黄龙山天然次生林辽东栎种群空间格局
2016-07-21王道亮李卫忠王广儒
王道亮,李卫忠,*,曹 铸,王 庆,王广儒
1 西北农林科技大学, 林学院,杨凌 712100 2 延安市黄龙山林业局,黄龙 715700
黄龙山天然次生林辽东栎种群空间格局
王道亮1,李卫忠1,*,曹铸1,王庆1,王广儒2
1 西北农林科技大学, 林学院,杨凌712100 2 延安市黄龙山林业局,黄龙715700
摘要:种群的空间分布格局是研究种群特征、种间相互作用以及种群与环境关系的重要手段。为更好地了解黄龙山天然次生林内辽东栎(Quercus wutaishanica)的种群特征,在该林区中油松(Pinus tabulaeformis)-辽东栎-白桦(Betula platyphylla)天然次生混交林和辽东栎天然次生纯林内分别选取并设立1块1 hm2的样地。根据样地调查规范对两块样地中的乔木进行空间定位并进行每木调查。研究参照静态生命表,采用径阶替代年龄的方法,将辽东栎种群不同生长阶段的分段问题转换为根据胸径(DBH)的大小划分生长阶段。研究中将辽东栎划分为4个不同生长阶段:幼龄阶段A, 3 cm ≤DBH<10 cm;中龄阶段B, 10 cm ≤DBH<20 cm;近熟龄阶段C, 20 cm ≤DBH<30 cm;成过熟龄阶段D, DBH≥30 cm。利用O-ring统计分析方法的手段,分析2 块样地中辽东栎的种群空间分布格局。本次研究分别采用单变量、双变量分析方法分析各生长阶段的空间格局、不同生长阶段间相互关联性。结果表明:(1) 辽东栎种群在两块样地内的径级结构均符合倒“J”型,从径级分布角度说明两块样地的天然更新状况都良好;(2) 辽东栎的空间分布格局与尺度有密切关系, 两块样地在大、中尺度水平上基本表现为随机分布,但是在较小尺度上均表现为聚集分布,在样地Ⅰ中0—6 m尺度上尤为突出;(3) 同一样地内各生长阶段的空间分布格局不同,幼龄个体在小尺度均表现为聚集分布,在样地Ⅱ中0—11 m尺度上尤为突出;(4)样地中不同生长阶段间相互关联性也不尽相同,样地Ⅰ中各生长阶段间关联性均不明显;样地Ⅱ中,幼龄的个体与中龄的个体间在小尺度上表现出显著的正相关性, 而与近熟个体和成、过熟个体在小尺度上表现为明显的负相关性。在2—5 m尺度上,中龄个体与成过熟个体也表现出一定的负相关性。辽东栎种群是黄龙山天然次生林重要的建群种之一,研究其不同生长阶段个体的空间分布规律有助于掌握黄龙山区辽东栎生长现状;研究不同生长阶段的空间关联性有利于深入了解其群落的发展趋势,并为对种群聚集性较大的区域开展结构化森林经营提供重要依据。
关键词:天然次生林;空间分布格局;空间关联性;O-ring统计分析;辽东栎
物种是森林结构的重要组成,其发育的形态不仅与所包含的物种相关,更与当地的自然环境因子有重要关系[1]。森林的物种组成特征一方面表征着森林的发育程度,另一方面也与森林生态功能的发挥有着不可分割的联系[2- 4]。种群空间分布格局是在种内竞争、种间竞争及自然环境等各种因素共同影响下形成的结果[5- 7],对其展开深入分析,不仅可以对物种分布规律有一定的了解,还可以分析推断其发育过程[4,8- 9],是研究种群特征、种间相互作用以及种群与环境关系的重要手段[2,5],有利于更好地理解种群格局形成的潜在生态过程[5]。
黄龙山林区地处黄土高原沟壑区,为陕西渭北旱塬与陕北黄土高原的过渡带, 被誉为“黄土高原的绿色明珠”,是我国水土保持的重要防护林区。辽东栎是黄龙山林区重要的优势种之一,属暖温带落叶阔叶栎林的重要树种[10]。
目前,在黄龙山林区,对辽东栎的研究内容主要集中于生态位[11]、生物多样性[12- 13]、群落稳定性[14]等方面,对于其不同生长阶段的空间分布及其空间关联性尚未见报道。以往文献主要以胸径为依据,通过丛生法[15]、方差/均值比法[16]、g(r)函数法[17- 18]等方法对物种空间格局分布进行研究。O-ring函数在优势种群的空间分布及种间关联性研究方面日渐多见[5,19- 20],而将O-ring函数运用于单一树种不同生长阶段之间关联性的文章却不多见[21]。本次研究选取黄龙山林区内辽东栎林和油松-辽东栎-白桦混交林,分别选设一块1 hm2级样地,并对符合调查条件的乔木进行每木检尺。本文根据辽东栎径阶,结合静态生命表[22],按照龄组的划分方法,将其划分为不同生长阶段,并运用O-ring点格局分析法进行空间格局分析及空间关联性分析,以期对种群聚集性较大的区域实施结构化森林经营。
1研究区概况
研究区位于陕西省黄龙山天然林保护区内,地理坐标为东经109°38′49″—110°12′47″ E,北纬35°28′46″—36°02′01″ N,位于温带半湿润与半干旱气候的过渡地带,属黄土高原丘陵沟壑区,海拔1100—1300 m,年平均降水量为611.8 mm,年均气温为8.6 ℃,地带性土壤为褐土和灰褐色森林土[23],森林植被属暖温带落叶混交林地带,主要优势种包括油松、辽东栎、白桦等,天然植被覆盖率高。
2研究方法
2.1样地设置及调查
根据空间格局研究样地的要求,样地必须有一定的规模才能保证物种的各种格局均能出现[4],且样地应以长方形或正方形最佳[24]。2013 年8 月,根据样地设置技术要求及研究目标[25],通过对虎沟门林场进行全面踏查,在该林场设立了2 块辽东栎生长状况良好并且具有典型特征的样地:样地Ⅰ油松-辽东栎-白桦混交林样地和样地Ⅱ辽东栎纯林样地,其样地面积均为1 hm2。采用相邻网格法,用高精度的拓普康全站仪将样地划分为25 块20×20 m2的小样地,对两块样地内乔木层展开调查,主要调查因子包含树种、胸径(DBH) (cm)、树高(m)、冠幅(m)、生长状态等,同时调查记录各林木的空间位置和坐标以及环境因子如土壤、枯落物等,并进行逐株挂牌。样地统计结果如表1。
根据调查结果可知:样地Ⅰ中,乔木层主要由油松、辽东栎、白桦等组成,灌木层主要由华北紫丁香(Syringaoblata)、茶条槭(Acerginnala)等组成,草本主要由大披针苔草(Carexlanceolata)、高羊茅(Festucaarundinacea)等组成。样地Ⅱ中,乔木层主要由辽东栎组成,灌木层主要由野山楂(Crataeguscuneata)、土庄绣线菊(Spiraeapubescens)等组成,草本层主要由大披针苔草、高羊茅、紫菀(Astertataricus)等组成。

表1 样地调查概况表
辽东栎树体高大,木质坚硬,因而不适合利用生长锥法等传统方法进行年龄的测定[26],而在相似立地条件和自然环境下的相同物种其龄级和径级对环境的反映规律具有一致性[27],样地调查开始之前,对样地林分平均胸径进行估测。经初步预测,本研究选取的两块样地平均胸径均不大于12 cm,结合林分结构规律[28],以林分平均胸径的0.4 倍所在径阶作为起测径阶。因此,文本起测径阶为4径阶,从而确定起测胸径为3.0 cm。根据样地调查结果,结合辽东栎种群静态生命表[22],将其划分为A—D 4个阶段:幼龄阶段A, 3 cm ≤DBH<10 cm;中龄阶段B, 10 cm ≤DBH<20 cm;近熟龄阶段C, 20 cm ≤DBH<30 cm;成过熟龄阶段D, DBH≥30 cm。其分布结果如图1。

图1 辽东栎空间分布图Fig.1 Spatial distribution of Quercus wutaishanica
2.2数据处理
点格局分析法(point pattern analysis)是以植物种的个体在空间的坐标为基本数据,以其空间分布点图为基础进行格局分析。该方法广泛应用于植物种群多尺度空间分布格局及两个物种之间多尺度空间关联的研究。
O-ring函数是在RipleyK(d)函数的基础上,结合Mark相关函数的一种新的点格局分析方法[5]。O-ring采用环形尺度分析方法,以宽度为1 m的环带替代圆形尺度分析,消除了在RipleyK(d)函数以圆形1 m步长为尺度的研究分析所带来的大小尺度的累计效应[29]。因此,本研究中采用单变量的O-ring函数[O(r)]分析不同生长阶段个体空间分布格局,用双变量的O-ring函数[O12(r)]分析两个阶段间的空间关联性。
零假设模型的筛选是保证数据分析正确与否的关键[5],因此在数据预处理阶段应仔细筛选。在进行单变量O-ring分析时,根据蒙特卡罗方法求得各生长阶段空间分布的置信区间,如果函数值处在置信区间,则接受零假设,空间分布为随机分布;如果在置信区间以上,则为聚集分布;如果函数值在置信区间以下,则为均匀分布。在群落的发育过程中,高龄个体先于低龄个体出现,而低龄个体空间的分布受高龄个体分布的影响。假设种群在发育过程中高龄个体对低龄个体有一定的庇护作用[30],采用先决条件零假设模型进行双变量O-ring空间关联分析。根据蒙特卡罗模拟结果,如果函数值在置信区间之间,表明接受零假设,二者之间相互独立;如果在置信区间以上,则二者空间上正关联;如果在置信区间以下,则二者空间上负关联。
数据处理过程主要通过Programita进行,而不同生长阶段的点格局分布则在GS+ 9.0软件中实现。采用空间尺度为0—50 m,根据相应的零假设检验模型,进行99次MentoKalo(蒙特卡罗)模拟,得到上下包迹线构成的99%置信区间[19,31- 34]。
3结果与分析
3.1辽东栎种群特征及生长阶段划分
经调查(图2),两块样地中辽东栎各径级个体树木统计如图2显示,在样地Ⅰ、Ⅱ中,辽东栎在幼龄、中龄、近熟龄和成过熟龄阶段个体数目分别为:349、161、19、20 株; 241、242、139、46 株。由生长趋势分布可以看出,样地Ⅰ内辽东栎种群径级结构分布更加接近于倒“J”型,以小径级的幼龄树居多,更新状态良好;样地Ⅱ中幼龄林木略少于样地Ⅰ,且从其空间分布状况对比可知(图1),样地Ⅰ中幼树簇生现象更加明显。
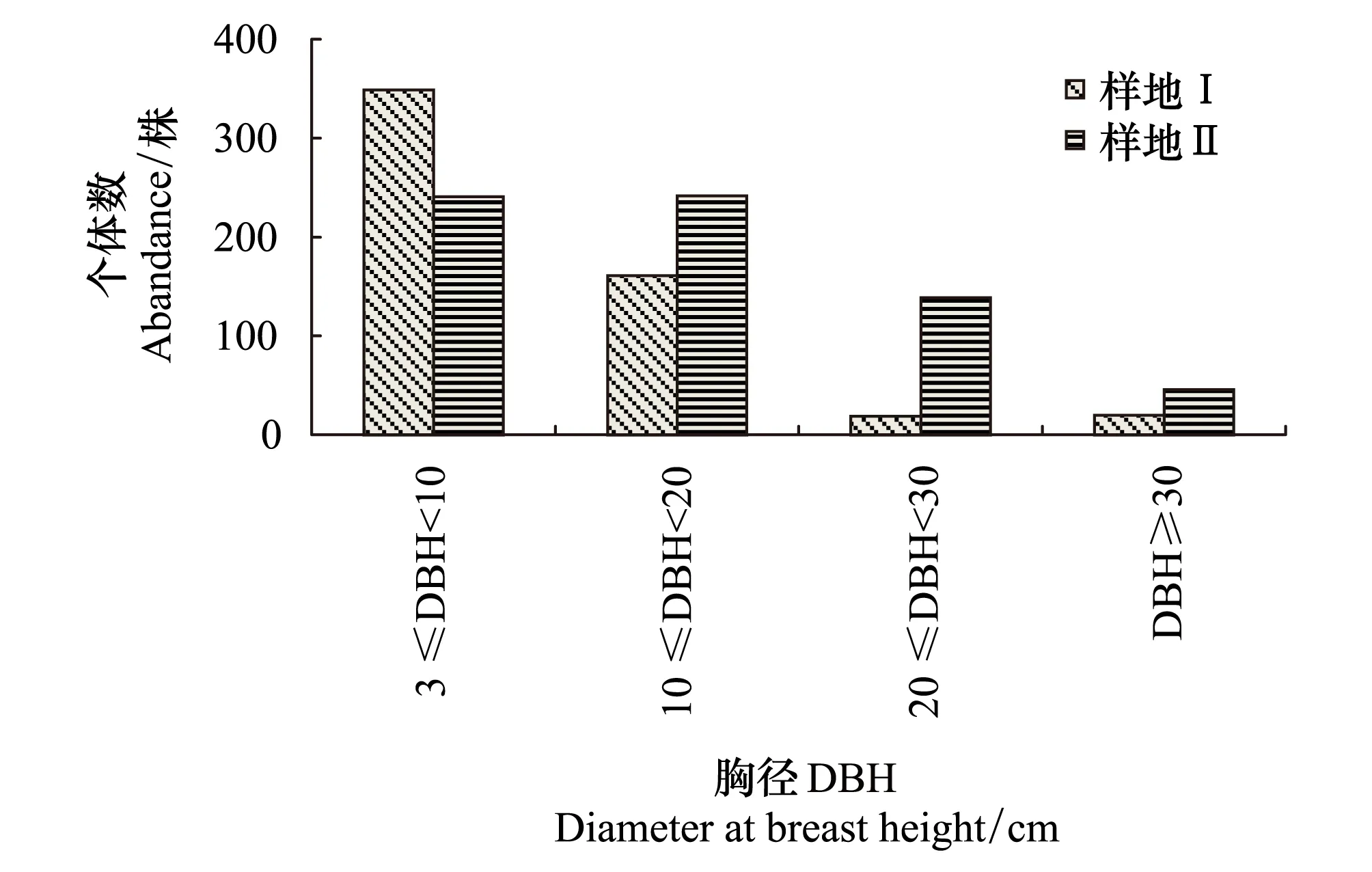
图2 样地Ⅰ和样地Ⅱ辽东栎径级分布图 Fig.2 Distribution of growth stages of Quercus wutaishanica in PlotⅠ and Polt Ⅱ
样地Ⅰ内林木达到检尺胸径3 cm的活立木2432 株,其中,油松808 株,辽东栎549 株,白桦481 株。样地内乔木平均胸径为9.86 cm,胸高总断面积为18.57 m2/hm2。辽东栎的平均胸径为10.22 cm,平均树高为8.21 m,胸高断面积为4.50 m2/hm2。样地Ⅱ内林木达到检尺胸径3 cm的活立木1018 株,其中辽东栎有668 株。样地内乔木平均胸径为12.98 cm,胸高总断面积为13.47 m2/hm2,辽东栎的平均胸径为15.36 cm,平均树高为8.03 m,胸高断面积为12.38 m2/hm2。通过对比我们可以得出,辽东栎的平均胸径在样地Ⅰ要小于样地Ⅱ,而树高正好相反,样地Ⅰ内辽东栎要比样地Ⅱ内辽东栎更高。分析其原因:在相似立地条件下,样地Ⅰ内林木株数密度要大于样地Ⅱ,样地Ⅰ内林木竞争环境要比样地Ⅱ中苛刻,对其林木生长和分布也产生一定影响。且样地Ⅰ、Ⅱ内郁闭度分别为0.79、0.48,在相同光照条件下,下层林木对光照的获得量差异较为明显;在竞争过程中,林木个体向高处生长以获得更多的光资源。而在样地Ⅱ中在光照充足的情况下这种竞争相对不明显,且林木发育状况比样地Ⅰ中要好。因此,混交林中辽东栎与竞争相对较弱的辽东栎纯林相比,树木表现出高而细的特点。
3.2辽东栎种群空间分布格局
根据调查结果(图1)明显可以看出:样地Ⅰ幼龄树明显多于样地Ⅱ,且有多处为萌生丛生分布,十分符合次生林的特点。通过对两块样地进行单变量O-ring统计分析发现(图3),随尺度变化,其空间分布格局具有不同的特点:样地Ⅰ内辽东栎在尺度6 m以内随着研究尺度的增大聚集强度逐渐减弱,在11 m、16 m、35—37 m处表现为轻度聚集,其他尺度则均以随机分布为主。样地Ⅱ内辽东栎主要表现为随机分布,仅在1—1.8 m之间,3—5.7 m之间,7—9 m之间表现为聚集分布。
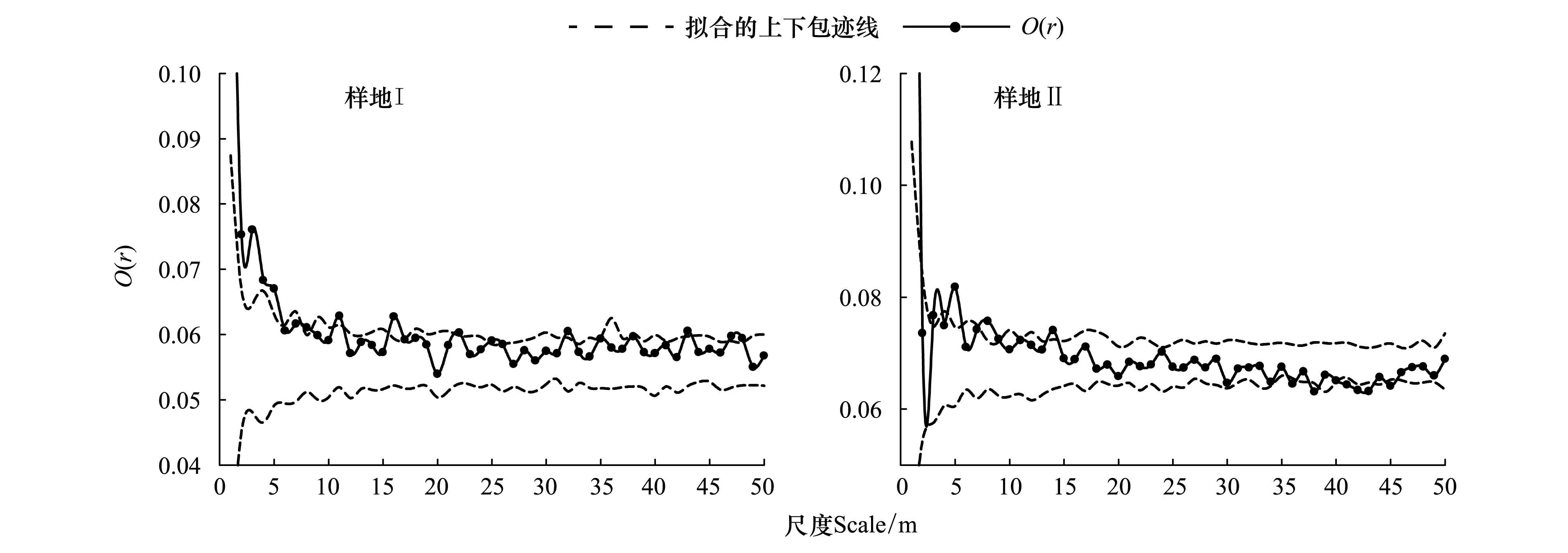
图3 样地Ⅰ和样地Ⅱ空间分布格局Fig.3 Spatial pattern of Quercus wutaishanica in PlotⅠ and Polt Ⅱ
3.3辽东栎种群不同龄级的空间分布格局
两个样地的辽东栎在4 个不同的生长阶段的点格局分析表明(图4,图5),两个样地中辽东栎胸径小于20 cm的幼龄和中龄个体空间分布稍有不同:样地Ⅰ中,辽东栎个体基本表现为随机分布;样地Ⅱ中胸径位于3—10 cm之间的个体在11 m以内为聚集分布,11 m以外为随机分布,10—20 cm之间的个体在2 m以内及3—4 m、5.4—7 m之间表现为聚集分布,其他为随机分布。在胸径大于20 cm的近熟和成过熟阶段个体,均表现为随机分布,并有呈均匀分布的趋势。
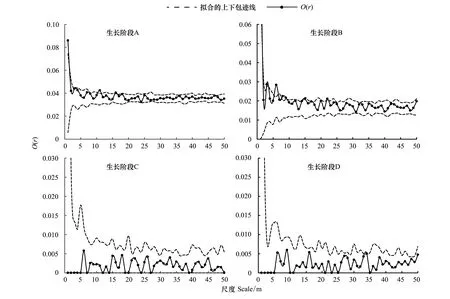
图4 样地Ⅰ各龄级空间分布格局Fig.4 Spatial pattern of each stage of Quercus wutaishanica in plotⅠA: 幼龄阶段A; B: 中龄阶段B; C: 近熟龄阶段C; D: 成过熟龄D

图5 样地Ⅱ各龄级空间分布格局Fig.5 Spatial pattern of each stage of Quercus wutaishanica in plot ⅡA: 幼龄阶段A; B: 中龄阶段B; C: 近熟龄阶段C; D: 成过熟龄D
3.4辽东栎种群不同龄级间的空间关联性
通过对两块样地进行双变量分析结果显示(图6,图7),样地Ⅰ中辽东栎各生长阶段之间关联性均不明显,而样地Ⅱ中各生长阶段之间表现不同:样地Ⅱ中幼龄个体与中龄个体间在小尺度上表现出显著的正相关性,而与近熟龄个体和成过熟龄个体在小尺度上表现为明显的负相关性。此外,在样地Ⅱ中2—5 m尺度上,中龄个体与成过熟龄个体也表现出一定的负相关性。

图6 样地Ⅰ各龄级间关联性Fig.6 Associations of different growth stages of Quercus wutaishanica in PlotⅠA: 幼龄阶段A; B: 中龄阶段B; C: 近熟龄阶段C; D: 成过熟龄D

图7 样地Ⅱ各龄级间关联性Fig.7 Associations of different growth stages of Quercus wutaishanica in Plot ⅡA: 幼龄阶段A; B: 中龄阶段B; C: 近熟龄阶段C; D: 成过熟龄D
4结论与讨论
辽东栎是黄龙山林区植被中重要的建群种之一。在自然环境条件下,不同生长阶段的种群处于不同的林层,受到周围环境的影响程度不尽相同,因此其表现出的分布格局也大不相同[35]。根据林木生长规律[36]可知:生长条件相似、年龄阶段相同的同种个体之间存在激烈的种内竞争,包括光照、养分等;生长条件类似、生长阶段不同的同种个体,由于竞争原因,高龄个体会对低龄个体产生一定的影响[30]。因此,本文根据辽东栎静态生命表将辽东栎划分为4个不同生长阶段,在研究各生长阶段空间分布格局的同时,分析其不同生长阶段的相互关联性,对了解该物种生长状况有着重要意义[37]。
(1)辽东栎的空间分布格局与尺度有密切关系:在2块典型的天然次生林样地中,辽东栎在小尺度上均表现为聚集分布,随尺度增加,其聚集特点逐渐减弱,在中、大尺度上均表现为随机分布。两块样地中辽东栎在小尺度上表现稍有不同:样地Ⅰ中,辽东栎个体在0—11 m以内为明显的聚集分布并随尺度的增大逐渐减弱;而样地Ⅱ中,在2—6 m尺度上,辽东栎表现为聚集分布,且随尺度变化特征不明显。
(2)不同样地相同生长阶段其空间分布特征不同:样地Ⅰ的分布主要表现为随机分布,而在样地Ⅱ中,幼、中龄个体在小尺度上表现为聚集分布,样地Ⅱ中幼树发育要比样地Ⅰ中有环境优势;在同一样地不同生长阶段林木的相互关联分析发现,样地Ⅰ中,低龄个体大多分布于高龄阶段树木树冠和根系范围之外,高龄阶段树木对低龄树木发育影响程度不显著,且从图6中各不同生长阶段间相互关系可以看出,各阶段间的相关性基本表现为不明显。在样地Ⅱ中,成、过熟个体对中幼龄个体生长具有负作用。这与伊力塔[27]、胡秀娟[30]等关于辽东栎整体空间分布规律具有一致性。
(3)相同样地不同生长阶段的关联性也不尽相同:样地Ⅰ中各阶段之间并没有太大的联系,而样地Ⅱ中,在小尺度上,幼、中龄个体分别与近、成过熟个体有着一定的负相关。从图1分析可以得出,样地Ⅱ中高龄阶段个体和低龄个体的分布重合较少,因此可以推断,在高龄个体生长的树冠和根系覆盖范围内,幼龄个体因受空间、养分、阳光等环境因子的限制而存活率相对较低,处于高龄个体树冠及根系覆盖范围外的低龄个体,由于空间、养分、阳光等环境条件较优,其发育空间竞争性较低,其个体的发育状况相对要好,从而表现为负相关性;而在样地Ⅰ中,由于郁闭度较高(0.79),物种之间的激烈竞争导致幼树更新地点的选择有很大的局限性。样地Ⅰ、Ⅱ中的林分郁闭度分别为0.79、0.48,通过比较可以得出,样地Ⅰ内上层木对阳光的遮挡作用对下层更新影响很大,样地Ⅱ的条件相对要比样地Ⅰ更有利于林下更新。针对这种情况,在进行森林抚育的时,可以对样地Ⅰ中上层木进行适当疏伐,以保证林下植物的生长及林分更新。
对辽东栎次生林而言,种群成簇萌生是影响其空间分布的重要因素[38],但是环境的异质性是物种后期发展的主导因素。种群在环境影响及其自身生长特性的共同影响下,形成本群落生存发展的有利结构与其他物种竞争环境资源,确保自身的正常发展[18]。研究结果表明,在两块样地中,低龄个体树木株数比高龄个体树木株数整体要多,其径级结构基本呈现倒“J”型,两块样地中辽东栎种群径级结构分布较为合理[25]。样地Ⅰ中乔木株数密度为2432 株/ hm2,样地Ⅱ中乔木株数密度为1018 株/ hm2,在相似立地条件下,样地Ⅰ竞争环境要比样地Ⅱ中苛刻,对其林木生长和分布也产生一定影响。
种群的空间格局对尺度有着很大的依赖性[27],尺度不同,其表现出来的特征也不尽相同。本研究采用公顷级样地对黄龙山林区辽东栎空间分布格局开展研究。对于辽东栎次生林而言,在较小尺度上,主要由其种群起源、萌生以及种子的传播等生物特性决定。而随着尺度的增大,则由地形地貌、光照、水分等自然条件产生的影响逐渐取代物种自身产生的影响,从而随自然条件形成不同空间分布格局[19]。研究不同生长阶段的空间关联性有利于深入了解其群落的发展趋势[39],并对其空间格局的形成过程有一定的了解。而研究种群分布的空间格局有助于充分了解林分的空间分布格局[17],并有针对性的对种群聚集性较大的区域开展结构化森林经营。
研究结果表明(图1),黄龙山林区辽东栎纯林和油松-辽东栎-白桦林中,松栎桦混交林种群结构相对稳定,其径级结构要比辽东栎纯林中更加符合倒“J”型特点。按照群落演替的一般规律,通过一段时间的竞争、淘汰,辽东栎幼龄个体在发育过程中逐渐完成自然稀疏,在此过程中,进行适当的人工干扰如疏伐、抚育等既可以加速该过程,又能改善其内部结构,使其更加有利于群落的发育。群落的形成影响因素有许多[4,33],不是简单的空间格局分析就能完成的。因此,辽东栎与其他物种(如油松、白桦等)、物种与环境因子(如土壤、坡度等)之间的相互关系及影响还有待进一步研究,以期更深入地认识辽东栎空间格局形成的起因与过程,更好地为当地森林经营提供依据。
参考文献(References):
[1]方坚, 王孝安, 郭华, 朱志红. 黄土高原马栏林区辽东栎林种内、种间竞争研究. 西北植物学报, 2007, 27(2): 334- 339.
[2]Nakashizuka T. Species coexistence in temperate, mixed deciduous forests. Trends in Ecology & Evolution, 2001, 16(4): 205- 210.
[3]Moloney K A, Wiegand T. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209- 229.
[4]白小军. 大兴安岭典型林区主要植物点格局分布研究. 现代农业科技, 2012, (9): 194- 196, 198- 198.
[5]赵中华, 惠刚盈, 胡艳波, 周红敏, 张连金. 2种类型阔叶红松林优势种群空间分布格局及其关联性. 林业科学研究, 2011, 24(5): 554- 562.
[6]Schurr F M, Bossdorf O, Milton S J, Schumacher J. Spatial pattern formation in semi-arid shrubland: a priori predicted versus observed pattern characteristics. Plant Ecology, 2004, 173(2): 271- 282.
[7]杨晓东, 吕光辉, 张雪梅, 孙丽君, 何书林. 艾比湖湿地自然保护区8个乔灌木种群空间分布格局分析. 植物资源与环境学报, 2010, 19(4): 37- 42.
[8]任珩, 赵成章. 高寒退化草地狼毒与赖草种群空间格局及竞争关系. 生态学报, 2013, 33(2): 435- 442.
[9]缪宁, 刘世荣, 史作民, 喻泓, 刘兴良. 川西亚高山红桦-岷江冷杉林优势种群的空间格局分析. 应用生态学报, 2009, 20(6): 1263- 1270.
[10]侯继华, 黄建辉, 马克平. 东灵山辽东栎林主要树种种群11年动态变化. 植物生态学报, 2004, 28(5): 609- 615.
[11]康永祥, 岳军伟, 雷瑞德, 梁宗锁, 陈永剑. 陕北黄龙山辽东栎群落优势种群生态位研究. 西北植物学报, 2008, 28(3): 574- 581.
[12]康永祥, 岳军伟, 张巧明. 黄龙山林区辽东栎群落类型划分及其生物多样性研究. 西北林学院学报, 2007, 22(3): 7- 10.
[13]张晓辉, 周建云, 何景峰, 卢彦昌, 张文辉. 黄龙山林区主要森林群落下物种多样性研究. 西北林学院学报, 2007, 22(4): 39- 44.
[14]郭其强, 张文辉, 曹旭平. 基于模糊综合评判的森林群落稳定性评价体系模型构建——以黄龙山主要森林群落为例. 林业科学, 2009, 45(10): 19- 24.
[15]冯云, 马克明, 张育新, 祁建. 北京东灵山地区辽东栎(Quercusliaotungensis)种群生活史特征与空间分布. 生态学杂志, 2009, 28(8): 1443- 1448.
[16]张育新, 马克明, 祁建, 冯云, 张洁瑜. 北京东灵山海拔梯度上辽东栎种群结构和空间分布. 生态学报, 2009, 29(6): 2789- 2796.
[17]袁志良, 王婷, 朱学灵, 沙迎迎, 叶永忠. 宝天曼落叶阔叶林样地栓皮栎种群空间格局. 生物多样性, 2011, 19(2): 224- 231.
[18]常新华, 赵秀海, 曾凡勇, 张春雨. 长白山针阔混交林主要树种空间分布及其环境解释. 北京林业大学学报, 2009, 31(1): 7- 12.
[19]杜志, 亢新刚, 孟京辉, 孔雷, 郭韦韦, 岳刚. 长白山杨桦次生林主要树种的空间分布格局及其关联性. 东北林业大学学报, 2013, 22(4): 36- 42.
[20]焦文婧, 郭东罡, 张婕, 上官铁梁. 灵空山自然保护区油松-辽东栎林建群种关联性. 生态学杂志, 2012, 31(12): 3050- 3057.
[21]岳永杰, 余新晓, 刘彦, 樊登星, 史宇, 王小平, 陈俊崎. 华北落叶松林不同发育阶段种群分布格局研究. 北京林业大学学报, 2008, 31(Z2): 171- 176.
[22]田丽, 王孝安. 黄土高原马栏林区辽东栎种群动态分析. 江西农业学报, 2011, 23(2): 39- 42.
[23]曹旭平, 郭其强, 张文辉. 黄龙山油松林和油松+辽东栎混交林物种组成及优势种群动态. 西北植物学报, 2010, 30(5): 1012- 1019.
[24]张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344- 349.
[25]霍萌萌, 郭东罡, 张婕, 王治明, 上官铁梁, 李润强, 刘卫华. 灵空山油松-辽东栎林乔木树种群落学特征及空间分布格局研究. 生态学报, 2014, 34(20): 5925- 5935.
[26]王巍, 刘灿然, 马克平, 于顺利. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 1999, 41(4): 425- 432.
[27]伊力塔, 韩海荣, 程小琴, 康峰峰, 张志杰. 灵空山林区辽东栎(Quercusliaotungensis)种群空间分布格局. 生态学报, 2008, 28(7): 3254- 3261.
[28]孟宪宇. 测树学. 北京: 中国林业出版社, 2006: 63.
[29]牛丽丽, 余新晓, 刘淑燕, 刘彦, 岳永杰, 王小平, 陈俊崎. 北京松山自然保护区油松种群生活史及其空间分布格局. 北京林业大学学报, 2008, 30(Z2): 17- 21.
[30]胡秀娟, 程积民, 杨晓梅, 范文娟, 孟蕾, 韩娟娟. 黄土高原子午岭林区辽东栎种群分布格局及群落特征研究. 西北林学院学报, 2010, 25(5): 1- 6.
[31]杜志, 亢新刚, 包昱君, 杨鑫霞. 长白山云冷杉林不同演替阶段的树种空间分布格局及其关联性. 北京林业大学学报, 2012, 34(2): 14- 19.
[32]张莉, 张小平, 陆畅, 李晓红. 安徽琅琊山青檀种群空间格局. 林业科学, 2012, 48(2): 9- 15.
[33]郭垚鑫, 康冰, 李刚, 王得祥, 杨改河, 王大伟. 小陇山红桦次生林物种组成与立木的点格局分析. 应用生态学报, 2011, 22(10): 2574- 2580.
[34]马莘, 夏颖, 马庆庆, 宋会兴. 四川牡丹群落物种生态位及空间关联度分析. 重庆师范大学学报: 自然科学版, 2011, 28(4): 57- 61.
[35]伊力塔, 豪树奇, 徐涌, 康峰峰. 辽东栎群落特征研究及其进展. 辽宁林业科技, 2010, (5): 25- 30, 39- 39.
[36]龚直文, 亢新刚, 顾丽, 杨华. 长白山杨桦次生林生长过程与演替动向分析. 林业科学研究, 2009, 22(3): 74- 79.
[37]宋于洋, 李园园, 张文辉. 梭梭种群不同发育阶段的空间格局与关联性分析. 生态学报, 2010, 30(16): 4317- 4327.
[38]孙书存, 陈灵芝. 辽东栎植冠的构型分析. 植物生态学报, 1999, 23(5): 433- 440.
[39]龚直文, 顾丽, 亢新刚, 杨华, 赵浩彦. 长白山森林次生演替过程中林木空间格局研究. 北京林业大学学报, 2010, 32(2): 92- 99.
Spatial pattern ofQuercuswutaishanicain natural secondary forest of Huanglong Mountain
WANG Daoliang1, LI Weizhong1,*, CAO Zhu1, WANG Qing1, WANG Guangru2
1CollegeofForestry,NorthwestA&FUniversity,Yangling712100,China2ForestBureauofHuanglongshaninYan′anCity,Huanglong715700,China
Abstract:Spatial pattern analysis is an important method for studying population characteristics, interspecies relationships, as well as the relationships between population and environment. In the present paper, population characteristics of Quercus wutaishanica in natural secondary forest of Huanglong Mountain were investigated. One natural secondary forest plot (with area of 1 hm2) was selected and set up in each Q. wutaishanica-Pinus tabuliformis-Betula platyphylla natural secondary mixed forest and Q. wutaishanica natural secondary pure forest. The data and coordinates of each tree in the two plots were measured and recorded following the standards for plot investigation. Using the method of diameter class (instead of the age), we converted the classification of different growth stages of Q. wutaishanica into size classes for diameter at breast height (DBH) according to the static life table. Thus, Q. wutaishanica was divided into four different growth stages: the immature stage A, 3 cm ≤ DBH < 10 cm; mid-mature stage B, 10 cm ≤ DBH < 20 cm; near-mature stage C, 20 cm ≤ DBH < 30cm; mature and over-mature stage D, DBH ≥ 30 cm. The spatial pattern of Q. wutaishanica was analyzed through O-ring statistical means for each growth stage by using single variable method and the interconnectedness between the growth stages by bivariate-analysis method. The results showed the following: (1) The diameter structure of Q. wutaishanica in the two plots was in line with inverted "J" type, indicating that the natural regeneration in the two plots is better from the perspective of diameter class distribution. (2) There was a close relationship between the spatial pattern and the scale of Q. wutaishanica. It showed random distribution in the two selected plots at a large and medium scale and aggregated distribution at a small scale, especially at the scale of 0—6 m in plot I. (3) The spatial patterns differed at different growth stages, young trees exhibited aggregated distribution at a small scale, especially at the scale of 0—11 m in plot II. (4) The correlation between different growth stages was not identical in the plots. There was no significant correlation between different growth stages within plot I. The immature and mid-mature individuals showed a significant positive correlation at a small scale of plot II, whereas the near-mature, mature, and over-mature individuals showed a significant negative correlation at a small scale. The mid-mature, mature, and over-mature individuals were negatively correlated at the 2—5 m scale. Q. wutaishanica stand in the natural secondary forest on Huanglong Mountain was one of the important constituent species. The study of spatial distribution at different growth stages of individual organisms is critical to capture the growth situation of Q. wutaishanica in Huanglong Mountain. Spatial correlation of different growth stages is vital to understand the development trend of the community as well as to provide an important basis for the development of structured forest management for greater population density.
Key Words:natural secondary forest; spatial distribution pattern; spatial correlation; O-ring statistical analysis; Quercus wutaishanica
基金项目:国家自然科学基金项目(31170587)
收稿日期:2014- 06- 14; 网络出版日期:2015- 08- 26
*通讯作者
Corresponding author.E-mail: wzhli6465@163.com
DOI:10.5846/stxb201406141233
王道亮,李卫忠,曹铸,王庆,王广儒.黄龙山天然次生林辽东栎种群空间格局.生态学报,2016,36(9):2677- 2685.
Wang D L, Li W Z, Cao Z, Wang Q, Wang G R.Spatial pattern ofQuercuswutaishanicain natural secondary forest of Huanglong Mountain.Acta Ecologica Sinica,2016,36(9):2677- 2685.
