地膜覆盖模式下芋头疫病发生规律及其与产量的关系
2016-07-25王安吴薇谢吉先焦庆清
王安+吴薇+谢吉先++焦庆清

摘要:由疫霉属真菌引起的芋头疫病是限制芋头产量的关键因素之一。以泰兴香荷芋为材料,设置不同地膜颜色及厚度10个处理,研究不同地膜覆盖处理下芋头疫病发生规律及其产量表现。研究结果表明,芋头疫病病情指数在不同处理间达到显著差异(P<0.05),D4、D7、D9这3个处理疫病病情指数均超过40,均显著高于其他各处理,表现为感病,D1、D2、D8的病情指数分别为26.21、26.35、26.50,表现为抗性;各处理间在倒一叶、倒二叶上疫病病级无显著差异,而在倒3叶上则有显著差异,叶位间疫病严重程度为:倒三叶>倒二叶>倒一叶;不同处理间芋头产量差异性较大,变幅为8 441(D10)~15 758 kg/hm2(D3);芋头病情指数与产量间的相关系数为-0.723,两者相关性达到极显著(P<0.01)。
关键词:芋头;疫病;产量
中图分类号: S436.32文献标志码: A文章编号:1002-1302(2016)06-0193-03
收稿日期:2016-02-03
基金项目:江苏省农业科技自主创新资金[编号:CX(13)3025];江苏省农业三新工程项目(编号:SXGC[2015]252)。
作者简介:王安(1988—),男,江苏连云港人,硕士,助研,主要从事特粮特经作物品种选育及其高效配套技术研究。E-mail:wangan_863@foxmail.com。
通信作者:焦庆清,硕士,副研究员,主要从事特粮特经作物新品种、新技术研究与开发应用研究。E-mail:mtjqq@163.com。疫病是由疫霉属真菌(Phytophthora)引起的植物病害[1],它是芋头生长过程中最严重的病害之一[2]。该病菌可以从植物的自然孔口或伤口侵入,在芋头感病早期表现为叶片病斑,褐色或黄褐色,后期斑面上下常长出白色霉状物(孢子囊及孢囊梗)及蜜黄色溢滴液,造成病叶干枯、破裂和穿孔,组织脱落,仅残留叶脉[3]。在田间主要通过气流、雨水和灌溉水传播,由于病程短,再侵染发生频繁,在多雨潮湿的条件下疫病流行速度特别快。疫病在全世界广泛分布,高温、高湿地区尤其猖獗,全球每年芋头产量因疫病造成减产20%~30%,严重年份可达50%以上[4-5]。在我国,疫病已成为芋头持续高产稳产的严重障碍,在长江中下游等雨水分布较多的地区发病尤为严重。由于疫霉属真菌存在生理分化现象,具有许多不同的生理小种,再加上连续阴雨天气,疫病防治工作一直是芋头生产中的一大难题。
一直以来,研究者通过选育芋头抗病品种作为防治疫病的主要手段[4,6],而在栽培方式对芋头疫病防治方面缺乏研究。目前,地膜覆盖是芋头大面积种植中应用比较广的栽培方式之一,关于地膜覆盖对芋头产量的研究报道有很多[7-9],而地膜覆盖条件下芋头疫病的发生与其抗性特征至今没有报道过。为此,本研究以香荷芋为材料,通过研究不同地膜处理下芋头疫病的发生规律,进而揭示地膜覆盖模式对芋头疫病防治机制,以期为实现芋头稳产高产提供参考。
1材料与方法
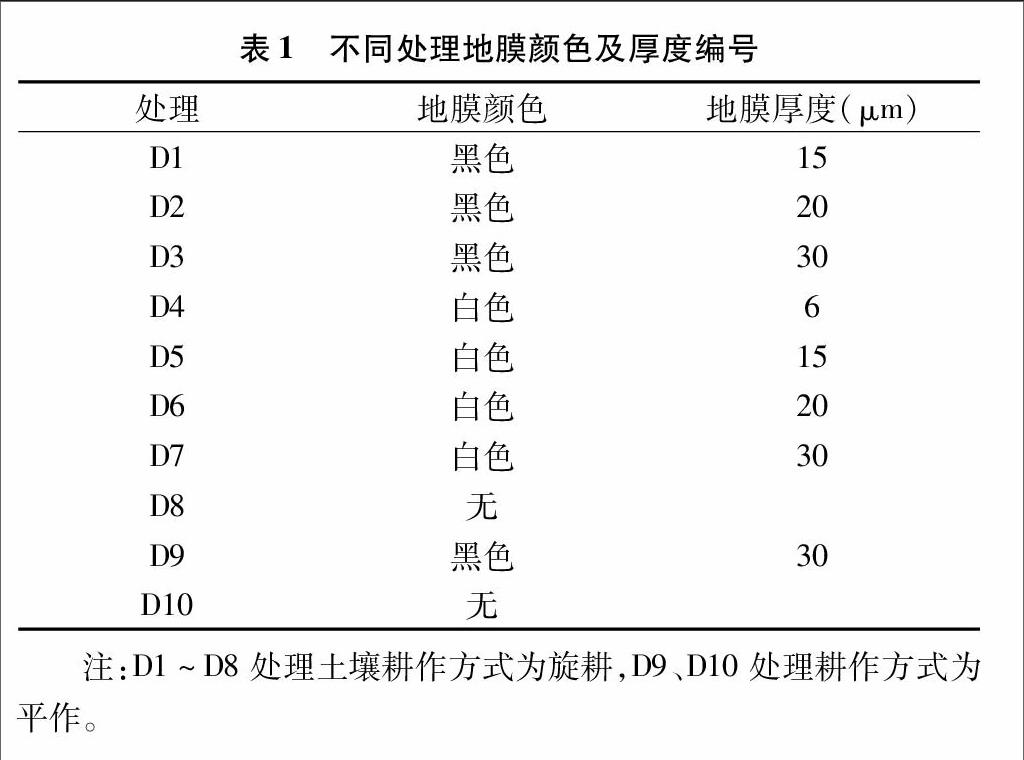
1.1试验设计
江苏省泰兴市农科所基地为试验点,该地区海拔在4~8 m,年平均气温在14.4~15.1 ℃之间,年平均降水量 1 000 mm 左右,降水主要集中在7—9月。土壤主要以沙土为主,地势平坦,排灌良好,肥力中等均匀。前茬水稻,收获后秸秆全量粉碎还田,冬季耕翻压草冻垡。2014年3月8日撒施有机复合肥4 500 kg/hm2,高钾复合肥900 kg/hm2,同时按表1方案进行设置。试验采用随机区组排列,重复3次,小区面积为1368 m2,小区间隔40 cm。垄宽为100 cm,高20 cm,垄面宽 60 cm,密度为52 620株/hm2,四周设置保护行。4月4日播种芋头,播种方式为穴播。其中,白色地膜覆盖小区播完种芋后覆上地膜,黑膜地膜覆盖小区覆膜后破膜播种种芋。4月20日灌水抗旱,5月6—8日陆续出苗,白膜小区选择晴天早晨或下晚及时放苗,5月15—18日陆续齐苗,5月25日查苗补缺。5月21日、6月25日、7月20日、8月23日4次人工拔除田间杂草,7月22日、8月10日进行培土及压边荷。8月22日对疫病进行统计调查,11月8日收获并计算各小区芋头子孙芋产量。
1.2疫病抗性鉴定
芋头疫病主要侵害叶片、叶柄及球茎。芋头疫病的抗性鉴定一般采用自然诱发鉴定,本病主要流行于降雨多发时期,鉴定部位主要以叶片为主,根据发病情况,将病情分为以下级别:0级,无病症;1级,零星坏死斑;2级,坏死面积不超过叶面积1/4;3级,坏死面积占叶面积的1/4~1/3;4级,坏死面积占叶面积的1/3~2/3;5级,坏死面积占叶面积的2/3以上。计算病情指数,公式为:
DI=∑(Si ni)/5N×100%。
式中:DI为病情指数;Si为发病级别;ni为相应发病级别的株数;i为病情分级的各个级别;N为总株数。
种质群体对疫病的抗性依病情指数分5级:1为高抗(HR)(DI≤10)、3为抗病(R)(10 8月18日芋头田出现疫病,8月22日疫病病害达到高峰,当日对芋头疫病严重程度进行调查,每个小区随机取样5株,根据叶片疫病发生面积对芋头全部叶片进行病害分级,并计算病情指数。并对倒一、倒二、倒三叶片疫病病级及叶片面积进行测量与分析,叶片面积采用公式A=L×W×0.75。其中A代表叶片面积,L代表叶片长度,W代表叶片宽度。 1.3数据分析 采用Excel 2007对疫病数据进行一般统计量分析,采用GraphPad Prism 5对疫病病级进行F测验,并对其进行Waller-Duncan新复极差法多重比较。
2结果与分析
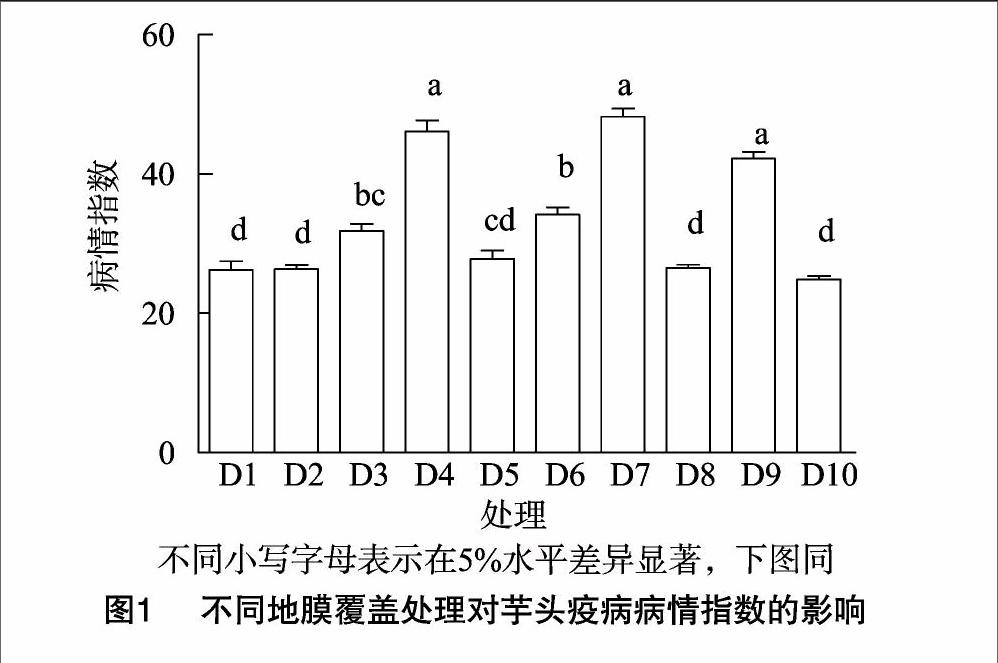
2.1芋头疫病在不同处理间的抗性表现
病情指数是衡量芋头疫病的主要指标之一,对芋头生长具有重要的指示作用。试验结果(图1)表明:D10处理芋头疫病病情指数最低,为24.85,表明该处理小区芋头种群对疫病表现为抗病(R),但还没有达到高抗(HR);D1、D2、D8与D10相比,病情指数无显著差异(P>0.05),但这3个处理病情指数分别为26.21、26.35、26.50,均属于中抗水平(MR)。D3、D6这2个处理群体病情指数DI分别为3184、3417,对疫病表现亦为中抗,但这2个处理病情指数与D1、D2、D8、D10有显著差异(P<0.05)。另外,D4、D7、D9这3个处理芋头群体对疫病的抗性表现为感病(S)(40 2.2叶位置疫病扩展差异性分析 不同地膜覆盖芋头倒三叶疫病级别特征分析结果(图2)表明:整体而言,疫病在芋头倒一叶的扩展表现为无病症或有零星坏死斑点,各处理间疫病病级无显著差异(P>0.05)。其中,D1、D5、D8这3种地膜覆盖模式下植株倒1叶(新叶)没有出现疫病;在倒二叶上,各处理芋头疫病均有小面积感染,疫病侵染面积均低于叶片总面积的1/3,其病级变幅在12(D4)~2.6(D7),各处理间疫病病级没有显著差异;疫病在倒三叶上的抗性表现有很大差异性,方差分析表明处理间疫病病级达到极显著差异(P<0.01),其中,病级最高的为D4,为4.8,几乎所有叶面积均出现坏死,病级最小的是D1,其大小为1.6。另外,从图2可以看出,叶位置对芋头疫病表现也有一定影响,叶位置疫病侵染严重程度为:倒三叶>倒二叶>倒一叶,可能原因是芋疫霉菌寄主中心区位于植株下半部老叶区,随着叶位的上升,疫病病级逐步减小。 2.3不同处理芋头子孙芋产量表现 不同处理小区芋头子孙芋产量结果(图3)表明:不同处理间小区芋头产量差异性较大,变幅为8 441(D10)~15 758 kg/hm2(D3)。D2、D3和D9小区芋头产量分别为 14 248、15 758、14 360 kg/hm2,这3个黑膜覆盖处理芋头产量均显著高于白膜覆盖处理(D4~D7)及无地膜处理(D8、D10)(P<0.05),表明黑膜处理能显著提高芋头产量。黑色地膜处理间(D1~D3、D9)芋头产量无显著差异(P>0.05),表明黑膜不同厚度对芋头产量的影响较小,而在白膜覆盖处理间,D4、D5、D7处理小区芋头产量依次降低,分别为 11 802、11 234、10 402 kg/hm2,表明芋头产量随着白膜处理厚度的增加而下降,可能原因是白膜厚度的增加会导致土壤温度的升高,进一步导致土壤含水量的下降,从而引起芋头植株产量表现受抑制。露地处理下(D8、D10)芋头产量最低,表明无地膜覆盖不利于芋头产量的提高。 2.4芋头疫病病情指数与子孙芋产量的相关性分析 对芋头疫病病情指数与子孙芋产量进行的相关性分析(图4)表明:芋头病情指数与产量间的相关系数为-0.722,两者相关性达到极显著水平(P<0.01),表明芋头疫病病情指数与产量为极显著负相关,病情指数越高,产量越低。由产量和病情指数的回归曲线y=-224.5x+20 089可知,当疫病病情指数低于40时,芋头子孙芋产量较高,高达 11 000 kg/hm2,而当病情指数高于50时,子孙芋产量则低于8 800 kg/hm2,表明疫病病害越重,产量越低。 3讨论 芋头疫病在我国长江中下游地区分布较为广泛,该地区每年6—8月月平均降水量高于150 mm,日平均温度高于255 ℃。温和湿润的环境为疫病的发生、流行创造了自然条件。在此环境下,疫病病菌的前体孢子囊借助风雨传播,产生芽管或游动孢子侵染芋头植株,后又在病部产生孢子囊进行再侵染。芋疫霉菌侵染芋头是一个受多因素影响的复杂过程,该过程受到田间菌源量、品种抗性、田间小气候、栽培管理措施等多种因素的影响。在芋头疫病抗性鉴定方面,国际上的研究报道有很多[10-12],而在国内相关的研究报道较少。莫俊杰等于2011年及2012年对63个芋头品种的疫病抗性鉴定及其不同品系遗传多样性进行分析,发现不同品系间抗病性差异显著,广州-2、江门-2、湛江-5等6个品系对疫病表现为抗病,而潮州-1、惠州-1、惠州-2等42品系对疫病表现为感病,占总数的66.7%[13]。 目前,国内研究者研究的主要方向为芋头疫病的药物防治。例如,朱敦军在50%烯酰吗啉水分散粒剂防治芋头疫病药效试验中发现,采用剂量为1 000倍液,作物生长周期内使用3~4 次,间隔为5~6 d,对芋头疫病有很好的控制作用[14];马建芳等通过不同药剂试验筛选出25%双炔酰菌胺悬浮剂对芋艿疫病的防效最好,显著优于氟菌·霜霉威等药剂[15]。然而,由于药剂的使用会对环境、人体产生危害,因此对芋头疫病的防治还应注重其他防治措施。由于疫病病害受到田间连作、低洼、排水不良、通风透光不良等栽培措施的影响[16],而目前在这些方面的研究还比较缺乏。因此,本研究以地膜覆盖模式为切入点,重点分析了不同地膜覆盖处理下芋头疫病的发病规律,针对倒一、倒二、倒三叶的疫病病级的病情特征进行了较为详细的研究,并对其与芋头子孙芋产量进行了相关性分析,这是本研究的一大创新点。当然,本研究仅仅是对1年间芋头疫病发生规律的研究,还需对其进行多年间的试验研究,以得到稳定可靠的数据结果,这将是下一步的工作重点。 参考文献:
[1]van der Plank J E. Plant diseases:epidemics and control[M]. Amsterdam,Netherlands:Elsevier,2013.
[2]Pillai S V,Tkankappan M. Breeding for leaf blight resistance in taro-problems and prospects[J]. Journal of Root Crops,1991,3:57-61.
[3]王德旭. 芋头疫病识别及防治[J]. 植物医生,1995,1(8):16.
[4]Sahoo M R,DasGupta M,Kole P C,et al. Antioxidative enzymes and isozymes analysis of taro genotypes and their implications in Phytophthora blight disease resistance[J]. Mycopathologia,2007,163(4):241-248.
[5]Misra R S,Sharma K,Mishra A K. Phytophthora leaf blight of taro (Colocasia esculenta)-a review[J]. The Asian and Australasian Journal of Plant Science and Biotechnol,2008,2:55-63.
[6]Sahoo M,Sahu S,Mukherjee A,et al. In vitro screening of taro genetic resources for tolerance to biotic and abiotic stresses[J]. Plant Resources Utilization,2002,4:241-248.
[7]Miyasaka S C,Hollyer J R,Kodani L S. Mulch and compost effects on yield and corm rots of taro[J]. Field Crops Research,2001,71(2):101-112.
[8]Pardales Jr J R. Effect of mulch application and planting depth on growth,development and productivity of upland taro(Colocasia esculenta)[J]. Annals of Tropical Research,1985,7:27-38.
[9]Asghar M,Fernando L H,Kaye A,et al. Effects of mulch,manure and fertilizer on taro [Colocasia esculenta (L.) Schott] production at Alafua,Western Samoa[J]. Alafua Agricultural Bulletin,1985,10(2):37-46.
[10]Brooks F E. Detached-leaf bioassay for evaluating taro resistance to Phytophthora colocasiae[J]. Plant Disease,2008,92(1):126-131.
[11]Jackson G V H,Gollifer D E,Newhook F J. Studies on the taro leaf blight fungus Phytophthora colocasiae in Solomon Islands:Control by fungicides and spacing[J]. Annals of Applied Biology,1980,96
(1):1-10.
[12]Bandyopadhyay R,Sharma K,Onyeka T J,et al. First report of taro (Colocasia esculenta) leaf blight caused by Phytophthora colocasiae in Nigeria[J]. Plant Disease,2011,95(5):618.
[13]莫俊杰,胡汉桥,梁钾贤,等. 芋疫病抗病性鉴定及不同品系遗传多样性分析[J]. 广东海洋大学学报,2012,32(4):67-72.
[14]朱敦军. 50% 烯酰吗啉水分散粒剂防治芋头疫病药效试验[J]. 现代农业科技,2009(13):151-151.
[15]马建芳,张仁杰,周华光,等. 奉化大芋艿疫病防治药剂筛选初报[J]. 浙江农业科学,2014(2):230-232.
[16]姜东明. 2012 年芋头疫病大发生原因分析及防治对策[J]. 福建农业科技,2012(12):25.王伟舵,刘永锋. 中国稻瘟病菌遗传多样性研究进展[J]. 江苏农业科学,2016,44(6):196-198.
