苦荞杂交后代主花序特征遗传变异研究
2016-07-24梁龙兵陈其皎石桃雄梁成刚张启迪饶庆琳陈庆富
梁龙兵,陈其皎,石桃雄,梁成刚,张启迪,唐 链,饶庆琳,陈庆富
(贵州师范大学 荞麦产业技术研究中心,生命科学学院 植物遗传育种研究所,贵州 贵阳 550001)
苦荞杂交后代主花序特征遗传变异研究
梁龙兵,陈其皎,石桃雄,梁成刚,张启迪,唐 链,饶庆琳,陈庆富*
(贵州师范大学 荞麦产业技术研究中心,生命科学学院 植物遗传育种研究所,贵州 贵阳 550001)
以小米荞为母本、晋荞2号为父本进行有性杂交,对群体F2、F3群体主花序二分叉花枝数和顶三花枝种子数进行了遗传变异分析和相关性分析。结果表明,苦荞杂交后代主花序二分叉花枝数广义遗传力、狭义遗传力平均值分别为0.690、0.314,主花序顶三花枝种子广义遗传力、狭义遗传力平均值分别为0.729、0.375。相比之下,主花序顶三花枝种子数遗传力大于主花序二分叉花枝数。主花序二分叉花枝数与主花序顶三花枝种子数呈微弱的负相关(-0.141**),可能是顶端优势相互制约的结果。F2主花序二分叉花枝数和主花序顶三花枝种子数与单株粒质量、单株粒数表型相关系数分别为0.317**和0.256**、0.344**和0.278**,F3表型相关系数为0.316**和0.272**、0.371**和0.516**。表明主花序二分叉花枝数和顶三花枝种子数均与单株粒数和单株粒质量呈极显著正相关。因此,在育种上应考虑选择主花序二分叉花枝数、顶三花枝种子数多的变异类型以提高产量。
苦荞; 主花序二分叉花枝数; 主花序顶三花枝种子数; 遗传参数
苦荞(FagopyrumtataritumGartn.)为蓼科(Polygonaceae)荞麦属(FagopyrumMill)栽培品种[1-2],具有营养保健和食疗双重功效,是国际粮农组织公认的粮药兼用作物,被誉为五谷之王、三降 (降血压、降血糖、降血脂)食品[3]。苦荞资源在我国广为分布,但集中于黄土高原和云贵高原的山西、陕西、青海、甘肃、云南、贵州、西藏、四川及其毗邻的鄂西北和湘西的武陵山区[4-5]。苦荞生长期发育快、耐贫瘠、生育期短,能作为其他大宗粮食作物因自然灾害不能补种的作物,具有特殊的农业地位。
由于苦荞杂交育种工作滞后,目前生产上使用的主要还是农家品种和系统育种培育的品种。苦荞的产量优势还未体现出来,严重制约苦荞的生产和开发利用。近年来,苦荞的遗传育种取得一定的进展[6-10]。国内外关于荞麦产量主要受分枝数、单株粒数以及单株粒质量等因素影响的报道较多[11-18]。而对主花序顶三花枝种子数及主花序二分叉花枝数2个性状的遗传研究尚未见报道,为此,本研究拟用晋荞2号和小米荞为研究材料,基于遗传型方差、变异系数、遗传力、遗传相关等遗传参数对苦荞后代主花序二分叉花枝数、顶三花枝种子数进行遗传变异分析,从而了解其性状的遗传规律,以提高选择效果,增加对杂交后代性状表现的预见性。
1 材料和方法
1.1 材料
供试材料亲本由贵州师范大学荞麦产业技术研究中心经选择纯化后提供,小米荞和晋荞2号2个苦荞品种的原产地分别为云南和山西。晋荞2号瘦果厚壳有沟槽不易脱壳,迟熟、抗旱,抗倒伏,耐贫瘠,田间生长势好,是既高产又稳产的品种[19-20];小米荞具有黄酮含量高、遗传稳定、种壳簿且易去壳、出粉率高等优点,可用于作为生产苦荞米和苦荞黄酮类医药保健产品的专用品种[21]。
2012年7月播种供试材料亲本于贵阳市观水基地用于杂交。杂交种子播种后生长得到杂种植株,进行严格自花授粉,经自交获得杂种种子。将3个不同母本植株获得的3个杂交后代群体分别取代号A、B、C。在贵阳市柏杨基地,将杂种植株所结种子稀播,获得杂种F2分离群体。该群体按单株收获,再次播种形成F3株系。亲本、杂种、杂种F2、F3群体栽培均采用株间距5 cm、行间距33~40 cm。
1.2 性状考察
2014—2015年苦荞成熟期对亲本、F2和F3株系群体主花序顶三花枝种子数、主花序二分叉花枝数、单株粒质量和单株粒数等性状进行考种和记录,其中对F3各株系随机选10株进行考种,取其10株平均数为F3株系的数值。
1.3 统计分析
试验数据用Excel 2010和SPSS 17.0进行统计和相关分析。杂交后代目标性状的遗传力、表型遗传相关和遗传变异系数等遗传参数参考文献[21-26]的计算方法。
2 结果与分析
2.1 苦荞亲本主花序性状的变异度分析
对苦荞亲本主花序及其单株粒质量变异度分析(表1)结果表明,小米荞主花序二分叉花枝数和主花序顶三花枝种子数平均值均高于晋荞2号,而单株粒质量性状为小米荞<晋荞2号,主花序二分叉花枝数变异系数为小米荞(0.32)<晋荞2号(0.64),主花序顶三花枝种子变异系数为小米荞(0.59)>晋荞2号(0.54),说明双亲在主花序二分叉花枝数和顶三花枝种子数上有一定差异。
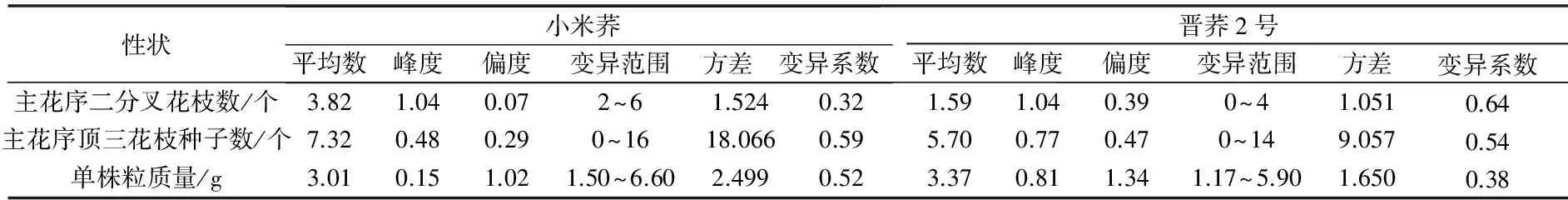
表1 苦荞亲本主花序及其单株粒质量变异分析
2.2 苦荞F2植株与对应F3株系群体性状分析
群体目标性状数据如表2所示,主花序二分叉花枝数变异总范围表现为F2(0~15)比F3(0~22)小,其平均数也小于F3株系;主花序顶三花枝种子数F2平均数比F3大,主花序顶三花枝种子数变异范围为F2(0~72)明显大于F3(0~26)。

表2 苦荞杂种F2和F3主花序二分叉花枝数和主花序顶三花枝种子数分析
2.3 苦荞3个群体表型和遗传变异系数及遗传力分析
苦荞杂交后代3个群体的表型及遗传的变异系数分析如表3所示。主花序二分叉花枝数F2、F3群体表型变异范围分别为55%~89%、35%~46%,平均数分别为72%、40%,而其遗传变异系数平均数分别为54%、34%,广义遗传力和狭义遗传力平均值分别为0.690、0.314;主花序顶三花枝种子的F2、F3群体表型变异系数分别为65%、84%,而遗传变异系数为55%、73%,其广义遗传力和狭义遗传力分别为0.729、0.375。相比之下,主花序顶三花枝种子数遗传力大于主花序二分叉花枝数。

表3 苦荞杂交后代F2和F3性状的变异系数及其遗传力分析
2.4 F2代与F3株系3个群体表型相关与遗传相关分析
F2群体4个性状的表型相关和遗传相关系数如表4所示。表型相关和遗传相关数值方向一致,遗传相关大于表型相关,主花序二分叉花枝数与主花序顶三花枝种子表型相关系数呈负相关,但相关系数很小,其相关系数平均为-0.297**。各个性状均对单株粒质量呈极显著正相关,其中主花序二分叉花枝数和主花序顶三花枝种子数均与单株粒质量、单株粒数表型相关系数平均分别为0.317**和0.256**、0.344**和0.278**,主花序目标性状对单株粒质量有极显著正影响。

表4 苦荞F2群体表型相关与遗传相关分析
注:每个群体都以1.000为对角线,对角线上三角为遗传相关系数rg,对角线下三角为表型相关系数rP。
F3群体表型相关系数与遗传相关系数之间的关系和F2相似,而表型相关中,主花序二分叉花枝数与单株粒质量、单株粒数极显著正相关,主花序二分叉花枝数与主花序顶三花枝种子数呈微弱的负相关(-0.141**)。

表5 苦荞F3群体表型相关与遗传相关分析
3 结论与讨论
杨玉霞等[27]研究5个国家55个苦荞品种有效花序和分枝数为46.94%、30.86%,发现有效花序变异较大。罗燕等[28]的研究发现,苦荞主花序种子数变异系数为33.66%。而目前尚未发现主花序二分叉花枝数的报道。本研究发现,小米荞和晋荞2号两苦荞品种后代F2、F3主花序顶三花枝种子数表型变异系数为65%、84%,可能暗示杂交后代更有选择潜力。本研究发现,苦荞主花序二分叉花枝数和顶三花枝种子数表型变异系数和遗传变异系数均不同程度的变化,随着群体数目增大而变化,表明群体性状随着世代加深趋于稳定,可选择性减小,可达到自交育种目的。江满霞等[29]对13个油菜品种的农艺性状遗传力分析得出,第1次有效分枝部位和第1次有效分枝数、主茎长的遗传力较高,因此根据遗传力大小可以提高选择效果和选择预见性。本研究得出主花序二分叉花枝数、顶三花枝种子数广义遗传力为0.690、0.729,狭义遗传力为0.314、0.375。表明亲代将这些性状传递给子代能力大。狭义遗传力较小,显性作用为主,可能出现超亲现象。李秀莲等[30]报道,二级分枝与单株粒质量的相关系数rP=0.371**,rg=0.453**;吴渝生[31]分析荞麦主要农艺性状得出,分枝数与单株粒质量相关系数rP=0.329,rg=0.388。而汪灿等[32]报道分枝数与单株粒质量相关系数为r=0.191 7,表明遗传分析得出的相关更能揭示性状的关联。本研究发现,F2、F3主花序二分叉花枝数与主花序顶三花枝种子数相关系数在平均水平上均呈极显著相关(F2为-0.297**,F3为-0.141**),可能是顶端优势相互制约的结果。且F2和F3主花序二分叉花枝数和主花序顶三花枝种子数均与单株粒质量呈极显著相关,优选主花序二分叉花枝数和主花序顶三花枝种子数作为育种的目标。李月等[33]对苦荞农艺性状与产量的相关分析结果为单株粒数与单株粒质量的简单相关系数r=0.987**,偏相关系数为r=0.983**。本研究该相关性分析F2群体与之相似,F3相关系数变小,可能因为是粒型发生变化而改变。目前尚未发现苦荞主花序顶三花数和主花序二分叉花枝数遗传研究,综上所述,在育种上考虑选择主花序二分叉花枝数、顶三花枝种子数多的变异类型,可以提高产量。
[1] 陈庆富.荞麦属植物科学[M].北京:科学出版社,2012.
[2] 林汝法.中国荞麦[M]北京:中国农业出版社,1994:160.
[3] 万丽英,穆建稳.贵州苦荞的营养保健功能与开发利用价值[J].贵州农业科学,2004,32(2):74-75.
[4] 赵钢,陕方.中国苦荞[M].北京:科学出版社,2009.
[5] 林汝法.苦荞举要[M].北京:中国农业科学技术出版社,2013.
[6] Fesenko I N.Non-shattering accessions ofFagopyrumtataricumGaertn.carry recessive alleles at two loci affecting development of functional abscission layer[J].Fagopyrum,2007,23(2):7-10.
[7] Wang Y J,Campbell C G.Tartary buckwheat breeding(FagopyrumtataricumGaertn.) through hybridization with its rice-tartary type[J].Euphytica,2007,156(3):399-405.
[8] Mukasa Y,Suzuki T,Honda Y.Suitability of rice-tartary buck-wheat for crossbreeding and for utilization of rutin[J].Jpn Agr Res Quart,2009,43(3):199-206.
[9] Li C H,Kobayashi K,Yoshida Y,etal.Genetic analyses of agronomc traits in Tartary buckwheat(Fagopyrumtartaricum(L.) Gaertn.)[J].Breed Sci,2012,62(4):303-309.
[10] 李春花,大泽良,小林喜和.苦荞杂交后代优良株系筛选研究[J].植物遗传资源学报,2015,16(1):168-172.
[11] 王安虎.苦荞杂交F1代的植物学性状表型研究[J].西南大学学报(自然科学版),2012,34(6):1-6.
[12] 王安虎.苦荞杂交F2代的植物学性状表型研究[J].西南大学学报(自然科学版),2012,34(6):12-17.
[13] 杜晓磊,张宗文,吴斌,等.苦荞 SSR分子遗传图谱的构建及分析[J].中国农学通报,2013,29(21):61-65.
[14] 陈庆富,陈其饺,石桃雄,等.苦荞厚果壳性状的遗传及其与产量因素的相关性研究[J].作物杂志,2015(2):27-31.
[15] 陆时万,徐祥生,沈敏健.植物学[M].北京:高等教育出版社,1991:109-112.
[16] 欧阳羽彬.库伦荞麦栽培技术[J].作物栽培,2000(3):21.
[17] 周长军.大豆有性杂交F2产量性状的遗传力分析[J].黑龙江农业科学,2006,15(6):14-15.
[18] 林国强,徐树传,黄建成,等.大豆不同亲本类型F2主要性状遗传参数分析[J].植物遗传资源科学,2001,2(2):12-15.
[19] 李秀莲,史兴海,高伟,等.苦荞新品种‘晋荞麦2号’丰产稳产性分析及应用前景[J].农学学报,2011,1(12):6-10.
[20] 王跃华,陈燕,张红玉,等.米荞种子萌发和组织培养研究[J].西南农业学报,2012,25(3):1043-1046.
[21] 姚世鸿,王景佑,陈庆富.遗传学[M].贵阳:贵州人民出版社,2001:157-175.
[22] 陈庆富.生物统计学[M].北京:高等教育出版社,2011.
[23] 李春喜,邵云,姜丽娜.生物统计学[M].北京:科学出版社,2008.
[24] 孔繁玲.植物数量遗传学[M].北京:中国农业大学出版社,2006.
[25] 马惠馨,吴炳玉,王淑贤,等.辣(甜)椒杂种F2数量性状遗传相关与遗传的研究初报[J].沈阳农业大学学报,1988,19(1):15-20.
[26] 杨光宇,郑惠玉,韩春风.栽培大豆(G.max)×半野生大豆(G.gracilis)后代主要农艺性状遗传参数的初步分析[J].作物学报,1992,18(6):439-446.
[27] 杨玉霞,吴卫,郑有良,等.苦荞主要农艺性状与单株籽重的相关和通径分析[J].安徽农业科学,2008,36(16):6719-6721.
[28] 罗燕,敖学成.秋播苦荞麦单株性状与株产种量的相关通径分析[J].牧草与饲料,2012,6(2):31-34.
[29] 江满霞,高冰可,卢列健,等.13个油菜品种的农艺性状遗传力和相关性分析[J].浙江农业科学,2015,56(4):472-474.
[30] 李秀莲,林汝法,乔爱花.苦荞主要性状的相关及通径分析[J].国外农学—杂粮作物,1997(2):26-28.
[31] 吴渝生.荞麦主要农艺性状的遗传相关分析[J].云南农业大学学报,1996,11(4):258-262.
[32] 汪灿,胡丹,杨浩,等.苦荞主要农艺性状与产量关系的多重分析[J].作物杂志,2013(6):18-22.
[33] 李月,石桃雄,黄凯丰,等.苦荞生态因子及农艺性状与产量的相关分析[J].西南农业学报,2013,26(1):34-41.
Genetic Analysis of Main Inflorescence Traits of Hybridization Progeny of Tartary Buckwheat
LIANG Longbing,CHEN Qijiao,SHI Taoxiong,LIANG Chenggang,ZHANG Qidi,TANG Lian,RAO Qinglin,CHEN Qingfu*
(Research Center of Buckwheat Industry Technology/Institute of Plant Genetics and Breeding,School of Life Science,Guizhou Normal University,Guiyang 550001,China)
This experience took Xiaomiqiao as female and Jinqiao No.2 as male through hybridization.The F2,F3populations obtained from hybrids of the tartary buckwheat selfing.The genetic variation analysis and correlation analysis of both main inflorescence dichotomy number and top three seed number of main inflorescence branches were done.The results showed that the broad-sense heritability estimates of main inflorescence dichotomy number and top three seed number of main inflorescence branches were 0.690,0.729,and narrow-sense heritability estimates were 0.314,0.375 respectively.In contrast,the heritability of top three seeds of main inflorescence branches was greater than that of the main inflorescence dichotomy number.There was a little negative correlation between main inflorescence dichotomy number and top three seed number of main inflorescence branches.The phenotypic correlation coefficients between F2main inflorescence dichotomy number,top three seeds number of main inflorescence branches and seed weight per plant,grain number were 0.317**,0.256**and 0.344**,0.278**,respectively; which of F3were 0.316**,0.272**and 0.371**,0.516**.The above results indicated that main inflorescence dichotomy number and top three seed number of main inflorescence branches were significantly corellated with number of seeds per plant and seed yield per plant.Therefore,the variation types with more main inflorescence dichotomy number and top three seed number of main inflorescence branches should be selected in breeding,so as to increase production.
tartary buckwheat; inflorescence dichotomy number; top three seeds number of main inflorescence branches; genetic paramete
2015-10-27
国家燕麦荞麦现代农业产业技术体系专项(CARS-08-A4);国家自然科学基金项目(31060207,31171609);贵州省高层次创新型人才培养计划项目(黔科合人才[2015]4020号)
梁龙兵(1988-),男,贵州贞丰人,在读硕士研究生,研究方向:荞麦遗传育种。E-mail:939320641@qq.com
*通讯作者:陈庆富(1966-),男,贵州贵阳人,教授,博士生导师,主要从事荞麦遗传育种研究。E-mail:cqf1966@163.com
S517
A
1004-3268(2016)05-0013-05
