长白山原始林红松径向生长及林分碳汇潜力
2016-07-21罗春旺徐倩倩孟盛旺李俊清刘琪璟
于 健,罗春旺,徐倩倩,孟盛旺,李俊清,刘琪璟,*
1 北京林业大学, 林学院省部共建森林培育与保护教育部重点实验室, 北京 100083 2 江苏农林职业技术学院, 句容 212400 3 中国农业科学院郑州果树研究所, 郑州 450009
长白山原始林红松径向生长及林分碳汇潜力
于健1,2,罗春旺1,徐倩倩3,孟盛旺1,李俊清1,刘琪璟1,*
1 北京林业大学, 林学院省部共建森林培育与保护教育部重点实验室, 北京100083 2 江苏农林职业技术学院, 句容212400 3 中国农业科学院郑州果树研究所, 郑州450009
摘要:长白山地处温带针阔混交林的中心地带,其植物生长对气候变化反映较为敏感。运用树木年代学的基本原理与方法,以该区保存完好的地带性顶级群落阔叶红松林为研究对象,探讨了红松 (Pinus koraiensis)径向生长对气候变化的响应,以期揭示全球气候变化对阔叶红松林及红松树木生长的影响。以长白山自然保护区北坡为研究地点,建立红松树轮年表,计算年均生物量生长量,与逐月各气候因子进行相关和响应函数分析,以期揭示红松径向生长与气候变化的关系。同时利用多元逐步回归分析方法分别建立了红松树轮指数、年均生物量生长量与主要气候因子的模拟回归方程。主要结论如下:(1)长白山红松年表特征分析表明其生长对气候变化敏感,适用于进行树轮气候学分析;(2)红松径向生长受当年生长季前期温度影响较大;(3)温度是限制红松径向生长的主要因子,气候变暖将促进红松的径向生长;(4)目前林分单位面积地上生物量为310.88t/hm2,单位面积林分生物量生长量约为5.077t hm-2a-1,即每公顷每年固碳约2.539t,表明该地区老龄阔叶红松林仍具有较强的固碳能力。
关键词:树轮宽度;气候变暖;红松;年均生物量生长量;碳汇潜力
过去130a全球平均温度已升高0.85℃,人类活动极有可能是20世纪中期以来全球气候变暖的主要原因,可能性在95%以上[1]。北半球高纬度较低纬度地区增温显著[2]。比如,我国北方尤其是东北地区气温增加态势表现得尤为突出,气温增加幅度大于西北和华北地区,并表现出明显的干旱化趋势[3- 4]。这意味着分布在北方高纬度地区的森林生态系统更易受外界因素如气候变化的影响[1,5]。长白山具有中国保持最为完整的原始森林,是温带针阔叶混交林生态系统的重要组成部分。长白山地处我国北方中高纬度,属于典型的温带大陆性季风气候区,是我国气候变化最剧烈的地区之一[6]。该区的阔叶红松林为中国东北东部山地温带针阔叶混交林的地带性顶极群落,具有丰富的物种多样性和较大的生产力,原始阔叶红松林目前已接近森林演潜的顶极阶段[7]。红松(Pinuskoraiensis)是长白山阔叶红松林主要建群树种,其垂直分布在海拔500—1300m,个体分布可达1400m[8]。探讨其径向生长与气候因子的关系,对了解全球气候变化背景下北方森林的响应,尤其是长白山森林的动态具有重要意义。
树木年轮资料由于定年准确、分辨率高、样本分布广泛、易于获取、时间序列长、并记录了树木生长的丰富信息等特点,近年来已被广泛应用于评估过去气候变化[9]。前人在该区围绕建群种红松开展了较多的树木年轮气候学的研究[10- 11],主要涉及在不同海拔[12- 13]、不同林型[14]、不同年龄[15]、不同树高[16]以及火干扰[10,17]等条件下红松径向生长对气候因子的响应关系。该区高海拔红松径向生长不仅受降水限制,还对气温存在“滞后响应”,低海拔红松生长主要受气温限制[13]。但是更多研究得出的结论是:高海拔地区或者近林线处树木径向生长主要受夏季和生长季温度控制,而低海拔地区主要受降水因子影响[18]。共同的结论是,区域气候的差异会造成不同海拔树木径向生长对气候响应的多样性[19]。尽管前人对低海拔红松与气候关系的研究取得了一定的成果,但针对森林下限红松生长与气候因子关系的研究结果差异较大。已有的研究往往仅从树轮宽度指数与气候因子的相关性来探讨气候变化对该区树木径向生长的影响,而极少用到树木生物量生长量这个三维尺度的树木生长指标来探讨对气候变化的响应[20]。综上所述,需要扩充采样范围,在不同地区开展树木径向生长与气候变化响应机制的研究,有助于评估和预测未来气候变化下森林生态系统动态。
为了明确该区红松径向生长的气候响应机制,探讨该区阔叶红松林森林固碳能力,本研究采用树木年代学方法建立了北坡红松树轮宽度年表,并计算红松年均生物量生长量;将这两个生长指标与逐月各气候因子进行相关分析,分别建立其与气候因子的多元逐步回归方程。本文还依据测定的年轮宽度信息,结合样地调查数据,分析了长白山北坡红松树轮径向生长和年均生物量生长量的生长过程,估算了阔叶红松林地上总生物量和年生物量增量,评估了该区老龄林的固碳能力。
1试验地概况及研究方法
1.1试验地概况与样地调查
研究地点位于长白山北坡阔叶红松林带,优势树种为红松(Pinuskoraiensis),其它乔木树种主要有紫椴 (Tiliaamurensis)、蒙古栎(Quercusmongolica)、水曲柳(Fraxinusmandschurica) 、核桃楸 (Juglansmandshurica)、色木槭(Acermono)、黄菠萝(Phellodendronamurense)等,下木种主要有毛榛 (Corylusmandshurica)、卫矛(Evonymusalatus)等。研究区气候特征为受季风影响的温带大陆性季风气候,夏季温暖多雨,冬季寒冷漫长,温度与降水四季变化明显。年均气温约2.8℃,平均年降水量600—900mm,降水多集中在夏季,6—9月份降水量占全年降水量的80%以上,年相对湿度在70%左右 (图1)。
2010年8月在长白山北坡开展调查,地点名称大阳岔(海拔838m,42°21′03″N;127°59′16″E),是平坦的火山台地,坡度﹤5°。设立了25个半径为8 m的样圆 (总面积约为0.5 hm2)进行调查,对样地内所有林木 (胸径≥10cm) 进行每木检尺,记录乔木层全部树木的树种名称、胸径和树皮厚度。林分郁闭度为0.9左右,林木株数密度为362株/hm2。
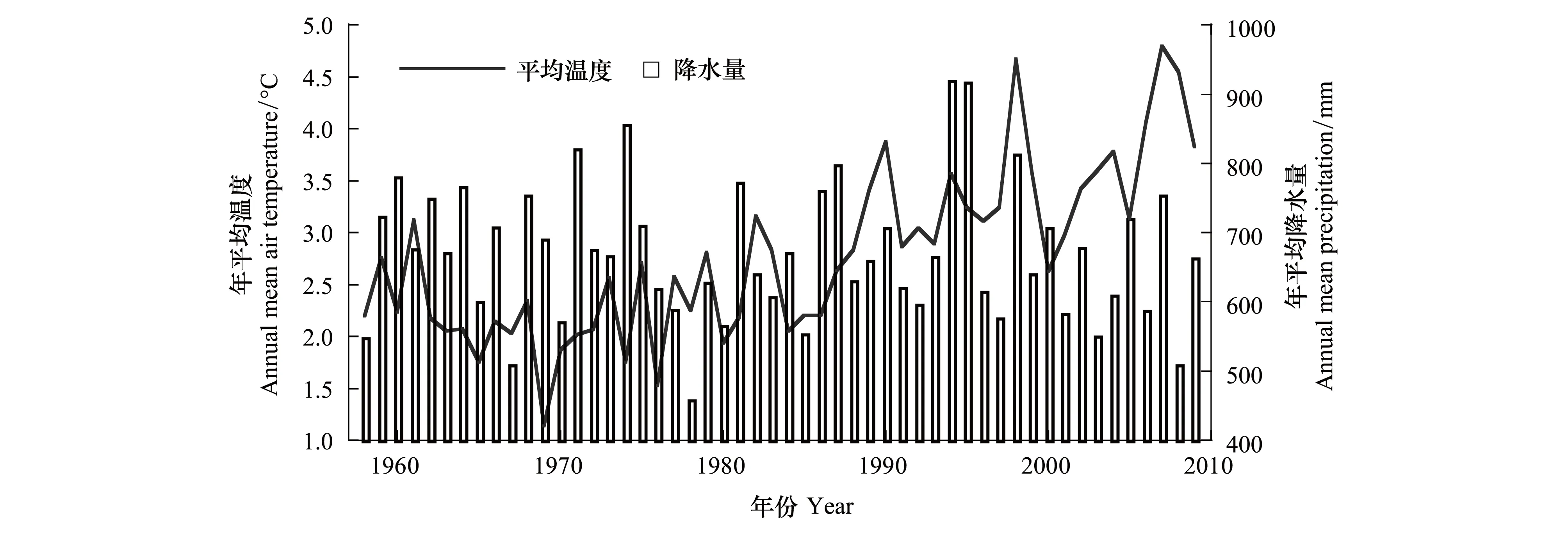
图1 研究区域温度和降水量的年变化Fig.1 Annual variations of temperature and precipitation in the study area
1.2气象资料
选取距离研究地点最近的气象站——松江气象站(地理位置: 42°32′N, 128°15′E, Height: 591.4m),位于样地东北31.03km。资料时间跨度为1957至2009年。气象要素包括月平均气温(Tm)、月降水量(Pm)、月平均最高气温(Tmax)和月平均最低气温(Tmin)。气候资料来源于中国气象科学数据共享网 (http://cdc.cma.gov.cn/home.do)。检查气象资料的均一性采用距离松江气象站较近的东岗气象站作为参考站。采用Mann-Kendall方法[21]对气温数据进行均一性检验与校正,利用Double-mass方法[22]对气象站降水资料进行均一性检验与校正。经过检验确认气象数据可靠,气温和降水数据资料变化相对均一,该站气象数据可用来代表气候的基本特征。本研究还选取了时间跨度为1958—2005年的旧版帕尔默干旱指数(PDSI)作为气候分析的要素之一。PDSI是反映干湿变化的指数之一,综合了降水和蒸发的影响,是水分亏缺量与持续时间的函数,比降水量更能解释树木在生长期的水分供应平衡[23]。PDSI值越高表明水分供应情况越良好,反之越低则表明某一时间越干旱。本研究所用PDSI数据是由Dai提供的2.5个经纬度网格点数据[24],选用距离采样点周围最近的4个格点(42°15′N/127°45′E、42°15′N/128°15′E、42°45′N/127°45′E、42°45′N/128°15′E)平均后的数据用于分析。
1.3树轮资料获取和年表建立
遵循树木年代学方法的基本要求,于2010 年6月在样地附近,利用生长锥在树木胸径位置钻取完整树轮样芯(尽量到髓心)用于分析生长过程及其同气候的关系,共钻取了82根大径级红松树轮样芯。为了减少对树木的伤害,每株树钻取一根树轮样芯。另外,为了分析林木近期生长速度,在所设置的样圆内采用全样地钻取浅层样芯,钻取深度5cm以内,共取样181株。样芯自然风干后,采用常规方法进行处理[25]。使用LINTAB 6.0年轮仪 (精度为0.01mm)测量年轮宽度。对靠近髓心部分缺失的年轮(多数情况钻不到髓心)根据年轮的弓高和弦长进行近似估计[26]。
采用COFECHA程序[27]对交叉定年和测量结果进行检验。经交叉定年检验,剔除树龄较短、与主序列相关性较差和难以交叉定年的序列(共计31根),最后有51根样芯进入主序列,用于年表建立。年表的建立通过计算机程序 ARSTAN 完成[28]。以2/3的年龄作为步长的样条函数去掉树木因本身遗传特性产生的生长趋势和树木之间干扰竞争产生的抑制及释放等的生长趋势,最终得到三类年表,分别为标准年表(STD)、差值年表(RES)和自回归年表(ARS),图 2为红松差值年表。

图2 红松差值年表Fig.2 Residual chronology of Pinus koraiensis
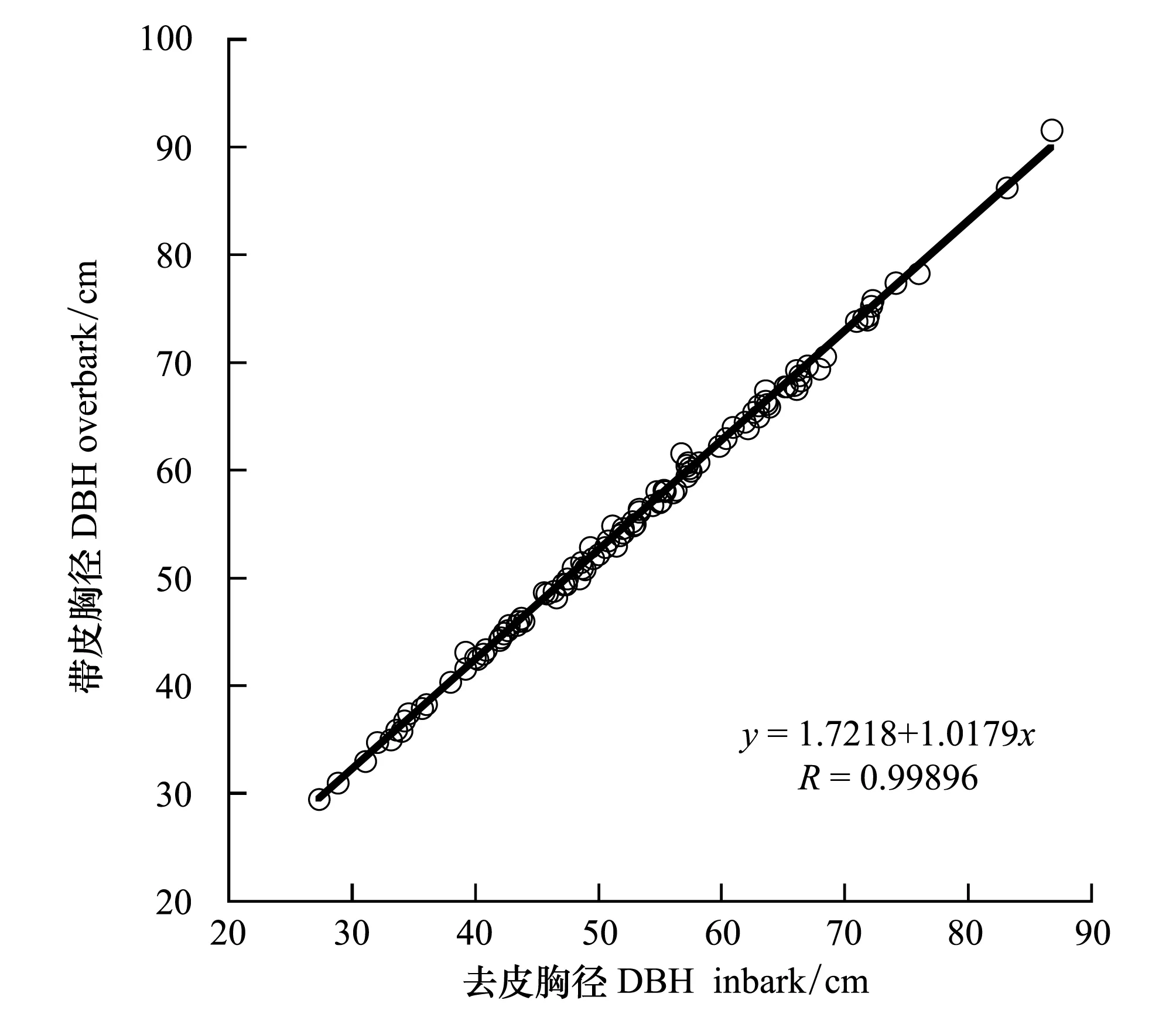
图3 红松去皮直径同带皮直径的关系 Fig.3 Relationship between over- and in- bark diameters for Pinus koraiensis
1.4生物量计算
借鉴前人[29]的研究方法对红松年均生物量生长量和林分每公顷每年生物量增量进行估算。通过当年测定的胸径D2009。由于取样时间为2010年6月,当年年轮还未完全形成,故取2009年作为最后一年,依次减去历年2个树芯年轮宽度之和wn,得到单木历年胸径Dn(式1)。应用树轮样芯和树干解析方法计算的年轮半径通常是去皮的,而在实际计算生物量时多使用带皮直径,因此需要结合样地调查数据拟合的红松带皮直径(Db)和去皮直径 (Dw)的回归方程 (式2,相关系数R=0.99),将红松历年去皮直径转换为带皮直径 (图3),以排除因忽略树皮而导致计算年生物量生长量时产生的误差。结合长白山已有的生物量回归模型[30- 32](表1),计算红松单木历年的年均生物量生长量及样地内各株地上生物量,各株地上生物量相加得到样地地上总生物量(式3),进而换算为单位面积的地上总生物量。
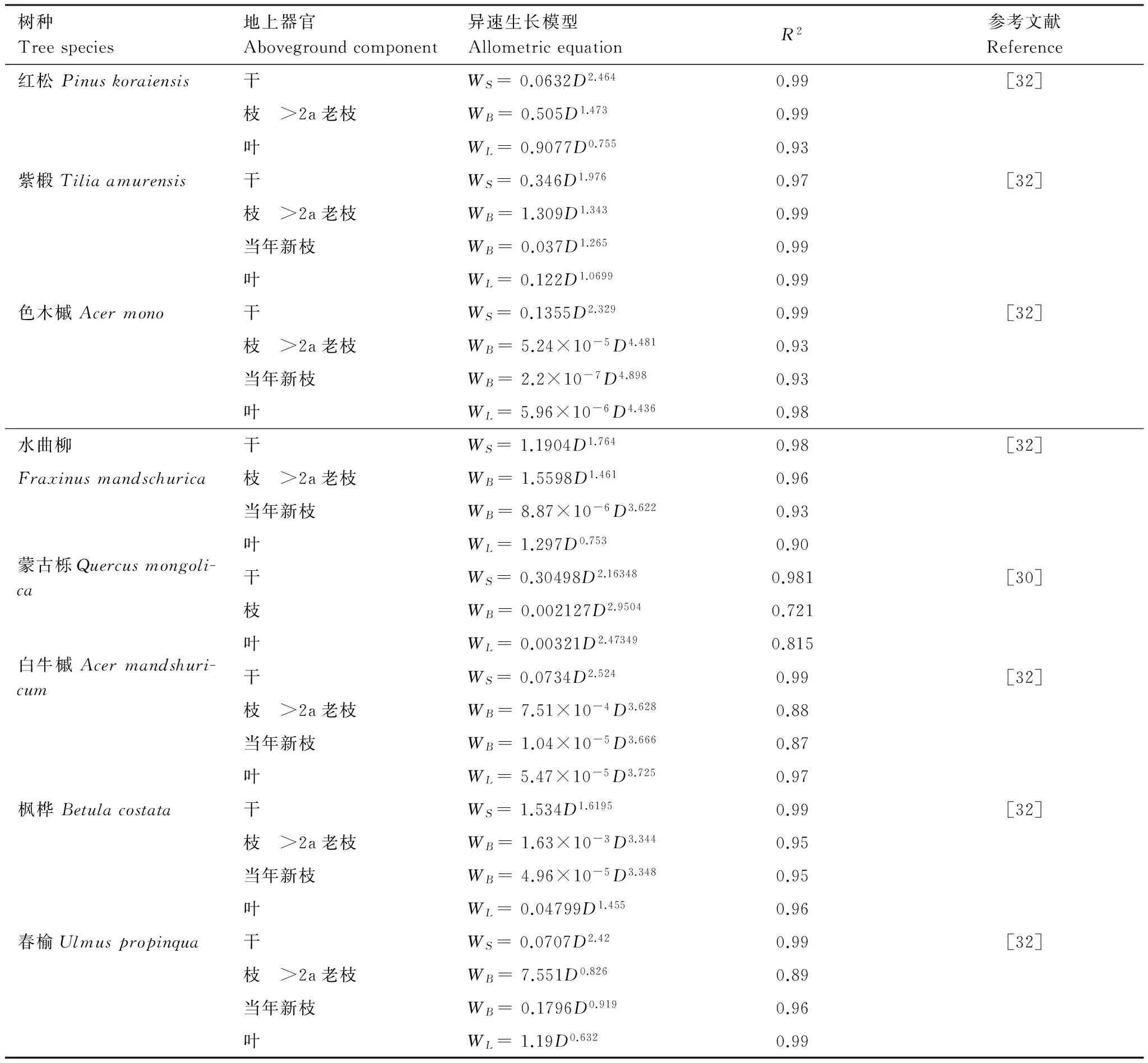
表1 长白山阔叶红松林乔木地上生物量异速生长模型
D:胸径 DBH;H:树高 Tree height;WS:干生物量 Stem biomass;WB:枝生物量Branch biomass;WL:叶生物量Leaf biomass

图4 红松生物量年表Fig.4 The Biomass chronology of Pinus koraiensis
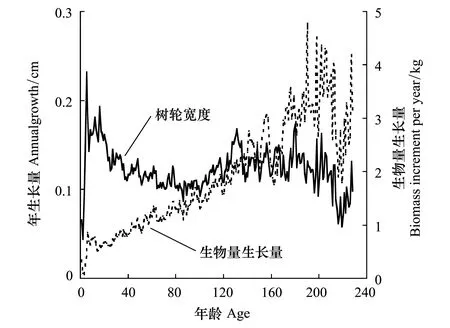
图5 红松径向生长曲线和生物量生长过程Fig.5 Growth curves of radius and biomass of Pinus koraiensis.
将51根通过交叉定年检验的红松年轮样芯利用上述方法分别计算红松年均生物量生长量。年均生物量生长量年表的建立通过计算机程序 ARSTAN 完成[28]。经过比较,最终以2/3的年龄作为步长的样条函数去掉树木本身遗传因素产生的生长趋势和树木之间干扰竞争产生的抑制和释放等的生长趋势,得到红松年均生物量生长量年表(图 4)。在51根经过交叉定年检验的红松样芯中,挑选18株树龄较长(年龄均在132a以上)且已经准确定年的红松样芯分析其树轮径向和年均生物量生长过程。经对比发现,这些年轮样芯的径向生长趋势非常一致。为了揭示红松的生长过程,将这18株红松的年轮宽度和年均生物量生长量按生理年龄对齐(即髓心对齐,这与气候响应分析时的做法不同)进行累加求平均得到径向生长量和年均生物量生长量过程曲线(图5)。
Dn=D2009-(w2009+w2008+···+wn+2+wn+1)
(1)
Db=1.7218+1.0179Dw
(2)
WT=WS+WB+WL
(3)
Bn=Wn-Wn-1
(4)
式中,Dn是n年的胸径,wn是n年的两个不同树芯年轮宽度之和。Db为红松带皮直径,Dw为红松去皮直径,H为树高。WT为地上总生物量,WS为树干生物量,WB为树枝生物量,WL为叶片生物量。Wn为n年的生物量,Bn为n年的生物量生长量。
1.5统计分析
在研究树木生长与气候因子之间的线性关系中,最常用的方法就是相关分析[33]和响应函数分析[34]。本文采用相关及响应函数分析树木径向生长与气候因子的关系,通过Dendroclim 2002程序完成[35]。由于树木生长对气候的响应存在“滞后效应”。研究选取上年5月到当年9月气候因子进行分析。利用多元逐步回归分析模拟红松树木径向生长及年均生物量生长量和逐月值气候因子的关系,选取最优方程。数据分析在SPSS 17.0软件中进行,统计的显著性水平取0.05。
2结果
2.1树轮年表的基本统计特征
年轮气候学研究表明,一般高质量的树轮年表具有信噪比高、平均敏感度大和样本总体代表性高等特点[10,33]。年表公共区间分析统计特征(表2)表明:建立的红松年表样本总体代表性均达到了95%以上,都超过了EPS的临界阈值0.85[36]。差值年表样本总体代表性为0.964,说明差值年表可以很好地反映该地区红松树木年轮的基本特征。年表信噪比反映建立的年表包含气候信息的多少,两种年表的信噪比均在24.0以上,差值年表的信噪比更达到26.551,表明差值年表气候信号占非气候信号的比率较大。所建立年表的平均敏感度均在0.15左右,说明年表对外界气候变化敏感。综上分析,所建立的年表能够充分反映外界环境的变化,所选红松适合进行年轮气候学分析。本研究选用差值年表与气候因子进行相关和响应分析。
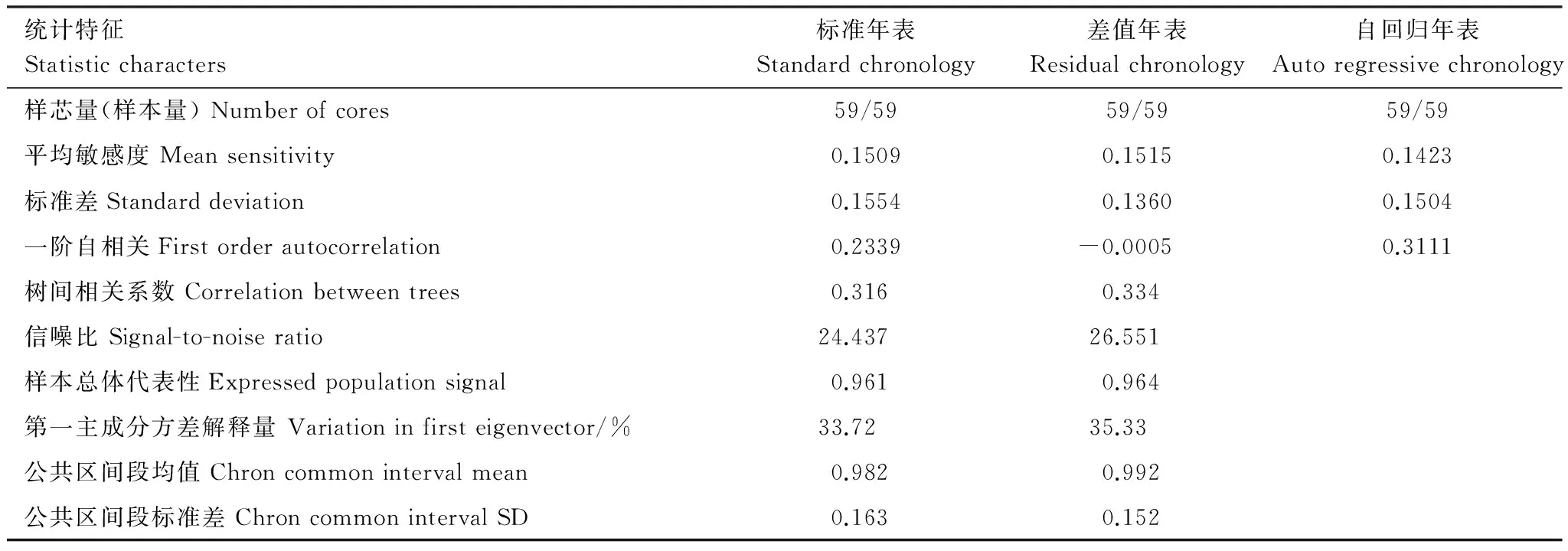
表2 年表统计特征及公共区间(1900—2008年)分析结果
2.2生物量生长量
由红松树轮径向生长过程和年均生物量生长量(髓芯对齐)可知(图5)。红松在幼龄林时期(0—60a)生长过程较快;在中龄林至成熟林阶段(61—160a)生长速度有所减缓,原因可能是树木间的竞争所致和该时期红松已处于老龄林阶段,红松径向生长缓慢;而到过熟林阶段(>161a)红松生长有加速趋势,这可能是与林分中部分树木枯死导致竞争减弱,以及近年来全球气候变暖有关。红松过熟林的快速生长也说明该地区老龄林仍然具有较强的固碳能力。根据样地调查数据,结合样地调查数据和该地区地上生物量方程计算得到目前林分单位面积生物量为310.88t/hm2,单位面积林分生物量生长量约为5.077t hm-2a-1,即每公顷每年固碳约2.539t。
2.3差值年表与逐月气候因子的关系
相关及响应函数分析结果见表3。相关分析表明,红松差值年表与当年4月平均温度、月最高温度和月最低温度呈显著正相关 (P﹤0.05),与当年9月平均温度呈显著正相关 (P﹤0.05),与上年6月最高温度呈显著正相关 (P﹤0.05),与上年7月降水量呈显著负相关 (P﹤0.05),与上年12月降水量呈显著正相关 (P﹤0.05),与当年6月PDSI呈显著正相关。响应函数分析进一步表明,红松差值年表与当年4月平均温度和最高温度呈显著正相关 (P﹤0.05),与上年7月降水量呈显著负相关 (P﹤0.05)。

表3 红松差值年表与逐月气候因子的相关及响应函数分析
P:上年,其后数字代表对应月份; *达到0.05的显著性水平
2.4年均生物量生长量与逐月气候因子的关系
相关分析表明(图 6),年均生物量生长量与当年4月的月平均温度、月平均最高温度和月平均最低温度呈显著正相关 (P﹤0.05),与上年7月的降水量呈显著负相关 (P﹤0.05),与上年12月的降水量呈显著正相关 (P﹤0.05),与上年7月的PDSI呈显著负相关,与当年6月的PDSI呈显著正相关 (P﹤0.05)。
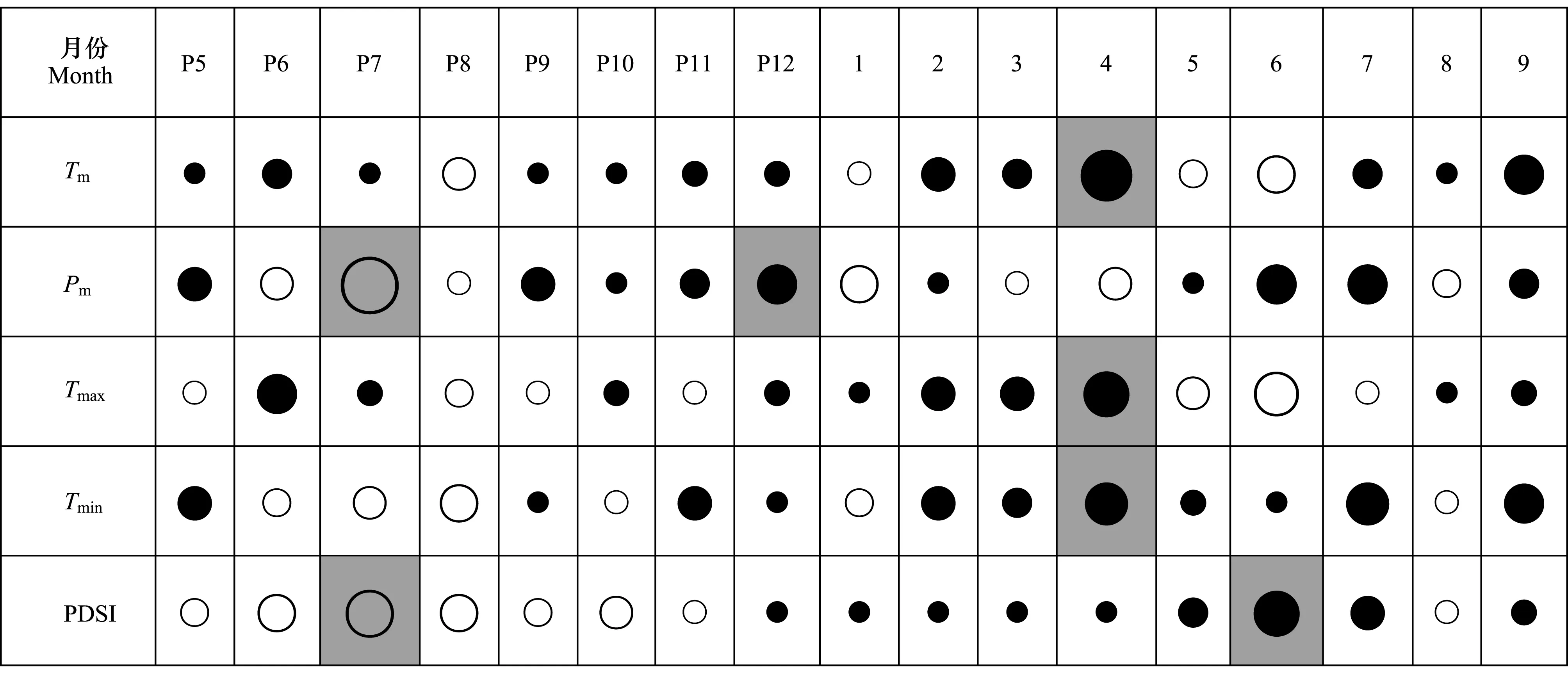
图6 红松年均生物量生长量与逐月气候因子统计值的相关性Fig.6 Correlation between mean annual biomass increment of Pinus koraiensis and monthly statistics of climate factorsTm:月平均气温 Mean monthly temperature;Tmax:月平均最高气温 Mean monthly maximum temperature;Tmin:月平均最低气温 Mean monthly minimum temperature;Pm:月总降水量 Monthly precipitation;PDSI:帕尔默干旱指数 Palmer drought severity index; P:上年;C:当年;其后数字代表对应月份; 实心圆和空心圆分别代表正相关和负相关
2.5差值年表、年均生物量生长量与气候因子的关系模拟
通过对红松差值年表、年均生物量生长量(日历年对齐)与气候因子的研究表明红松的径向生长与气候要素关系密切,本研究尝试采用多元逐步回归方程模拟红松差值年表、年均生物量生长量与气候因子之间的关系。并得到红松差值年表、年均生物量生长量与气候因子的最优方程:
Y= 0.957-0.001X1+0.035X2+0.030X3(R = 0.548, R2= 0.301, F = 5.441, P = 0.024)
式中,Y为当年树木轮宽指数;X1为上年7月降水量;X2为当年4月平均温度;X3为当年6月PDSI。
ABI= 0.976+0.035X1-0.001X2+0.032X3(R = 0.526, R2= 0.276, F = 4.188, P = 0.047)
式中,ABI表示红松年均生物量生长量;X1为当年4月平均温度;X2为上年7月降水量;X3为当年6月PDSI。
配对t检验表明,差值年表模拟值和真实值无显著差异,两者相关系数为0.547 (P﹤0.01),年均生物量生长量的模拟值与真实值也无显著差异,二者相关性为0.526 (P﹤0.01),说明该模拟方程能够拟合红松年轮宽度指数、年均生物量生长量与气候因子之间的关系。红松差值年表、年均生物量生长量的模拟回归方程各自能够解释30.1%和27.6%的红松生长变异。由回归方程可知,差值年表、年均生物量生长量与气候因子的模拟方程中的自变量均为3个,月降水量的系数很小,红松径向生长量主要受温度的影响。
3讨论
3.1气候因子对红松径向生长的影响
当年生长季前期、生长季温度和上年生长季降水对红松径向生长影响较大,这与前人对长白山红松的研究结论一致[12,14,37]。温度是红松径向生长的主要限制因子,这与长白山地区敦化的红松年轮研究结论相似[38]。长白山北坡降水量大,水分对树木径向生长影响小,因此,温度的作用尤为突出[39]。这也印证了前人研究得出长白山海拔700m以上地区,降水量不会成为树木生长的限制因子这一结论[40]。红松是喜温暖湿润的树种。长白山森林下限生长季主要集中在5—9月,4月正值春季,是树木由休眠进入生长季的关键时期,气温升高有助于地温升高和林地积雪融化,促进根系活动和地上部分萌动,因此有利于树木生长[12,41]。同为针叶树种的落叶松也受春季温度的影响[39]。红松径向生长与上年7月降水量呈显著负相关,这在以往的研究中同样有所发现[12]。7月属于红松生长最旺盛的时期,降雨量增加影响到光合作用的效率,造成体内干物质积累减少,对下一年生长不利。自当年6月PDSI基本上与差值年表呈现出持续的正相关,但这种正效应在一年的大部分时间里并不显著,在当年9月相关性达到显著水平,以往的研究也得到类似结论[42- 43]。PDSI作为表征土壤干旱程度的重要指数,不仅考虑到某一时段的土壤水分吸收,同时还综合了空气温度、地表蒸发和径流等因子,是土壤有效水分、温度和降水的综合反映,它比温度和降水单独对树木生长的影响更有意义[24]。树木生长所需的水分主要通过根部从土壤中吸收得到,自上年12月之后持续的正相关关系说明土壤干湿变化对长白山红松径向生长具有积极的作用。
3.2差值年表、连年生物量生长量与气候因子相关分析的比较
标准年表、差值年表、连年年均生物量生长量分别代表不同的含义。标准年表反映的是气候的低频变化;差值年表是在标准化年表的基础上,去掉树木个体特有的由前期生长条件对后期生长造成的持续性影响而建立的一种新年表,反映了气候的高频变化[33]。连年年均生物量生长量是反映树木生长的指标。以往采用标准年表和差值年表进行年轮气候学的研究非常普遍[44- 45],但是采用年均生物量生长量、年初级净生产力(NPP)和气候因子进行相关分析的研究仍然较少[29]。目前,国内外学者也主要是开展了断面积增长量、材积生长量等反映树木径向生长的综合指标来探讨其与气候因子的相关关系[46- 47]。本文采用年均生物量生长量进行气候相关分析,其结果与年轮宽度指数的分析结果互相印证,更能说明气候变化对树木生长的影响。
3.3气候变化对红松生长的影响
红松差值年表、连年生物量生长量与气候因子的回归分析显示,模拟回归方程仅能解释较低比例的树木生长变异,这也与以前的研究结论一致[20,48]。在长白山中海拔地区建立的红松径向生长模型同样也呈现出影响因子系数偏低的现象[37]。这可能是由于二者红松的采样高度较为接近,都位于海拔800m左右,该处的红松位于阔叶红松林群落内部,树木枝叶茂密,年轮较宽,气候条件基本可以满足树木的正常生长。因此,红松径向生长对各气候因子综合解释率偏低[33]。同时也说明气候因子是影响树木生长的重要因素,但并非全部,其他因素例如立地条件以及竞争等也可能对树木生长有重要影响[42,49]。
IPCC第五次评估报告称,1880年到2012年,全球海陆表面平均温度呈线性上升趋势,升高了0.85℃;2003年到2012年平均温度比1850年到1900年平均温度上升了0.78℃[1]。假设研究区域未来气候变化延续过去130a的变化趋势,由树木轮宽指数与气候因子模拟回归方程系数无法直接预测未来红松径向生长量的增减。然而,由红松年均生物量生长量与气候因子模拟回归方程可见,气温上升有助于红松生物量生长量增加。而这种增加趋势在其近年来红松径向生长量和年均生物量生长量变化趋势上已有体现(图4)。有报道认为,年均气温增加1℃将使红松年轮宽度增加48.4%[40]。气温升高将导致该区其他树木生长量的加速[50,39],也将促进中海拔红松生长[37],这些均与本研究结果一致。
本研究阔叶红松林地上总生物量计算结果高于前人的研究结果,但年均生物量生长量低于以往的报道[51]。许多研究表明,林龄是影响长白山林木及群落碳储量积累的主要因子之一[52]。例如,林龄是导致大兴安岭5种典型林型森林生物碳储量增加的主要原因[53]。由于群落的空间异质性,本研究的地上生长量和以往的报道有[53]较大差异。然而该区阔叶红松林已进入过熟龄阶段,树轮径向生长量较为缓慢,并且表现出年际波动,但生物量积累仍较快。长期以来人们认为老龄林已经处于碳中性状态[54]。然而有研究发现占全球森林总面积15%的老龄林仍具有固碳能力,并且固碳能力可观,其提供的森林生态系统净生产力约占全球森林生态系统净生产力的10%[20,55]。本文研究结果也支持了这一观点。
致谢:杨君珑、张国春、刘文慧和贾全全等参加野外调查、取样和部分室内分析,王晓春、高露双和封晓辉在交叉定年、年表研制等数据处理及论文写作中给予指导,特此致谢。
参考文献(References):
[1]IPCC. Working Group I Contribution to the IPCC Fifth Assessment Report (AR5). Climate Change 2013: The Physical Science Basis. Final Draft Underlying Scientific-Technical Assessment. [2013-10-30].
[2]Mann M E, Bradley R S, Hughes M K. Northern hemisphere temperatures during the past millennium: inferences, uncertainties, and limitations. Geophysical Research Letters, 1999, 26(6): 759- 762.
[3]郭志梅, 缪启龙, 李雄. 中国北方地区近50年来气温变化特征的研究. 地理科学, 2005, 25(4): 448- 454.
[4]翟盘茂, 邹旭恺. 1951—2003年中国气温和降水变化及其对干旱的影响. 气候变化研究进展, 2005, 1(1): 16- 18.
[5]Neilson R P. Vegetation redistribution: A possible biosphere source of CO2during climate change. Water, Air, & Soil Pollution, 1993, 70(1/4): 659- 673.
[6]邵雪梅, 吴祥定. 利用树轮资料重建长白山区过去气候变化. 第四纪研究, 1997, (1): 76- 85.
[7]阳含熙, 谢海生. 长白山红松混交林干扰历史的重构研究. 植物生态学报, 1994, 18(3): 201- 208.
[8]王战, 徐振邦, 李昕. 长白山北坡主要森林类型及其群落结构特点 (之一) // 王战. 森林生态定位研究. 沈阳: 中国科学院长白山森林生态系统定位站, 1980.
[9]Hasenaur H, Nemani R R, Schadauer K, Running S W. Forest growth response to changing climate between 1961 and 1990 in Austria. Forest Ecology and Management, 1999, 122(3): 209- 219.
[10]高露双, 赵秀海, 王晓明. 火干扰后红松生长与气候因子的关系. 生态学报, 2009, 29(11): 5963- 5970.
[11]高露双, 王晓明, 赵秀海. 长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应. 植物生态学报, 2011, 35(1): 27- 34.
[12]陈力, 吴绍洪, 戴尔阜. 长白山红松和落叶松树轮宽度年表特征. 地理研究, 2011, 30(6): 1147- 1155.
[13]陈力, 尹云鹤, 赵东升, 苑全治, 吴绍洪. 长白山不同海拔树木生长对气候变化的响应差异. 生态学报, 2014, 34(6): 1568- 1574.
[14]陈列, 高露双, 张赟, 张思行, 赵秀海. 长白山北坡不同林型内红松年表特征及其与气候因子的关系. 生态学报, 2013, 33(4): 1285- 1291.
[15]王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡不同年龄红松年表及其对气候的响应. 生态学报, 2011, 31(21): 6378- 6387.
[16]张雪, 高露双, 丘阳, 郭静. 长白山红松 (Pinuskoraiensis) 不同树高处径向生长特征及其对气候响应研究. 生态学报, 2015, 35(9): 1- 10.
[17]高露双, 赵秀海, 王晓明. 长白山火烧红松年表特征分析. 林业科学, 2011, 47(3): 189- 193.
[18]Fritts H C. Relationships of ring widths in arid-site conifers to variations in monthly temperature and precipitation. Ecological Monographs, 1974, 44(4): 411- 440.
[19]张文涛, 江源, 王明昌, 张凌楠, 董满宇, 郭媛媛. 芦芽山阳坡不同海拔白杄径向生长对气候变暖的响应. 植物生态学报, 2013, 37(12): 1142- 1152.
[20]Lin B, Xu Q Q, Liu W H, Zhang G C, Xu Q Y, Liu Q J. Dendrochronology-based stand growth estimation ofLarixolgensisforest in relation with climate on the eastern slope of Changbai Mountain, NE China. Frontiers of Earth Science, 2013, 7(4): 429- 438.
[21]Kendall M G. Rank Correlation Methods. London: Griffin, 1970: 125- 130.
[22]Kohler M A. On the use of double-mass analysis for testing the consistency of meteorological records and for making required adjustments. Bulletin of the American Meteorological Society, 1949, 82: 96- 97.
[23]Palmer W C. Meteorological Drought Research Paper 19, No. 45. Washington D. C.: Weather Bureau, 1965.
[24]Alley W M. The palmer drought severity index as a measure of hydrologic drought. Journal of American Water Resources Association, 1985, 21(1): 105- 114.
[25]Stokes M A, Smiley T I J. An Introduction to Tree-Ring Dating. Chicago: The University of Chicago Press, 1968.
[26]Rozas V. A dendroecological reconstruction of age structure and past management in an old-growth pollarded parkland in northernSpain. Forest Ecology and Management, 2004, 195(1/2): 205- 219.
[27]Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69- 75.
[28]Cook E R, Holmes R L. Users manual for ARSTAN: Laboratory of Tree-ring Research. Tucson: University of Arizona, 1986.
[29]程瑞梅, 封晓辉, 肖文发, 王瑞丽, 王晓荣, 杜化堂. 北亚热带马尾松净生产力对气候变化的响应. 生态学报, 2011, 31(8): 2086- 2095.
[30]陈传国, 朱俊凤. 东北主要林木生物量手册. 北京: 中国林业出版社, 1989.
[31]罗天祥. 中国主要森林类型生物生产力格局及其数学模型[D]. 北京: 中国科学院地理科学与资源研究所, 1996.
[32]徐振邦, 李昕, 戴洪才, 谭征祥, 章依平, 郭杏芬, 彭永山, 代力民. 长白山阔叶红松林生物生产量的研究. 森林生态系统研究, 1985, (5): 33- 48.
[33]吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990.
[34]Fritts H C. Tree Rings and Climate. London: Academic Press, 1976: 64- 90, 1- 242, 376- 412.
[35]Biondi F, Waikul K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree- ring chronologies. Computer & Geoscience, 2004, 30(3): 303- 311.
[36]Wigley T M L, Briffa K R, Jones P D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Climate and Applied Meteorology, 1984, 23(2): 201- 213.
[37]高露双. 长白山典型树种径向生长与气候因子的关系研究[D]. 北京: 北京林业大学, 2011.
[38]李牧, 王晓春. 敦化三大硬阔、红松年轮——气候关系及生长季低温重建. 南京林业大学学报: 自然科学版, 2013, 37(3): 29- 34.
[39]于大炮, 王顺忠, 唐立娜, 代力民, 王庆礼, 王绍先. 长白山北坡落叶松年轮年表及其与气候变化的关系. 应用生态学报, 2005, 16(1): 14- 20.
[40]王淼, 白淑菊, 陶大立, 单建平. 大气增温对长白山林木直径生长的影响. 应用生态学报, 1995, 6(2): 128- 132.
[41]李广起, 白帆, 桑卫国. 长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应. 植物生态学报, 2011, 35(5): 500- 511.
[42]Foster T E, Brooks J R. Long-term trends in growth ofPinuspalustrisandPinuselliottiialong a hydrological gradient in central Florida. Canadian Journal of Forest Research, 2001, 31(10): 1661- 1670.
[43]Henderson J P, Grissino-Mayer H D. Climate-tree growth relationships of longleaf pine (PinuspalustrisMill.) in the Southeastern Coastal Plain, USA. Dendrochronologia, 2009, 27(1): 31- 43.
[44]王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系. 植物生态学报, 2011, 35(3): 294- 302.
[45]张先亮, 何兴元, 陈振举, 崔明星, 黎娜. 大兴安岭山地樟子松径向生长对气候变暖的响应——以满归地区为例. 应用生态学报, 2011, 22(12): 3101- 3108.
[46]Moore D J P, Aref S, Ho R M, Pippen J S, Hamilton J G, De Lucia E H. Annual basal area increment and growth duration of Pinus taeda in response to eight years of free- air carbon dioxide enrichment. Global Change Biology, 2006, 12(8): 1367- 1377.
[47]Linares J C, Delgado-Huertas A, Carreira J A. Climatic trends and different drought adaptive capacity and vulnerability in a mixedAbiespinsapo-Pinushalepensisforest. Climate Change, 2011, 105(1/2): 67- 90.
[48]Yu D P, Wang G G, Dai L M, Wang Q L. 2007. Dendroclimatic analysis ofBetulaermaniiforests at their upper limit of distribution in Changbai Mountain, Northeast China. Forest Ecology and Management, 240(1/3): 105- 113.
[49]Lo Y H, Blanco J A, Seely B, Welham C, Kimmins J P. Relationships between climate and tree radial growth in interior British Columbia, Canada. Forest Ecology and Management, 2010, 259(5): 932- 942.
[50]吴祥定, 邵雪梅. 采用树轮宽度资料分析气候变化对树木生长量影响的尝试. 地理学报, 1996, (S1): 92- 101.
[51]李文华, 邓坤枚, 李飞. 长白山主要生态系统生物生产量的研究. 森林生态系统研究, 1981, (2): 34- 50.
[52]马炜, 孙玉军, 郭孝玉, 巨文珍, 穆景森. 不同林龄长白落叶松人工林碳储量. 生态学报, 2010, 30(17): 4659- 4667.
[53]胡海清, 罗碧珍, 魏书精, 魏书威, 文正敏, 孙龙, 罗斯生, 王立明, 马洪斌. 大兴安岭5种典型林型森林生物碳储量研究. 生态学报, 2015, 35(17): 1- 21.
[54]Jarvis P G, Morison J I L, Chaloner W G, Cannell M G R, Roberts J, Jones H G, Amtmann R. Atmospheric carbon dioxide and forests. Series B: Philosophical Transactions of the Royal Society of London, 1989, 324(1223): 369- 392.
[55]Luyssaert S, Schulze E, Börner A, Knohl A, Hessenmöller D, Law B E, Ciais P, Grace J. Old-growth forests as global carbon sinks. Nature, 2008, 455(7210): 213- 215.
Radial growth ofPinuskoraiensisand carbon sequastration potential of the old growth forest in Changbai Mountain, Northeast China
YU Jian1,2, LUO Chunwang1, XU Qianqian3, MENG Shengwang1, LI Junqing1, LIU Qijing1,*
1TheKeyLaboratoryforSilvicultureandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China2JiangsuPolytechnicCollegeofAgricultureandForestry,Jurong212400,China3ZhengzhouFruitResearchInstitute,ChineseAcademyofAgriculturalSciences,Zhengzhou450009,China
Abstract:The vegetation of Changbai Mountain, located in the heartland of temperate coniferous and broad-leaved mixed forest, is subject to climate change. Using the basic principles and methods of dendrochronology, and Pinus koraiensis growing on northern slopes of Changbai Mountain as objects, objectives of this work were to: (1) study the variation of tree-ring width of P. koraiensis to better understand the response of the forest ecosystem to climate change; (2) establish chronologies of P. koraiensis, correlation, and response functions with monthly climatic factors, and calculate the average biomass growth in successive years; and (3) establish regression equations for tree-ring index and the average annual biomass increment of P. koraiensis. The main conclusions are: (1) Growth of P. koraiensis was sensitive to climatic factors according to the analysis of chronology, which is suitable for dendroclimatological study. (2) Radial growth of P. koraiensis was mainly affected by the temperature of the early growing season. Radial growth was more strongly correlated to temperature than to precipitation. (3) Temperature was the main factor limiting growth, which indicated that climate warming might accelerate tree growth. (4) At present, the aboveground biomass of P. koraiensis growing on northern slopes of Changbai Mountain is 310.88 t ha-1, with an increasing rate of 5.077 t hm-2a-1or 2.539 t carbon, indicating that the old-growth forest of P. koraiensis in the region still has a strong potential for carbon sequestration.
Key Words:tree-ring width; climate warming; Pinus koraiensis; biomass increment; carbon sequestration potential
基金项目:国家863计划项目(2013AA122003); 国家林业公益性行业科研专项项目(200804001)
收稿日期:2014- 11- 15; 网络出版日期:2015- 08- 25
*通讯作者
Corresponding author.E-mail: liuqijing@bjfu.edu.cn
DOI:10.5846/stxb201411152263
于健,罗春旺,徐倩倩,孟盛旺,李俊清,刘琪璟.长白山原始林红松径向生长及林分碳汇潜力.生态学报,2016,36(9):2626- 2636.
Yu J, Luo C W, Xu Q Q, Meng S W, Li J Q, Liu Q J.Radial growth ofPinuskoraiensisand carbon sequastration potential of the old growth forest in Changbai Mountain, Northeast China.Acta Ecologica Sinica,2016,36(9):2626- 2636.
