海南霸王岭毛棉杜鹃根部真菌的多样性
2016-07-25廖映辉黄彩微史佑海丁琼
廖映辉++黄彩微++史佑海+丁琼
摘要:以生长于海南岛霸王岭的毛棉杜鹃(Rhododendron moulmainense)为材料,用分子检测技术对其根部真菌的rDNA-ITS序列进行扩增、克隆、测序,以及序列分析,共检测出28种真菌,其中子囊菌门(Ascomycota)19种,担子菌门(Basidiomycota)8种,接合菌门(Zygomycota)1种,其中物种最丰富的真菌类群为子囊菌门锤舌菌纲的柔膜菌目。采用Bootstrap法估算真菌的物种丰富度,表明霸王岭地区毛棉杜鹃的根系真菌至少有37种,并且物种累积曲线显示:随着根系真菌的宿主植物个体数增加,Shannon-weiner多样性指数和物种丰富度增大,且物种数增加的潜力仍然较大。研究结果表明,毛棉杜鹃根部真菌种类极丰富,有必要全面开展杜鹃花属植物根部真菌分子检测,并从中筛选出对杜鹃花属植物生长有良好效应的真菌。全面揭示杜鹃花属植物的根部共生真菌多样性必将有助于杜鹃花属植物的保育与引种栽培。
关键词:杜鹃花属;根系真菌;菌根;多样性
中图分类号: S685.210.2文献标志码: A文章编号:1002-1302(2016)06-0432-06
收稿日期:2016-01-26
基金项目:海南省重大科技项目(编号:ZDZX2013012)。
作者简介:廖映辉(1992—),女,福建龙岩人,硕士研究生,主要从事杜鹃花菌根真菌的多样性研究。E-mail:328949021@qq.com。
通信作者:丁琼,博士,讲师,主要从事菌根研究。E-mail:dingqiong1@gmail.com。杜鹃花科(Ericaceae)植物根系在自然生境中与真菌形成的互利共生体,称之为杜鹃花菌根(Ericoid mycorrhiza,ERM)[1]。野外条件下,杜鹃花类植物的菌根真菌能有效地加强其对土壤中可溶性无机态N、P或不可溶性N、P复合物的吸收利用[1-2]。北美地区的园艺生产中,常用杜鹃花类菌根真菌接种蓝莓,以减少化肥的施用[3]。杜鹃花属(Rhododendron)是杜鹃花科的物种较为丰富的属,古诗有云“花中此物为西施,芙蓉芍药皆嫫母”,具有极高的观赏价值和园林应用价值,在市场上具有极大的产业发展前景。我国是杜鹃花属植物最为重要的起源地和分布中心,共有约530种,占世界总数的59%[4]。相对于其他杜鹃花科植物欧石楠属(Calluna)和越橘属(Vaccinium)而言,杜鹃花属植物根系真菌的研究少之又少,自2000年来主要涉及宿主如R. fortunei、R. dauricum、R. decorum、R. obtusum var. kaempferi、R. argyrophyllum、R. floribundum、R. lochiae、R. pulchrum、R. maximum、R. rubropilosum等,且能形成杜鹃花类菌根真菌的大多数为子囊菌门,包括柔膜菌目真菌及树粉孢属真菌,少部分为担子菌门或其他类真菌[5-23]。
海南岛是我国唯一地处热带的岛屿,丰富的光照和温度资源、复杂的地形和地貌孕育了多样性的热带特色野生花卉资源,共分布9种1变种杜鹃花[24]。毛棉杜鹃(R. moulmainense Hook.f.)亦是其中一种,其分布范围还遍及我国江西、福建、湖南、广东、广西、四川、贵州和云南等地,多数生长在海拔700~1 500 m的山顶或山地林中。毛棉杜鹃花冠通常为淡紫色、粉红色或者淡红白色,狭漏斗形。花期长,花朵繁盛,颜色艳丽,还是杜鹃属植物中少有的耐热性种质资源,具有非常高的园林应用潜力。当前野生杜鹃引种、栽培十分困难是其园林应用的瓶颈。采用真菌分子检测技术,揭示毛棉杜鹃在自然环境中的根系真菌多样性,有助于今后筛选出能促进毛棉杜鹃生长的菌根真菌,可以有效地指导其引种、栽培及园林应用。本研究结果不仅对毛棉杜鹃种群的保育具有重要意义,而且对我国热带地方特色花卉资源利用及花卉产业的资源优势发挥有重要意义。
1材料与方法
1.1样品采集地点
样品采集于海南霸王岭国家级自然保护区(18.87°~1920° N,108.88°~109.33° E),地处海南岛西南部昌江县境内。保护区的气候属热带季风气候,年平均温度23.6 ℃,年均降水量1 500~2 000 mm[25]。海拔高度在100~1 700 m范围,保护区内的土壤以砖红壤为主要类型,随着海拔高度的增加不断过渡为山地红壤[26]。低地雨林和山地雨林是该地区分布最广的2个主要植被类型,低地雨林是以青梅(Vatica mangachapoi)、荔枝(Litchi chinensis)等为优势种,山地雨林以鸡毛松(Podocarpus imbricatus)、陆均松(Dacrydium pierrei)等为优势种[27]。
1.2样品采集
采集时,选取成年的植株,沿主干找到最细的发根部分,在不同位置上用枝剪小心剪下。将取到的发根同附着的土块在水中浸泡约1 h,用水小心冲洗干净后置于装有75%乙醇的塑料管中,标号,-80 ℃冰箱中保存。共采集8株毛棉杜鹃植株。
1.3真菌分子检测
1.3.1DNA提取杜鹃花属植物根系真菌DNA的提取采用改良的CTAB法[28]。在提取DNA之前,对根系样品进行预处理。每份样品提取12份DNA,每份DNA样品为随机挑选的50份约1.5 cm长的毛根,并置于1.5 mL的离心管中,加入20 μL CTAB提取缓冲液,放置于-80 ℃冰箱冰冻1夜后在冰浴中使用电钻研磨成沫。
1.3.2ITS扩增采用普通的PCR方法扩增真菌的内转录间隔区,扩增引物为ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS4 (5′-TCCTCCGCTTATTGATATGC-3′),引物序列由上海生物工程有限公司合成。PCR反应体系总共25 μL,分别含有:1 μL 模板DNA,0.2 mmol/L dNTPs,2.5 μL 10×PCR缓冲液(TaKaRa,Otsu,Japan),ITS1F与ITS4引物各0.2 μmol/L,0.8 μmol/L牛血清白蛋白(Bovine serum albumin,BSA),1.5 U Taq DNA聚合酶(TaKaRa,Otsu,Japan)。PCR反应程序:95 ℃预变性5 min;94 ℃变性40 s,55 ℃退火40 s,72 ℃延伸55 s,共30个循环;最后72 ℃延伸10 min。最后将ITS-PCR产物在0.8%的琼脂糖凝胶上电泳,用凝胶成像仪观察、拍照。
1.3.3PCR产物克隆及测序将同一植株的12份PCR产物混合后电泳、切胶回收。将回收的PCR产物作为外源DNA,克隆到pMD19-T Vector (TaKaRa,Otsu,Japan),在16 ℃ 下连接过夜,用E. coli DH5α感受态细胞(TaKaRa,Otsu,Japan)进行转化。从每株宿主植物的根系真菌克隆文库中随机挑选30个呈白斑的克隆,用引物ITS1F和ITS4扩增;PCR产物在0.8%的琼脂糖凝胶上电泳检测。电泳检测呈阳性PCR产物测序委托北京三博远志生物技术有限责任公司。
1.3.4生物信息学分析在Bioedit软件中对真菌的 ITS1-5.8S-ITS2 全序列进行编辑和比对。ITS序列一致性(identity)≥97%的真菌被视为同一个分类单元(operational taxonomical unit,OTU)。每个OTU的参考序列与NCBI(GenBank)数据库中的序列进行比对,确定与试验克隆亲缘关系最近的种属。将ITS序列通过ClustalX2进行序列间的匹配排序(必要时需要手动调整)。NJ系统发育树的构建采用MEGA 5。将本试验获得的ITS序列提交至NCBI数据库中,获得相应的序列号为KU550104-KU550131。
1.4数据分析
用软件Estimate S 9中提供的Bootstrap方法估计研究区物种丰富度,并计算Shannon-Weiner多样性指数,并用物种累积曲线评估抽样到的真菌群落的物种是否能较全面地反映当地物种库。
2结果与分析
2.1毛棉杜鹃根系真菌多样性
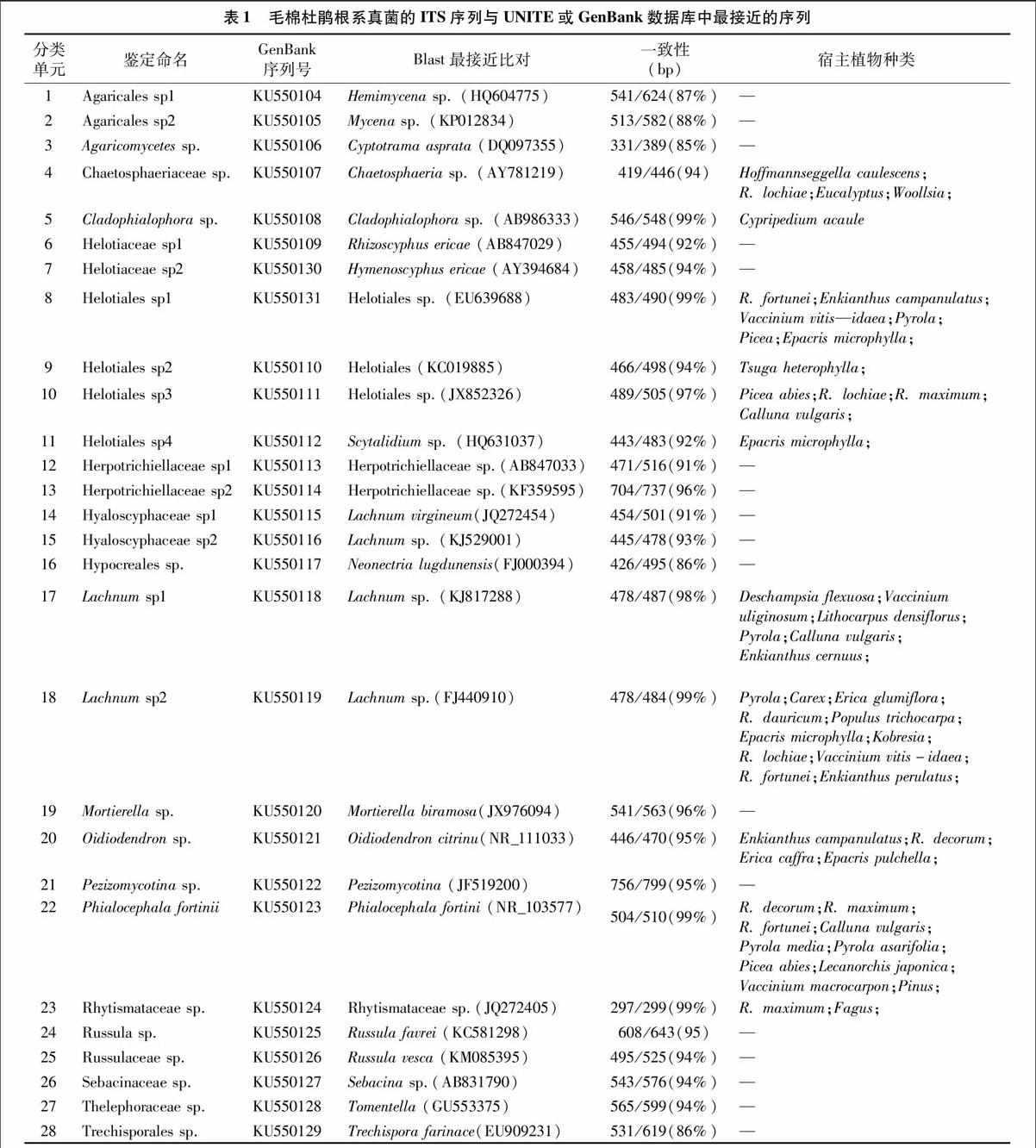
对8株毛棉杜鹃的根系真菌进行检测,获得真菌ITS有效序列84条,并划分为28个OTUs(以后统称为物种)(表1)。子囊菌门(Ascomycota)19种,其中包括锤舌菌纲(Leotiomycetes)13种、粪壳菌纲(Sordariomycetes)2种、散囊菌纲(Eurotiomycetes)3种和未能鉴定到纲水平的盘菌亚门(Pezizomycotina)1种;担子菌门(Basidiomycota)8种,均为伞菌纲(Agaricomycetes)的物种;除此以外,还有1种为接合菌门(Zygomycota)的物种。与此同时,共有26种真菌鉴定到了目的水平,分别为子囊菌门的柔膜菌目(Helotiales)、刺盾炱目(Chaetothyriales)、Chaetosphaeriales、肉座菌目(Hypocreales)、斑痣盘菌目(Rhytismatales)和树粉孢属(Oidiodendron);担子菌门的伞菌目(Agaricales)、红菇目(Russulales)、蜡壳耳目(Sebacinales)、革菌目(Thelephorales)、Trechisporales及接合菌门的被孢霉目(Mortierellales)(图1)。在目水平上,物种丰富度最高的真菌类群为锤舌菌纲的柔膜菌目(11种)(图2)。
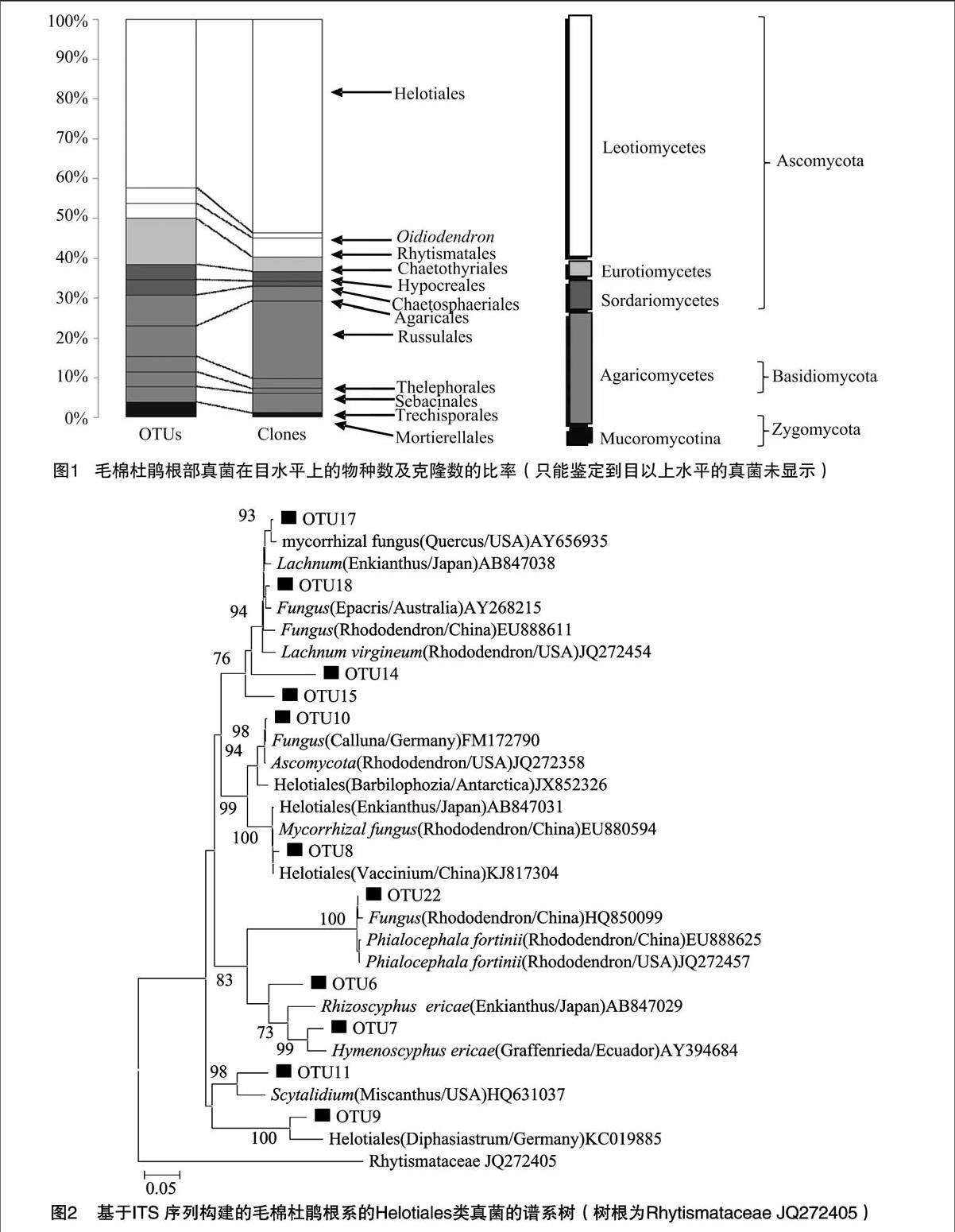
通过与NCBI数据库中的序列比对发现,有11个OTUs与来自草本或木本植物的根系真菌同源性达97%以上(表1)。其中7个OTUs(OTU 8-11;OTU 17-18;OTU 22)属于柔膜菌目,OTU 4与Chaetosphaeria sp.(AY781219)具有较高的相似性,OTU 5和OTU 23分别与Cladophialophora sp.(AB986333)、Rhytismataceae sp. (JQ272405)同源性高达99%,而OTU 20与Oidiodendron citrinum(NR_111033)同源性达95%,因此被鉴定为树粉孢属真菌。在这11个OTU中,9个OTUs都曾在杜鹃花科植物根系中检测到,1个OTU(OTU 5)在兰科植物Cypripedium的根系中出现,另一个(OTU 9)则在铁杉属根系中检测到。在这9个OTU中,除OTU 20和OTU 11仅在杜鹃花科植物根系中出现外,其余OTU(OTU 4、OTU 8、OTU 10、OTU 17、OTU 18、OTU 22、OTU 23)不仅在杜鹃花科植物根系(Rhododendron、Eucalyptus、Woollsia、Enkianthus、Vaccinium、Pyrola、Epacris、Calluna)中检测到,还在少数的兰科植物(Hoffmannseggella;Lecanorchis)和外生菌根根段(Picea;Deschampsia;Lithocarpus;Pinus;Fagus)中检测到。
2.3根系真菌物种丰富度估计及物种累积曲线
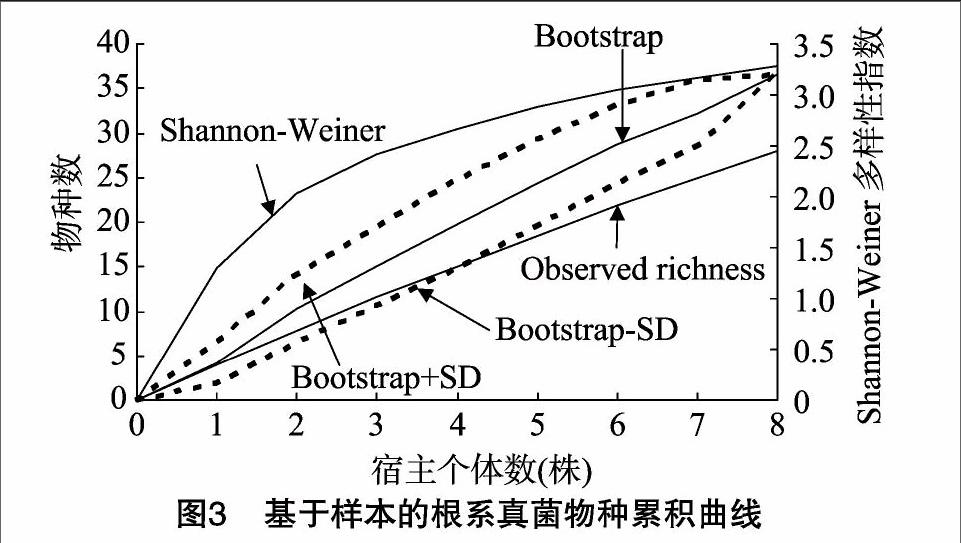
用Bootstrap算法估计的海南霸王岭毛棉杜鹃根系真菌物种数37种,高于实际观察到的物种数(28种)。随着根系真菌的宿主植物个体数增加,物种丰富度不断增大,且从物种累积曲线来看检测到的真菌物种数增加的潜力仍然较大,然而Shannon-Weiner多样性指数却趋于平缓(图3)。
3讨论与结论
本次从海南霸王岭毛棉杜鹃根系中共检测到28种真菌,通过序列比对和真菌ITS序列谱系树分析发现子囊菌门的真菌种类丰富约占68%(19种),其中又以柔膜菌目真菌占的比例最大,共有11种,其中有7种与来自杜鹃花科植物或少数其他草本或木本植物根系真菌同源性达97%以上。子囊
菌门柔膜菌目的真菌是一类多样且典型的杜鹃花菌根真菌,国内外大多学者在对杜鹃花菌根真菌研究中也都发现了该类真菌,在多种杜鹃花根系中都检测到[5,9-13,18,22,29]。在这7种柔膜菌目真菌中,OTU 9仅与来自Tsuga heterophylla的根系真菌(FJ152534)同源性达98%,OTU 11仅与Epacris microphylla的根系真菌同源性达98%[30],其余5个OTU都曾在多种杜鹃花科植物根系及少数外生菌根根段中检测到。OTU 7曾被报道与中国的Rhododendron fortunei和Vaccinium vitis-idaea(KJ817304),日本的Enkianthus campanulatus,美国的Pyrola和澳大利亚的Epacris microphylla等多种杜鹃花科植物根系共生[13,30-32],且曾在美国的Picea根系中出现[33],OTU10则在澳大利亚的Rhododendron lochiae,美国的Rhododendron maximum,德国的Calluna vulgaris(FM172790)和瑞士的Picea abies均有报道[5,9,34]。OTU 17和OTU 18都为粒毛盘菌(Lachnum)的物种,都与来自多种不同草本或木本植物的根系真菌同源性达97%以上。OTU 17曾在杜鹃花科植物Vacciniumuliginosu(KJ817288)、Pyrola、Calluna vulgaris和Enkianthus cernuus,禾本科植物Deschampsia flexuosa及壳斗科植物
Lithocarpus densiflorus的根系中出现[31-32,35-36]。OTU 18则曾在多种杜鹃花属植物Rhododendron lochiae、Rhododendron fortunei、Rhododendron dauricum(KJ817276)和其他多种杜鹃花科植物Pyrola、Epacris microphylla、Erica glumiflora、Enkianthus perulatus、Vaccinium vitis-idaea(AJ430215)中检测到[5,13,30-31,37]。OTU 22与Phialocephala fortinii(NR_103577)同源性高达99%,被鉴定为Phialocephala fortinii。Phialocephala fortinii是北半球地区侵染植物根系的典型的暗色有隔内生真菌(dark septate endophytes,DSE)复合体中常见的一种真菌。关于这种内生真菌或菌根真菌的习性在以往的研究中争议较大,有学者认为Phialocephala fortinii是菌根性的真菌[38-39],它曾在Rhododendron decorum、Rhododendron maximum、Rhododendron fortunei、Calluna vulgaris、Pyrola media、Pyrola asarifolia、Vaccinium macrocarpon出现[9-10,13,40-42]。而周诗涵等在台湾地区的红毛杜鹃根系中也检测到Phialocephala,菌根形态鉴定与rDNA序列分析,以及回接试验均证实Phialocephala能与杜鹃花类植物形成杜鹃花菌根[23]。
除此之外,还有4个OTU(OTU4、OTU5、OTU20、OTU23)
与NCBI中不同植物的根系真菌同源性达97%以上。OTU5与美国的Cypripedium acaule的根系真菌同源性达98%[43]。OTU4和OTU23不仅在杜鹃花科植物中检测到,还在其他植物根系中也有出现。前者曾报道在Hoffmannseggella caulescens、Rhododendron lochiae、Eucalyptus、Woollsia pungens根系出现[5,44-46],后者则在Rhododendron maximum和Fagus(FN619991)根系中检测到[9]。OTU20与Oidiodendron citrinum(NR_111033)同源性达95%,因此鉴定为树粉孢属真菌。树粉孢属是常见的杜鹃花菌根真菌,特别是在北半球的杜鹃花根系中是非常常见的,本研究中树粉孢属与日本的Enkianthus campanulatus(99%)、中国四川及云南的Rhododendron decorum(98%)、南非的Erica caffra(98%)及澳大利亚热带森林的Epacris pulchella的根系共生真菌同源性高达98%或99%[10,32,37,47]。然而,在本研究中并没有出现分布最广且在杜鹃花类菌根真菌中研究最多的Oidiodendron maius。从这些同源性达97%的根系真菌的宿主植物分析中推测杜鹃花类菌根真菌类型在目的水平上总体相似,但不同的种之间可能受到其寄主植物的影响。
担子菌门的真菌物种数在本研究中仅占29%,为伞菌纲中伞菌目、红菇目、蜡壳耳目、革菌目、Trechisporales。在早期,蜡壳耳目的真菌被认为是腐生真菌,后续的研究表明Sebacinales 真菌可以划分为Clad A与Clad B两大支系[48]。Clad A被报道既能与兰科形成内生型菌根,又能与壳斗科、松科植物形成外生菌根[49],Clad B与杜鹃花科植物,如杜鹃花属、越橘属,帚石楠属等宿主共生。红菇目和革菌目是典型的外生菌根真菌,在美国大烟雾山国家公园Rhododendron maximum及澳大利亚Rhododendron hirsutum根系中也曾分离到红菇[7-9],中国云南的Rhododendron decorum根系中曾有革菌目的出现[10]。
群落物种丰富度的研究表明,受抽样样本大小的限制,在一个地区观察到的物种往往只能代表当地物种库中的一部分,然而,可以采用物种丰富度估计的方法来最大限度地估计当地物种数。本研究用Bootstrap估计的杜鹃花根系真菌物种数37种,高于实际观察到的物种数(28种)。物种累积曲线表明,随着杜鹃花宿主植物个体数增加,检测到的根系真菌物种数增加的潜力仍然较大,因此,全面揭示杜鹃花菌根真菌资源的多样性是杜鹃花菌肥开发利用的关键环节。
另外,本试验研究检测到的真菌有93%的物种只能鉴定到目,能鉴定到科、属、种水平的真菌仅占少数。主要是因为NCBI等公共数据库中暂时缺少与这些真菌高同源性,并且已知分类地位的代表性序列。随着NCBI数据库中分类信息准确的真菌DNA条形码序列累积,本研究检测到的真菌序列将会获得更精细的分类信息。
参考文献:
[1]Read D J. The structure and function of the ericoid mycorrhizal root[J]. Annals of Botany,1996,77(4):365-374.
[2]Stribley D P,Read D J. Biology of mycorrhiza in Ericaceae .4. Effect of mycorrhizal infection on uptake of N-15 from labeled soil by Vaccinium-Macrocarpon Ait[J]. New Phytologist,1974,73(6):1149-1155.
[3]Scagel C F. Inoculation with ericoid mycorrhizal fungi alters fertilizer use of highbush blueberry cultivars[J]. HortScience,2005,40(3):786-794.
[4]Sekar K C,Srivastava S K. Rhododendrons in Indian Himalayan region:diversity and conservation[J]. American Journal of Plant Sciences,2010,1(2):131-137.
[5]Bougoure D S,Cairney J W G. Fungi associated with hair roots of Rhododendron lochiae (Ericaceae) in an Australian tropical cloud forest revealed by culturing and culture-independent molecular methods[J]. Environmental Microbiology,2005,7(11):1743-1754.
[6]Fumiaki U,Junichi P A,Makoto K. Diversity of ericoid mycorrhizal fungi isolated from hair roots of Rhododendron obtusum var. kaempferi in a Japanese red pine forest[J]. Mycoscience,2003,44(2):97-102.
[7]Grunewaldt-Stcker G,von den Berg C,Knopp J,et al. Interactions of ericoid mycorrhizal fungi and root pathogens in Rhododendron:in vitro tests with plantlets in sterile liquid culture[J]. Plant Root,2013,7(0):33-48.
[8]Lin L C,Lee M J,Chen J L. Decomposition of organic matter by the ericoid mycorrhizal endophytes of Formosan Rhododendron (Rhododendron formosanum Hemsl.) [J]. Mycorrhiza,2011,21(5):331-339.
[9]Baird R,Wood-Jones A,Varco J,et al. Rhododendron decline in the Great Smoky Mountains and surrounding areas:intensive site study of biotic and abiotic parameters associated with the decline[J]. Southeastern Naturalist,2014,13(1):1-25.
[10]Sun L F,Pei K Q,Wang F,et al. Different distribution patterns between putative ercoid mycorrhizal and other fungal assemblages in roots of Rhododendron decorum in the southwest of China[J]. 2012,7(11):e49867.
[11]Tian W,Zhang C Q,Qiao P,et al. Diversity of culturable ericoid mycorrhizal fungi of Rhododendron decorum in Yunnan,China[J]. Mycologia,2011,103(4):703-709.
[12]Vohník M,Albrechtová J. The co-occurrence and morphological continuum between ericoid mycorrhiza and dark septate endophytes in roots of six European Rhododendron species[J]. Folia Geobotanica,2011,46(4):373-386.
[13]Zhang C Y,Yin L J,Dai S L. Diversity of root-associated fungal endophytes in Rhododendron fortunei in subtropical forests of China[J]. Mycorrhiza,2009,19(6):417-423.
[14]陈真,杨兵,张春英,等. 锦绣杜鹃菌根真菌 rDNA ITS 序列分析及接种效应研究[J]. 菌物学报,2011,30(5):729-737.
[15]李雪玲. 亮毛杜鹃根际真菌与内生真菌的多样性研究[J]. 楚雄师范学院学报,2005,19(3):87-90.
[16]刘仁阳,欧静,李冠楠,等. 梵净山雷山杜鹃根部真菌分离与鉴定[J]. 西北农业学报,2014,23(4):178-185.
[17]刘振华. 杜鹃花菌根真菌分离鉴定及多样性分析[D]. 北京:中国林业科学研究院,2010:1-71.
[18]刘振华,李潞滨,童方平,等. 灰背杜鹃菌根真菌的分离与鉴定及 DGGE 的应用[J]. 湖南林业科技,2012,39(4):16-20.
[19]姚娜,杨凯,王涛,等. 树枫杜鹃菌根真菌分离与鉴定[J]. 林业科学研究,2012,25(6):795-797.
[20]吴重华,王吉忍. 太白山自然保护区金背杜鹃菌根调查研究[J]. 西北林学院学报,2000,15(3):68-70.
[21]杨兵,张春英,王献,等. 杜鹃花根系内生菌资源调查及多样性分析[J]. 河南农业大学学报,2010 (3):290-294.
[22]郑钰,高博,孙立夫,等. 银叶杜鹃和繁花杜鹃根部真菌的多样性[J]. 生物多样性,2010,18(1):76-82.
[23]周诗涵,李明仁. 红毛杜鹃与内生菌 Phialocephala sp. 形成杜杜鹃菌根[J]. 中华林学季刊,2010,43(3):355-365.
[24]史佑海,李绍鹏,梁伟红,等. 海南野生杜鹃花属植物种质资源调查研究[J]. 热带作物学报,2010,31(4):551-555.
[25]李肇晨,罗微,陈永富,等. 海南霸王岭陆均松空间分布格局及其与微生境异质性的关系[J]. 生态学报,2015,35(8):2545-2554.
[26]胡玉佳,丁小球. 海南岛坝王岭热带天然林植物物种多样性研究[J]. 生物多样性,2000,8(4):370-377.
[27]陈玉凯,杨琦,莫燕妮,等. 海南岛霸王岭国家重点保护植物的生态位研究[J]. 植物生态学报,2014,38(6):576-584.
[28]Guo L D,Hyde K D,Liew E C Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences[J]. New phytologist,2000,147(3):617-630.
[29]贾锐. 兴安杜鹃菌根及其真菌多样性研究[D]. 呼和浩特:内蒙古农业大学,2011:1-37.
[30]Davies P,McLean C. Molecular investigation of sterile root-associated fungi from Epacris microphylla R. Br.(Ericaceae) and other epacrids at alphine,sub-alpine and coastal Heathland sites[J]. Australasian Mycologist,2004,23(3):94-104.
[31]Hynson N A,Bruns T D. Evidence of a myco-heterotroph in the plant family Ericaceae that lacks mycorrhizal specificity[J]. Proceedings of the Royal Society of London B:Biological Sciences,2009,276(1675):4053-4059.
[32]Obase K,Matsuda Y. Culturable fungal endophytes in roots of Enkianthus campanulatus (Ericaceae)[J]. Mycorrhiza,2014,24(8):635-644.
[33]Taylor D L,Hollingsworth T N,McFarland J W,et al. A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning[J]. Ecological Monographs,2014,84(1):3-20.
[34]Stenstrm E,Ndobe N E,Jonsson M,et al. Root-associated fungi of healthy-looking Pinus sylvestris and Picea abies seedlings in Swedish forest nurseries[J]. Scandinavian Journal of Forest Research,2014,29(1):12-21.
[35]Bergemann S E,Garbelotto M. High diversity of fungi recovered from the roots of mature tanoak (Lithocarpus densiflorus) in northern California[J]. Botany,2006,84(9):1380-1394.
[36]Poosakkannu A,Nissinen R,Kytviita M M. Culturable endophytic microbial communities in the circumpolar grass,deschampsia flexuosa in a sub‐Arctic inland primary succession are habitat and growth stage specific[J]. Environmental Microbiology Reports,2015,7(1):111-122.
[37]Bizabani C,Dames J F. Assimilation of organic and inorganic nutrients by Erica root fungi from the fynbos ecosystem[J]. Fungal Biology,2015,11(006):1-6.
[38]Jumpponen A. Dark septate endophytes-are they mycorrhizal?[J]. Mycorrhiza,2001,11(4):207-211.
[39]Mandyam K,Jumpponen A. Seeking the elusive function of the root-colonising dark septate endophytic fungi[J]. Studies in Mycology,2005,53(1):173-189.
[40]Bougoure D S,Parkin P I,Cairney J W G,et al. Diversity of fungi in hair roots of Ericaceae varies along a vegetation gradient[J]. Molecular Ecology,2007,16(21):4624-4636.
[41]Hazard C,Gosling P,Mitchell D T,et al. Diversity of fungi associated with hair roots of ericaceous plants is affected by land use[J]. FEMS microbiology ecology,2014,87(3):586-600.
[42]Toftegaard T,Iason G R,Alexander I J,et al. The threatened plant intermediate wintergreen (Pyrola media) associates with a wide range of biotrophic fungi in native Scottish pine woods[J]. Biodiversity and conservation,2010,19(14):3963-3971.
[43]Bunch W D,Cowden C C,Wurzburger N,et al. Geography and soil chemistry drive the distribution of fungal associations in ladys slipper orchid,Cypripedium acaule[J]. Botany,2013,91(12):850-856.
[44]Chambers S M,Curlevski N J A,Cairney J W G. Ericoid mycorrhizal fungi are common root inhabitants of non-Ericaceae plants in a south-eastern Australian sclerophyll forest[J]. FEMS microbiology ecology,2008,65(2):263-270.
[45]Midgley D J,Chambers S M,Cairney J W G. Spatial distribution of fungal endophyte genotypes in a Woollsia pungens (Ericaceae) root system[J]. Australian Journal of Botany,2002,50(5):559-565.
[46]Oliveira S F,Bocayuva M F,Veloso T G R,et al. Endophytic and mycorrhizal fungi associated with roots of endangered native orchids from the Atlantic Forest,Brazil[J]. Mycorrhiza,2014,24(1):55-64.
[47]Bougoure D S,Cairney J W G. Assemblages of ericoid mycorrhizal and other root‐associated fungi from Epacris pulchella (Ericaceae) as determined by culturing and direct DNA extraction from roots[J]. Environmental Microbiology,2005,7(6):819-827.
[48]Selosse M A,Bauer R,Moyersoen B. Basal hymenomycetes belonging to the Sebacinaceae are ectomycorrhizal on temperate deciduous trees[J]. New Phytologist,2002,155(1):183-195.
[49]Selosse M A,Setaro S,Glatard F,et al. Sebacinales are common mycorrhizal associates of Ericaceae[J]. New Phytologist,2007,174(4):864-878.
