群落相似性系数为0.884,β多样性指数为0.178,表明侵蚀沟植物群落组成与自然坡面的相似性较高; (3) 自然坡面和侵蚀沟道内硬质早熟禾(Poa sphondylodes Trin)、苔草(Carex dispalata )的重要值和生态位宽度均较大,为主要的优势种;多数物种之间存在生态位重叠,但重叠值较小。[结论] 侵蚀沟道处于演替初期,趋向于恢复到群落自然状态,硬质早熟禾和苔草为主要的先锋优势种。
关键词:多样性指数; 生态位宽度; 亚高山草甸
文献参数: 赵方莹, 刘飞, 程婕, 等.北京市灵山亚高山草甸植被群落特征[J].水土保持通报,2016,36(3):165-171.DOI:10.13961/j.cnki.stbctb.2016.03.030
亚高山草甸是以多年生中生草本植物为主的隐域性植被类型,在中国分布很广,具有重要的研究价值。目前,对亚高山草甸的生态学研究,往往是将亚高山草甸作为植被类型整体进行研究[1],对其进行分类和排序[2],分析群落多样性和生态位[3-5],而对损毁草甸的植被恢复研究较少。由于亚高山地区人为活动加剧造成植被退化,土壤侵蚀和水土流失现象严重,生态系统的恢复问题亟待解决。亚高山草甸地区侵蚀沟道内的植被恢复属于次生演替,次生演替趋向于恢复破坏前的原始群落类型。宋晓谕[6]在甘南亚高山草甸弃耕演替过程的研究指出,群落优势种呈一年生杂草向豆科、禾草转变的规律。纪磊等[7]对不同退化程度的高山草甸与亚高山草甸的演替过程进行研究,指出在放牧干扰下高山草甸和亚高山草甸的植被丰富度和优势种具有相异性,而盖度和高度随放牧强度的增大均降低。因此,运用生态学理论研究亚高山草甸,探讨亚高山草甸的生物多样性、群落结构及次生演替规律,对于实现亚高山草甸植被恢复与生态重建具有重要价值。
北京灵山亚高山草甸是典型的山地草甸之一,草层茂密,是北京郊区著名的生态旅游地。许多生态学者对灵山亚高山草甸进行了研究,多集中在植物多样性、植物区系划分及旅游等人为活动的影响方面[8-11]。马克平等[8]研究指出灵山物种丰富度和物种多样性指数随海拔升高而下降,物种均匀度则随海拔升高而增加。高贤明等[9]的研究发现旅游和放牧活动造成亚高山草甸物种多样性下降速度加快。向春玲等[10]对东灵山的高山草甸的研究指出物种丰富度和多样性指数随着海拔的增加而下降,并受到旅游和放牧的影响。目前,针对亚高山草甸区损毁草甸的植被恢复研究很少。灵山高山地区坡度大、生长季短,植被覆盖度降低后土壤易受侵蚀,引起水土流失及植物种资源的严重缺失。目前,灵山土壤侵蚀主要包括为沟蚀、面蚀和少量的风力侵蚀。由于灵山风景区旅游的开发和过度放牧等人类活动对环境的影响逐渐增大,超出草甸生态系统的承载力,造成亚高山草甸区的植被退化严重,坡面沟蚀和面蚀的发展速度极快[12-13]。因此,治理侵蚀沟道对促进损毁草甸植被恢复,保护物种多样性,维持良好的生态环境,实现亚高山草甸地区的可持续发展具有十分重要的意义[10,12]。
本文通过对灵山草甸的自然植被状况进行调查和研究,分析自然坡面及侵蚀沟道自然修复次生演替植被的差异,选配适宜的植物种进行植被恢复,为北京地区亚高山侵蚀沟修复及植被多样性保护提供理论依据和技术支撑。
1研究区概况
灵山位于北京市门头沟区,北纬39°48′—40°04′,东经115°24′—115°36′之间,主峰海拔2 303 m。灵山不仅是北京地区的最高峰,也是华北地区的主要高峰之一。灵山地区包括百花山、东灵山、小龙门、龙门涧等地,属于太行山系,小五台山脉的余脉。灵山地区为温带半湿润季风气候,夏季受东南季风影响,冬季受西伯利亚寒流的控制,表现为夏季高温多雨,冬季寒冷干燥。年平均降水量约638.8 mm,其中6—8月份降水量为400~500 mm,占全年降水量的60%~80%;冬季降水量仅占全年的20%,多西北风。年平均气温2~7 ℃,>0 ℃年积温2 300~3 600 ℃,年日照2 600 h,全年无霜期在160 d以下。土壤主要以山地棕壤、亚高山草甸土和褐土为主,一般呈微酸性反应。成土母质主要有:花岗岩、砂岩、砾岩、安山岩等。从灵山下部至上部,植被依次分布有:次生灌丛(多为荆条、绣线菊、山杏等)、落叶阔叶林、针叶林和亚高山草甸。由于人为活动的干扰,原生植被几乎没有,主要是次生植被。
研究区内的3条侵蚀沟位于阴坡,总体地势南高北低,比降为337‰。其中,1号侵蚀沟沟长148 m,平均宽度3 m,比降329‰;2号侵蚀沟沟长357 m,平均宽度6 m,比降297‰;3号侵蚀沟沟长390 m,平均宽度5 m,平均深度1 m,比降279‰。侵蚀沟溯源侵蚀及下切侵蚀比较严重。人畜扰动破坏及比降较大,是造成侵蚀严重的主要原因。目前,对侵蚀沟治理主要是通过减少人畜扰动破坏,提高沟头区域地表植被覆盖,增强雨水入渗,减少溯源侵蚀;铺设自然石和沟壁铺设、放坡处理等措施,控制侵蚀沟中段下切速度和横向发育;侵蚀沟下部主要布设消能和分散措施,阻止侵蚀沟继续向下发展。此外,通过人工促进与自然恢复相结合的方式,实现侵蚀沟内的植被恢复。
2研究方法
2.1调查方法
2014年8月,在北京市灵山海拔亚高山草甸区(1 700~2 300 m)选择3条侵蚀沟及其周边进行样地调查,由于试验区带状分布,沿侵蚀沟沟头至沟尾的方向,每隔10 m布设1个样方,在侵蚀沟道沟底及沟两侧随机布设,同一水平位置处的自然坡面随机布设。采用1 m×1 m调查草本样方,并记录种数、株高、个数(丛数)、分盖度、总盖度,共调查了30个样方。
2.2数据处理
(1) 植物群落α多样性分析方法。重要值是研究某个种在群落中的地位和作用的综合数量指标。是相对盖度、相对频度的总和,其公式为:

(1)

Shannon-Wiener(H) 多样性指数:
(2)
(3)
式中:Ni——种i的个体数;N——该区全部物种个体数。
Pielou(J)均匀度指数:
J=(-∑pilnpi)/lnS
(4)
式中:S——物种数。
Simpson(λ)生态优势度指数:
(5)
Margalef(D)丰富度指数:
D=(S-1)/lnN
(6)
(2) 植物群落β多样性分析方法。群落相似性系数:
CSI=2C/(A+B)
(7)
式中:CSI——群落相似性系数;C——两群落共有的物种数;A,B——两群落各自的物种数。当2个群落物种组成完全相同时,相似性系数为1;当2个群落物种组成完全不同时,相似性系数为0。
β多样性指数:
β=(lnR0-lnRn)/ln2
(8)
式中:R0——2个最大相似性群落的相似系数;Rn——环境梯度上最不相似的2个群落的相似系数;β——多样性指数。
(3) 生态位测度方法。生态位宽度是物种对环境资源利用多样性的一种测度,反映不同物种的生态适应幅度。
Levins生态位宽度:
(9)
式中:Pij——种i对第j个资源的利用占其对全部资源利用的频度,即Pij=nij/Ni,而nij为种i在资源j上的优势度;Ni——种i所在利用全部资源位上的重要值之和;r——资源等级数;B(L)i具有域值[1/r,1]。
生态位重叠是指一定资源序列上,2个物种竞争同等资源而相互重叠的情况,为物种在其余生态因子联系上的相似性,反映不同物种对生态条件要求的相似程度,较常用的计算方法如下:
(10)
式中:Lih——物种i重叠物种h的生态位重叠指数;Phj——物种h对第j个资源利用占其对全部资源利用的频度;B(L)——生态位宽度指数。
3结果与分析
3.1物种组成
本次调查中共出现24种植物,分布于18科(表1)。其中自然坡面上有22种植物,包括菊科有4种植物,蔷薇科3种植物。其余紫草科、豆科、禾本科、车前科、报春花科等各有1种植物。侵蚀沟道中有19种植物,菊科4种,蔷薇科和紫草科各2种,其他科植物各1种。从组成植物群落生活型来看,以多年生草本为主,占总种数的91.67%,并有一年生草本伴生。
3.2植被盖度及密度分析
如表2所示,与自然坡面群落相比,侵蚀沟道植物群落内的物种数较少,植物平均高度也比较低,其平均范围在2.39~12.38 cm。侵蚀沟道内植被平均盖度(27.39%)低于自然坡面(65.40%),差异性显著(p=0.001<0.05),而植被密度相差不大,不存在显著差异(p=0.72>0.05),说明侵蚀沟道内的植被生长状况较差。这是由于侵蚀沟道内植物生长所需的土壤和水文条件较差,导致植株矮小,覆盖度低。
3.3物种重要值分析
灵山亚高山草甸上自然坡面及侵蚀沟内主要植物种的重要值如表3所示。自然坡面上的主要物种包括白毛委陵菜、硬质早熟禾、苔草、车前,其重要值大小分别为39.42,26.22,23.75,20.49。而侵蚀沟道内植物群落的优势种和自然坡面的相差不大,其主要的物种包括硬质早熟禾、小红菊、中华风毛菊、车前、苔草,其重要值分别40.66,17.34,17.28,16.30,16.24。可以看出在自然坡面及侵蚀沟道植物群落内,硬质早熟禾、车前和苔草作为优势种或亚优势种存在,表现为很强的生长优势。

表1 主要的植物种组成
注:O表示物种存在; ×表示物种不存在。

表2 物种盖度及密度情况

表3 主要植物的重要值 %
3.4物种多样性分析
物种多样性作为植物群落的重要特征,它不仅受物种数量的影响,也受物种空间分布的均匀性的影响。因此,物种多样性不仅标示植物群落的物种组成状况,也可以衡量群落的物种空间分布结构。在多样性指数中,Shannon-Wiener指数为变化度指数,是能够较好地反映个体密度、生境差异、群落类型、演替阶段的指数,它是物种丰富度和均匀度的函数,物种数量越多,其分布越均匀,值也越大[14-16]。Margalef指数指一个群落或环境中物种数目的多寡。Simpson指数是反映群落优势度较好的指标,该值越大优势种的地位越突出,群落多样性越低。Pielou指数值物种个体的相对丰富度或所占的比例,指数越高,群落内各物种的个体数越接近。
由表4可知,自然坡面和侵蚀沟道中Shannon-Wiener指数分别为1.456,1.367,说明了侵蚀沟道中的物种多样性小于自然边坡。这与侵蚀沟道内的土壤和水文条件较差,不利于植物的生长繁殖有关,并且侵蚀沟内植物自然恢复时间较短,植物数量和种类均较少,造成其物种多样性较低。此外,与Shannon-Wiener指数变化特征相似,侵蚀沟道的Margalef指数(1.383)小于自然坡面的丰富度(1.590);而Simpson指数表现为侵蚀沟道(0.561)>自然边坡(0.477);均匀度也表现为侵蚀沟道(0.763)>自然边坡(0.710)。由于侵蚀沟道内样地内优势种相近分布较多;而其他物种的分布较少数量相差不大有关。

表4 不同立地类型的生态群落多样性
β多样性指数可以用来测度群落的物种多样性沿着环境梯度变化的速率或群落间的多样性[17]。群落间的共同种越少,β多样性越高。侵蚀沟道植物群落和自然坡面植物群落相比,群落相似性系数为0.884,β多样性指数为0.178,侵蚀沟道内的植物群落组成与自然坡面的相似性较高,共有物种较多,说明侵蚀沟内植物恢复符合自然演替规律。
3.5植物群落的生态位分析
生态位宽度可以用来衡量植物种对环境资源的利用情况,物种的生态位宽度越大,在群落环境中的适应性和对资源的利用能力较强,占据较宽的生态幅,使得物种竞争力较强。反之,物种生态位宽度较小,则其适应性、竞争力较薄弱[18-19]。生态位重叠反映了种群间对资源利用的相似程度和竞争关系,生态位重叠较大的物种间具有相近的生态特性或对环境因子有互补性的要求[14]。
表5为自然边坡和侵蚀沟道内不同植物种的生态位宽度,自然边坡中车前的生态位宽度最大,其值为0.504。其次,苔草、钝萼附地菜、藁本的生态位宽度也较大,分别为0.495,0.487,0.458。在侵蚀沟道内,车前和小红菊的重要值较大,且分布均匀,因而其生态位宽度较大,分别为0.669,0.685。此外,苔草、地榆、硬质早熟禾的生态位宽度也较大,分别为0.499,0.479,0.478。虽然白毛委陵菜的重要值较大,但是由于其分布比较集中,因而其生态位宽度较窄。车前和苔草作为重要的植物种,其生态位宽度较宽,说明这2个物种对环境的适应能力和对资源的利用能力较大,具有较强的竞争力。
由表6可知,自然边坡内植物群落中,具有生态位重叠的种对数共有276对,占总对数的59.74%,没有生态位重叠的种对数共有186对,占总对数的40.26%。在群落中,生态位重叠值在0~0.134之间,车前、苔草、钝萼附地菜、藁本的生态位较宽且分布均匀,与大多数物种间都具有生态位重叠。其中钝萼附地菜与萹蓄、岩青蓝与萹蓄、白毛委陵菜与蓝花棘豆的生态位重叠值,分别为0.134,0.121,0.108,生态位重叠值较小。这说明大多数种群之间不存在强烈竞争,关系较为协调、平衡,对资源的分享利用比较充分[4]。
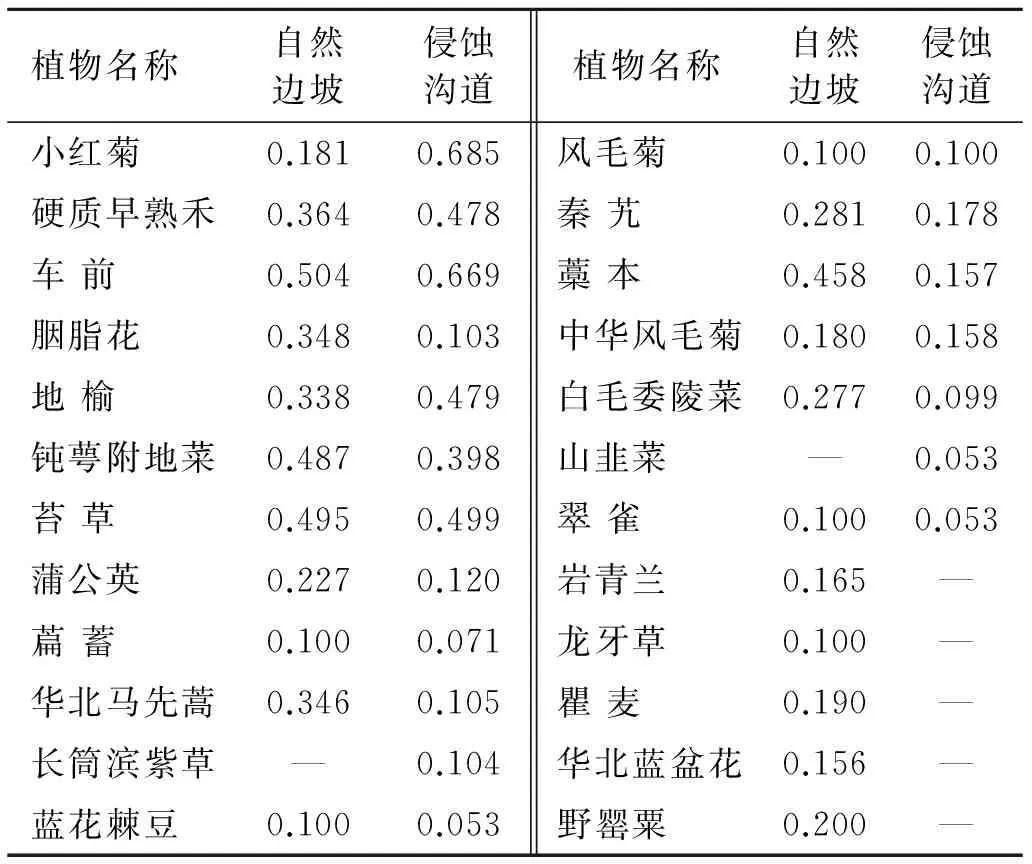
表5 不同立地类型内植物种的生态位宽度
侵蚀沟道内植物群落的生态位重叠情况如表7所示,具有生态位重叠的种对数共有244对,占总对数的71.35%。没有生态位重叠种对数共有98对,占总对数的28.65%。在侵蚀沟道植物群落中,各物种的生态位重叠值均较小,其中小红菊与蒲公英、苔草与白毛委陵菜的生态位重叠相对较大,也仅为0.070,0.068。这说明侵蚀沟道恢复的植物之间不存在激烈竞争,对资源的利用差异性较大。

表6 自然坡面植物群落生态位重叠

表7 侵蚀沟道植物群落生态位重叠
4讨论与结论
4.1讨 论
由于侵蚀沟道处于演替初期,一些传播快、数量多的物种迅速萌发,占据空间优势,群落结构简单,物种多样性Shannon-Wiener指数和Margalef丰富度较低,而自然边坡物种多样性较高是植物物种多年演替的结果。侵蚀沟道内的物种Simpson优势度指数高于自然坡面,这是次生演替初期有少数几个优势种,而自然状态下的优势种较多且分布局均匀。这与杜国祯等[20]关于亚高山草甸废弃地的研究结果相一致。侵蚀沟道植物群落和自然坡面植物群落β多样性指数较低,群落相似性较高,说明侵蚀沟内植被恢复过程中向着恢复群落原生状态演替,符合自然演替规律。
在自然边坡中,白毛委陵菜、硬质早熟禾和苔草的重要值较大,但白毛委陵菜的生态位宽度较小,而苔草和硬质早熟禾的生态位较宽,苔草和硬质早熟禾为主要的优势种。侵蚀沟道中小红菊、硬质早熟禾、车前的生态位宽度较大,但是硬质早熟禾的重要值较大占绝对优势,为群落优势种,小红菊和车前为主要的伴生种。群落优势种与生态位宽度之间不存在必然联系,这与李斌等[4]在云顶山的研究相一致。侵蚀沟道中的种群生态位重叠值均较低,说明演替初期物种之间不存在激烈竞争,这与环境对物种的自然选择有关,易于传播和萌发的物种首先占据沟道生长,对资源的利用不存在激烈竞争。
4.2结 论
(1) 北京灵山亚高山草甸上硬质早熟禾、小红菊、苔草、车前等重要值和生态位宽度较大,为主要优势种,其对环境的适应力和竞争力较大。大多数物种间均有生态位重叠,但是重叠程度较低,因此物种之间的相互影响也较小。
(2) 与自然坡面植物群落相比,侵蚀沟道内的植物的盖度、高度均较小,植物生长情况较差。此外侵蚀沟道内物种多样性和Margalef指数均低于自然坡面,而Simpson优势度指数和Pielou均匀度指数均高于自然坡面,这与侵蚀沟道内植被的自然修复时间较短、土壤及水文环境较差有关,说明侵蚀沟道正处于演替初期。
(3) 侵蚀沟植物群落组成与自然坡面的相似性较高,说明侵蚀沟道植被恢复趋向于恢复到破坏前的自然状态。
综上所述,侵蚀沟道内自然恢复的植物群落和自然坡面群落的相似性较大,但是在多样性、优势度方面均小于自然群落,与自然坡面存在一定的差距。这与侵蚀沟道处于演替初期有关。根据人工植被恢复优先选择乡土物种的原则,合理选配小红菊、苔草、硬质早熟禾等优势种作为先锋物种,并结合一定的灌木配置,促进植被的自然演替进程,建立与环境相适应的、持续稳定的植物群落,从而有效的控制灵山坡面侵蚀和沟道侵蚀,并防治水土流失。
[参考文献]
[1]王琳,张金屯,欧阳华.历山山地草甸的生态关系[J].山地学报,2004,22(6):669-674.
[2]江洪.东灵山植物群落生活型谱的比较研究[J].植物学报,1994,36(11):884-894.
[3]李军玲,张金屯,郭逍宇.关帝山亚高山灌丛草甸群落优势种群的生态位研究[J].西北植物学报,2003,23(12):2081-2088.
[4]李斌,李素清,张金屯.云顶山亚高山草甸优势种群生态位研究[J].草业学报,2010,19(1):6-13.
[5]宋爱云,刘世荣,史作民,等.卧龙自然保护区亚高山草甸的数量分类与排序[J].应用生态学报,2006,17(7):1174-1178.
[6]宋晓谕.甘南亚高山草甸弃耕演替过程中植物群落特征变化模式研究[D].兰州:兰州大学,2010.
[7]纪磊,干友民,罗元佳,等.川西北不同退化程度高山草甸和亚高山草甸的植被特征[J].草业科学,2011,28(6):1101-1105
[8]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究(Ⅱ):丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[9]高贤明,马克平,陈灵芝,等.旅游对北京东灵山亚高山草甸物种多样性影响的初步研究[J].生物多样性,2002,10(2):189-195.
[10]向春玲,张金屯.东灵山亚高山草甸物种多样性变化及其影响因子[J].北京师范大学学报:自然科学版,2009,45(3):275-278.
[11]刘晓.北京东灵山亚高山草甸维管植物区系与多样性研究[D].北京:北京林业大学,2011.
[12]李薇,潘祺志,唐海萍.北京东灵山亚高山草甸旅游开发与保护研究[J].资源开发与市场,2007,23(1):81-83.
[13]高俊峰.北京东灵山地区人类活动对植物多样性分布的影响研究[D].北京:北京林业大学,2007.
[14]徐广平,张德罡,徐长林,等.东祁连山高寒草地不同生境类型植物群落α及β多样性的初步研究[J].草业科学,2006,23(6):1-5.
[15]Tilinan D. Cause consequences and ethics of biodiversity [J]. Nature,2000(405):208-211.
[16]邱波,任青吉,罗燕江,等.高寒草甸不同生境类型植物群落的α及β多样性研究[J].西北植物学报,2004,24(4):655-661.
[17]王世昌.云顶山亚高山草甸物种多样性研究[J].山西林业科技,2011,40(3):17-19,23.
[18]白永飞,邢雪荣,许志信,等.内蒙古高原针茅草原群落β多样性研究[J].应用生态学报,2000,11(3):408-412.
[19]李明,蒋德明,押田敏雄,等.科尔沁沙地人工固沙群落草本植物种群生态位特征[J].草业科学,2009,26(8):10-16.
[20]杜国祯,王刚.亚高山草甸弃耕地演替群落的种多样性及种间相关分析[J]. 草业科学,1991,8(4):53-57.
收稿日期:2015-06-01修回日期:2015-07-09
文献标识码:A
文章编号:1000-288X(2016)03-0165-07
中图分类号:S812
Vegetation Characteristics of Subalpine Meadow Community in Lingshan Mountain of Beijing City
ZHAO Fangying1, LIU Fei1, CHENG Jie2, TANG Xiaofen1, SHI Zhenhua1
(1.BeijingShenghailinEcologicalEnvironmentTechnologiesCo.,Ltd,Beijing100083,China; 2.MentougouWaterAuthorityofBeijingCity,Beijing102300,China)
Abstract:[Objective] The characteristics of plant communities in the natural slope and erosion gully of the subalpine meadow in Lingshan Mountain in Beijing City were investigated and analyzed to provide theoretical basis and technical support for vegetation restoration in erosion gullies. [Methods] The characteristics of plant communities at different sites were studied using the methods of standard ecological investigation methods and statistical analysis. [Results] (1) The Shannon-Wiener index and Margalef index were all as follows: erosion gully
Keywords:diversity index; niche breadth; subalpine meadow
第一作者:赵方莹(1974—),男(汉族),江苏省扬州市人,博士后,高级工程师,主要研究方向:边坡绿化与生态修复。 E-mail:shenghailin2015@163.com。
