不同生境外生菌根真菌对铝胁迫的响应
2016-07-21周志峰王明霞黄建国
周志峰, 王明霞, 袁 玲, 黄建国
西南大学资源环境学院, 重庆 400716
不同生境外生菌根真菌对铝胁迫的响应
周志峰, 王明霞*, 袁玲, 黄建国
西南大学资源环境学院, 重庆400716
摘要:以南北方不同生境下的10株外生菌根真菌为研究对象,采用液体培养的方法,研究了铝对不同菌根真菌的生物量、有机酸分泌及养分含量的影响,以期筛选出抗铝性强的优良菌株,并探讨其抗铝机理。结果表明:外生菌根真菌Sl 08抗铝性最强;Pt 715、Ld 03、Bo 11、Sl 01、Bo 15也具有不同程度的耐铝性;Sl 14、Gc 99、Cg 04抗铝性较差;Sg 11抗铝性最差。来自南方酸性森林土壤的菌株总体抗铝性强于来自北方石灰性土壤的菌株,这表明外生菌根真菌的铝耐受能力与其原始生境有着密切的联系。外生菌根真菌能分泌多种有机酸,且不同菌株分泌的有机酸种类不同。其中,受铝胁迫分泌量增加最多的是草酸。研究中,铝胁迫能增加大多数铝抗性菌株的草酸分泌量,其中铝抗性最强的Sl 08表现最为明显。但铝胁迫并没有促进具备一定铝抗性的Bo 11 和Sl 01草酸的分泌量,同时在铝敏感的菌株中均观察到了草酸分泌量的增加。这表明分泌草酸可能并不是外生菌根真菌抵抗铝毒的唯一途径。对各菌株铝胁迫下对氮,磷及钾的吸收研究表明,除铝敏感菌株Sl 14外,铝胁迫均能促进各供试菌株对氮,磷或钾的吸收。综上,在一定铝浓度下,一些外生菌根真菌可通过增加草酸分泌来抵御铝毒。此外,铝胁迫下外生菌根真菌还可通过调控氮、磷、钾等营养元素的吸收来抵抗铝毒,即通过增加对营养元素的吸收来增强其在铝胁迫下的生存能力,这可能是其抵御铝胁迫的应激反应之一。
关键词:环境;外生菌根;铝;有机酸;养分
自20世纪80年代以来,由于酸性沉降物(特别是酸雨)、施肥等人为因素而导致土壤酸化的环境问题,开始引起研究学者的极大关注。研究表明,土壤或水体酸化导致的活性铝增加是酸性土壤中限制植物生长的主要因素[1]。而一些优良的外生菌根(ECM)真菌能显著增加宿主植物的抗铝性[2- 4]。在一些常规造林困难的地区,选择适宜的树种和适应性强的ECM真菌,采用人工培育菌根苗技术可以大大提高造林的成活率,加速植被的恢复。然而,不同菌根真菌对铝的抗性大不相同。例如,在铝浓度为0.03 mmol/L的培养液中,Lacatariusrufus和L.hepaticus对铝非常敏感,而L.bicolor对铝有较强的抗性。在培养基铝浓度为0.5 mmol/L时,L.bicolor和Pisolithustinctorius生长量不减少[5-6]。而Laccarialaccata在铝浓度为0.37 mmol/L时表现敏感[7]。研究表明,菌根植物根系的有机酸分泌量增加与其抗铝能力有关[8-9]。此外,菌根真菌侵染植物后可增加磷、钾、钙、镁等营养元素的吸收,有益于提高其抗铝性[10-11]。但这些研究结果大多来自于欧美等国的菌种资源,对于我国本土菌株的研究还很少。本研究选用我国南北方不同生态条件下的10株ECM真菌,采用液体培养的方法,研究了铝胁迫下ECM真菌的一些生理生化反应,以期筛选出抗铝型和铝敏感型菌株,探讨ECM真菌抗铝性与其有机酸分泌、养分吸收的关系,为进一步明确ECM真菌耐受铝胁迫的机理提供理论基础,对酸性土壤的天然林保护和植被恢复也具有重要意义。
1材料与方法
1.1供试材料
本试验选用10株来自我国南、北方不同生态条件下的外生菌根真菌。它们是:彩色豆马勃(Pisolithustinctorius)Pt715,采自四川西昌桉树林下;褐环乳牛肝菌(Suillusluteus)Sl08和Sl14、牛肝菌(Boletus)Bo15和Bo11,均采自重庆金佛山马尾松林下;松乳菇(Lactariusdeliciosus)Ld03,采自重庆缙云山马尾松林下;褐环乳牛肝菌Sl01、黄空柄牛肝菌(Gyroporuscyanescens)Gc99、厚环乳牛肝菌(Suillusgrevillei)Sg11和土生空团菌(Cenococcumgeophilum)Cg04,均采自内蒙古大青山油松林下。
1.2试验设计
取在4℃下保存1—2个月的上述菌株,接种于Pachlewski固体培养基[12]之上,在25℃条件下暗培养21 d备用。
用分析纯Al2(SO4)3·18H2O提供活性铝,配成Al3+浓度分别为0、0.2、0.4、1.0 mmol/L的Pachlewski液体培养基,形成无铝(对照,CK)、低铝、中铝、高铝4个处理[13-14]。取150 mL三角瓶分装不同处理的液体培养基,每瓶20 mL,封口后于121℃高压蒸汽灭菌20 min。冷却后接种直径为6 mm的供试琼脂菌种1个,在25℃的黑暗条件下静置培养21 d待测。各处理重复5次。
1.3测定项目与方法
培养结束后,过滤收集菌丝和滤液。菌丝用于测定生物量和氮(N)、磷(P)、钾(K)含量。菌丝生物量采用称重法获得。烘干的菌丝体经H2SO4和H2O2消化后,采用奈氏试剂分光光度计法测定N含量,钼黄分光光度计法测定P含量,火焰光度法测定K含量[15]。滤液用于测定H+分泌量与有机酸的种类、含量。H+分泌量采用pH计法测定[16],换算成H+浓度后除以生物量,得到单位质量的H+分泌量。滤液再经定容,过滤(0.22 μm微孔滤膜)后,用外标法于高效液相色谱仪上分析不同有机酸的含量[12]。检测的有机酸包括草酸、柠檬酸、酒石酸、琥珀酸、乳酸、甲酸和乙酸。
1.4数据处理与分析
试验数据采用Excel进行基本计算,SPSS 14.0统计软件进行统计分析,采用LSD法进行多重比较,显著水平P<0.05。
2结果与分析
2.1铝对外生菌根真菌生物量的影响
不同菌株对铝处理的反应不同(图1)。在0.2 mmol Al3+/L培养液中,来自内蒙古大青山森林土壤中的Sl11、Gc99、Cg04的生物量减少,其中Sl11比对照(无铝)减少了32.9%;其余7个菌株的生物量均有所增加,但差异未达到显著水平。当铝浓度为0.4 mmol/L时,大部分菌丝的生物量开始下降(除了Sl01),其中Sl11、Gc99、Cg04和Sl14的生物量显著低于对照,分别比对照减少了56.6%(Sl11)、8.2%(Gc99)、31.6%(Cg04)和14.3%(Sl14)。当铝浓度达到1.0 mmol/L时,菌株生物量继续下降。除Sl08外,其他几个菌株生物量均低于对照。其中Sl11比对照减少了70.7%,减少幅度最大。总体看来,Sl08生物量受铝影响不显著,在高铝处理下仍有所增加,表现出较强的抗铝性;Pt715、Ld03、Bo11、Sl01、Bo15不同程度地具有一定抗铝性;Sl14、Gc99、Cg04抗铝性较差;Sg11抗铝性最差。
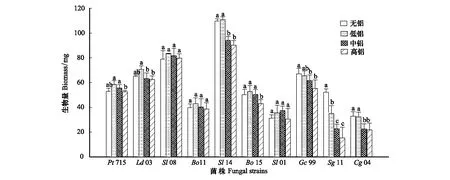
图1 不同铝处理下外生菌根真菌的生物量Fig.1 Biomass of the ECM fungal isolates in culture solution with different Al3+ concentrations图中同一菌株的不同字母表示差异显著(P<0.05)
2.2铝对外生菌根真菌有机酸分泌的影响
2.2.1H+分泌
由图2可见,除Sg11外,其他9个菌株H+分泌量均随培养液中Al3+浓度的增加而显著增加。当铝浓度达到1.0 mmol/L时,H+分泌量分别比对照增加了286.9%(Pt715)、228.4%(Ld03)、76.9%(Sl08)、204.0%(Bo11)、105.2(Sl14)、62.6%(Bo15)、552.1%(Sl01)、106.2(Gc99)和58.8%(Cg04)。Sg11的H+分泌量随培养液中铝浓度的增加而减少,高铝处理时减少为对照的30.5%。

图2 铝对外生菌根真菌H+分泌的影响Fig.2 Effects of Al3+on proton effluxes by ECM fungi
2.2.2有机酸分泌
本试验在ECM真菌培养液中检测出含有乙酸、柠檬酸、草酸、琥珀酸和酒石酸。将低铝、中铝和高铝处理的有机酸分泌量取平均值得到各菌株有铝(+Al)处理的有机酸分泌量。由表1可见,所有供试菌株均检测出有草酸分泌,且分泌量在众多有机酸中最多,成为本研究关注的重点。对于乙酸的分泌,Pt715和Sl14未检测出;其他菌株的分泌量均在加铝后减少,其中Ld03和Sg11加铝后未检测出。供试菌株中只有Sl08和Bo15检测出有柠檬酸分泌,Sl08加铝后柠檬酸分泌量增加至无铝(-Al)的1.44倍,Bo15加铝后减少至无铝的39.0%。不同菌株琥珀酸的变化趋势不一致。Bo11、Sl01、Sg11和Cg04无论在有铝还是无铝时均未检测出琥珀酸分泌。Sl08和Gc99在无铝培养时有少量琥珀酸分泌,加铝后未检测出。加铝培养后,Pt715的琥珀酸分泌量减少为对照的71.7%,而Ld03、Sl14和Bo15的分泌量增加。由于Pachlewski培养基中含有酒石酸氨,因此检测出来的酒石酸可能并不是菌根真菌所分泌的,而是其消耗所剩余的酒石酸盐。Pt715和Ld03无论在无铝还是有铝时,均未检测出酒石酸存在,可能已基本消耗殆尽。其他8个菌株在加铝后酒石酸含量均有不同程度地减少,说明铝胁迫下外生菌根真菌需消耗更多的酒石酸盐。
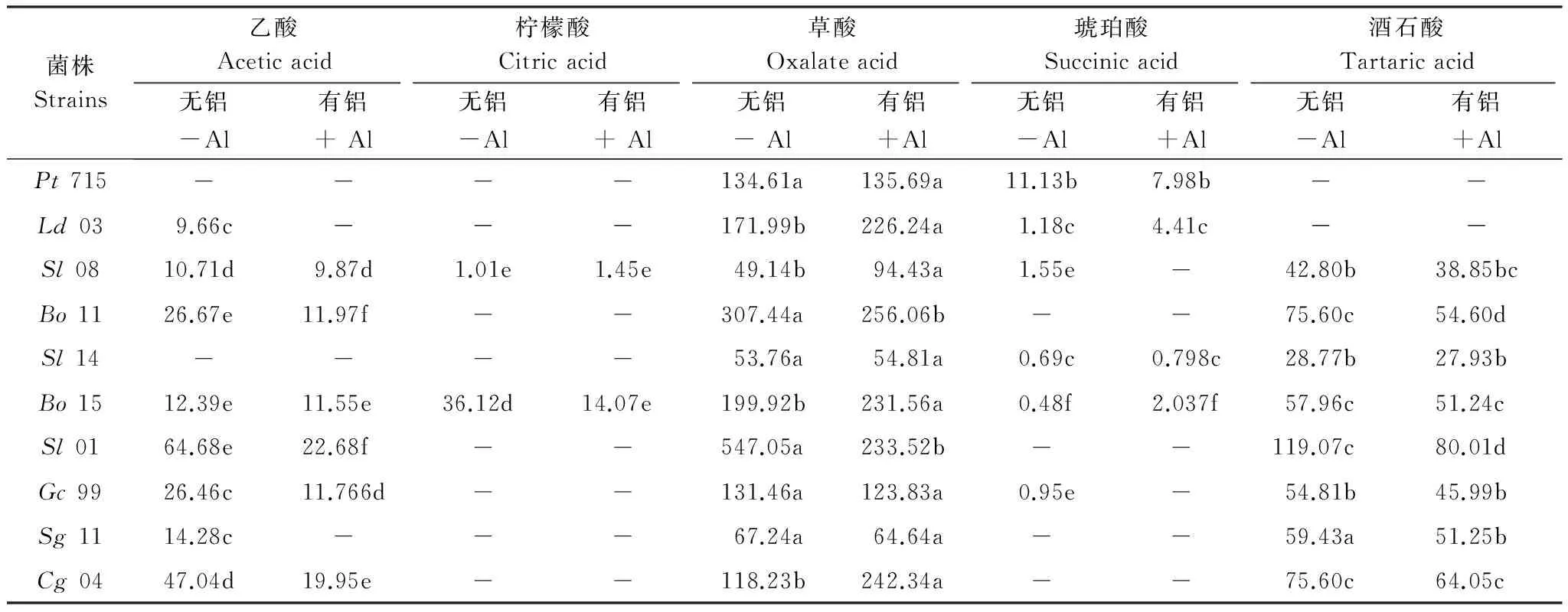
表1 铝对外生菌根真菌有机酸分泌的影响/(μmol/g干重)
表中同一行中的不同字母表示差异显著(P<0.05), -表示未检测出
2.2.3草酸分泌
由图3可见,铝处理后,Bo11、Sl01的草酸分泌量显著降低,高铝处理时分别减少为对照的82.5%和30.3%。其它8个菌株的草酸分泌均随铝浓度的增加而呈先增加后降低的趋势。草酸分泌量增加最多的是Cg04,中铝时增加至对照的2.40倍。其次为Sl08,中铝时增加至对照的2.14倍。
将各菌株4个处理的草酸分泌量取平均值,得到各菌株的平均分泌量。各菌株草酸分泌量差异显著。分泌量最多的是Sl01,平均分泌量为311.90 μmol/g干重,其次为Bo11(268.91 μmol/g干重)>Bo15(228.65 μmol/g干重)>Ld03(212.68 μmol/g干重)>Cg04(183.56 μmol/g干重)>Pt715(135.42 μmol/g干重)>Gc99(125.74 μmol/g干重)>Sl08(83.11 μmol/g干重)>Sg11(65.29 μmol/g干重)>Sl14(54.55 μmol/g干重)。草酸分泌量与菌根真菌抗铝性无显著相关性。因此,草酸分泌的绝对含量并不能成为衡量外生菌根真菌抗铝性的指标。

图3 铝对外生菌根真菌草酸分泌量的影响Fig.3 Effects of Al3+ on the quantity of oxalate effluxes by ECM fungi
2.3铝对外生菌根真菌氮磷钾含量的影响
2.3.1铝对外生菌根真菌氮含量的影响
由表2可见,受铝胁迫后,Pt715、Sl14和Cg04的含氮量随培养液铝浓度的增加而增加。高铝处理后,含氮量比对照增加了57.1%(Pt715)、17.1%(Sl14)和47.5%(Cg04)。Sl08、Bo11、Sl01和Gc99的含氮量随培养液铝浓度的增加呈现先增加后降低的趋势,且均在中铝处理时达到最大值,分别比对照增加了2.17%(Sl08)、45.5%(Bo11)、253.5%(Sl01)和25.9%(Gc99)。而Ld03、Bo15和Sg11的含氮量随培养液铝浓度的增加而降低。高铝处理时,含氮量比对照减少了17.5%(Ld03)、25.8%(Bo15)和34.3%(Sg11)。
2.3.2铝对外生菌根真菌磷含量的影响
由表3可见,受铝胁迫后,Pt715和Ld03的含磷量随培养液铝浓度的增加而增加。高铝处理时,含磷量分别比对照增加了83.0%和51.0%。Bo11、Bo15、Sg11和Cg04含磷量随培养液铝浓度的增加呈现先增加后降低的趋势。Bo11和Bo15在低铝处理时磷含量达到最大值,分别比对照增加了95.9%和35.1%。Sg11和Cg04在中铝处理时磷含量达到最大值,分别比对照增加了176.0%%和92.3%。Sl08、Sl14、Sl01和Gc99的含磷量随培养液铝浓度的增加变化不显著。
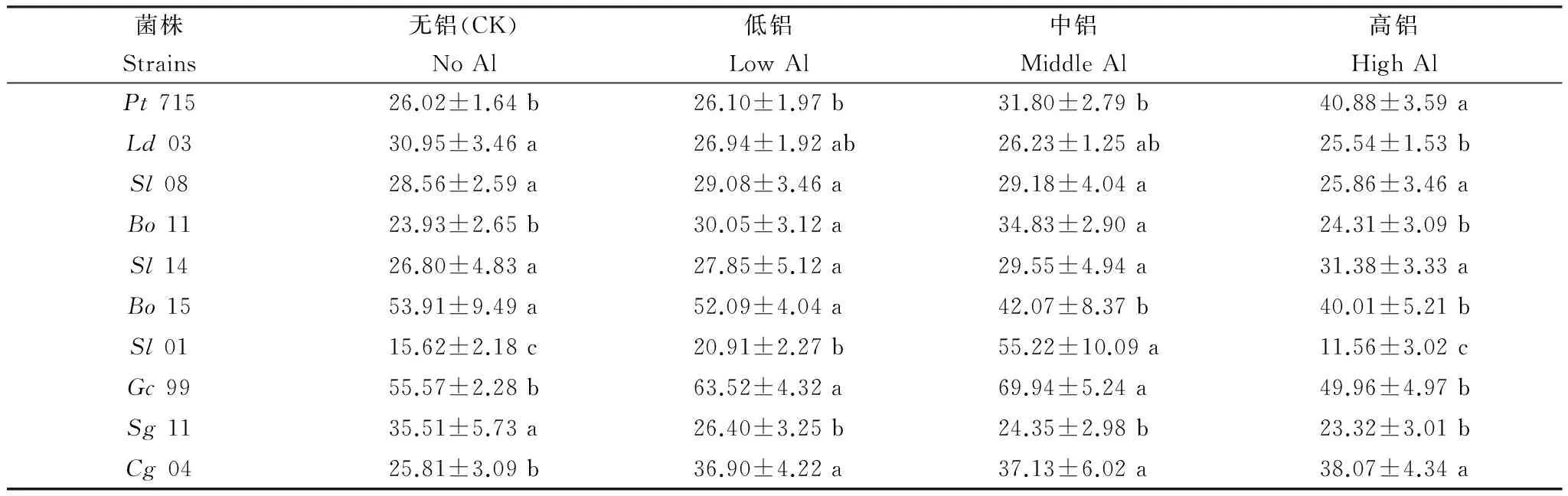
表2 铝对外生菌根真菌氮含量的影响/(mg/g干重)

表3 铝对外生菌根真菌磷含量的影响/(mg/g干重)
2.3.3铝对外生菌根真菌钾含量的影响
由表4可见,铝处理后,Ld03、Sl14和Gc99的含钾量变化不显著。Bo15的含钾量在中、高铝处理时显著降低,分别降至对照的27.0%和18.9%。其余6个菌株的含钾量均在一定铝浓度范围内显著增加。Pt715和Sg11的含钾量在中铝处理时达到最大值,比对照增加了27.7%和55.0%。Sl08、Bo11、Sl01和Cg04在低铝处理时达到最大值,分别比对照增加了34.7%(Sl08)、128.50%(Bo11)、24.9%(Sl01)和48.2%(Cg04)。

表4 铝对外生菌根真菌钾含量的影响/(mg/g干重)
3讨论
3.1铝对外生菌根真菌生长的影响
在酸性森林土壤中,外生菌根显著提高树木的抗铝性受到众多学者的关注[17-18]。但是,不同菌种(株)拮抗铝毒的能力不同。在铝胁迫条件下,菌丝生长量是指示外生菌根真菌抗铝能力的可靠指标之一[16]。本实验中,不同菌株对铝的抗性也不同。在0.2 mmol Al3+/L培养液中,来自内蒙古大青山森林土壤的菌株Sl11、Gc99、Cg04的生物量减少。其中Sl11生物量比对照(无铝)减少了32.9%,显示出较差的抗铝性。其余来自南方酸性森林土壤的6个菌株和来自内蒙古大青山森林土壤的Sl01的生物量均有所增加,但差异未达到显著水平。当铝浓度为0.4 mmol/L时,大部分菌丝的生物量开始下降(除Sl01外),其中Sl14生物量显著低于对照,比对照减少了14.3%。当铝浓度达到1.0 mmol/L时,大部分菌株生物量均低于对照,只有Sl08菌株生物量仍高于对照,表现出较强的抗铝性。由此看来,Sl08抗铝性最强;Pt715、Ld03、Bo11、Sl01、Bo15不同程度地具有一定抗铝性;Sl14、Gc99、Cg04抗铝性较差;Sg11抗铝性最差。同时,来自南方酸性森林土壤的菌株总体抗铝性强于来自北方石灰性土壤的菌株。前人的研究表明,降低pH值和增加活性铝浓度能影响土壤中ECM真菌的种类,甚至影响其对松树根系的侵染[19]。在欧洲,ECM真菌种类减少已经被认为与森林土壤酸化和铝毒有关[20]。因此,在酸性森林土壤中,长期的自然选择和进化导致铝敏感菌株逐渐消失,而耐铝型菌株得以幸存。我国南方马尾松林心土层pH值一般小于4.5,属于强酸性森林土壤,活性铝含量很高。本文中来自南方酸性森林土壤的菌株抗铝性强于来自北方石灰性土壤的菌株的研究结果,是符合自然选择的规律的。
3.2铝对外生菌根真菌有机酸分泌的影响
铝胁迫下,外生菌根真菌能分泌多种有机酸,包括甲酸、乙酸、草酸、琥珀酸、柠檬酸、苹果酸等[17,21-22]。这些有机酸能与铝形成稳定的螯合物,增大分子量,增加分子体积,使之不能通过细胞膜上的离子通道,降低菌丝和根系细胞对铝的吸收。本研究的10个供试菌株中,有9个菌株的H+分泌量均随培养液中铝浓度的增加而显著增加,其H+分泌量与培养液铝浓度呈极显著正相关(P<0.01)。说明有机酸分泌是外生菌根真菌拮抗铝的重要机制之一。其分泌的有机酸种类有柠檬酸、苹果酸、琥珀酸、乙酸、草酸等。但不同菌株分泌的有机酸种类不同。其中,受铝胁迫分泌量增加最多的是草酸。这与前人的研究结果一致[23-24]。
草酸根离子对二、三价阳离子具有较强的络合力[13,25]。[A1(C2O4)3]3-的稳定常数为2.0×1016,分子量比铝增大9.8倍,既减小了菌丝周围的Al3+活性,又减缓了Al3+通过菌丝细胞膜的速率,抑制铝的吸收。Pinusdensiflora接种外生菌根真菌后,乙醛酸、草酸和柠檬酸分泌量增加,抗铝能力随之提高[21]。因此,大多数学者认为,草酸分泌量与外生菌根真菌拮抗铝胁迫的能力密切相关[26-27]。本研究中,在铝胁迫下,抗铝性最强的Sl08的草酸分泌量显著增加,增加为对照的1.79—2.14倍。有一定抗铝性的Pt715、Ld03和Bo15的草酸分泌增加幅度不及Sl08,其中Ld03在中铝条件下的草酸分泌量,增加为对照的1.53倍。这说明在一定铝浓度下一些外生菌根真菌可通过增加草酸分泌来抵御铝毒。而有一定抗铝性的Bo11、Sl01的草酸分泌量显著降低,对铝较敏感的Cg04草酸分泌量显著增加,说明草酸分泌不是ECM真菌拮抗铝毒的唯一机制。铝胁迫下,铝敏感菌株Sg11、Sl14、Gc99的草酸分泌量会有所增加,主要是由于其生物量受铝胁迫减少较多,从而导致其单位菌丝的草酸分泌量有所增加。
3.3铝对外生菌根真菌氮磷钾含量的影响
外生菌根真菌与植物根系共生后,所形成的外生菌根能促进植物养分吸收,其中包括N、P、K等[28-29]。例如,铝胁迫下接种外生菌根真菌Pisolithustinctorius可显著增加pinusrigida根茎的含磷量[6]。纯培养条件下,无论是抗铝还是铝敏感型菌种铝胁迫后都会发生一系列有益于抗铝的生化反应,其中包括菌丝磷钾含量的增加[16]。本研究中,在铝处理后,抗铝菌株Pt715和Bo11的N、P、K含量均显著增加;Sl01的N、K含量显著增加;Sl08的K含量显著增加;Ld03和Bo15的P含量显著增加,N含量显著减少。铝处理后,敏感菌株Cg04的N、P、K含量均显著增加;Sg11的P、K含量显著增加,N含量显著减少;Gc99的N含量显著增加;Sl14的N、P、K变化均不显著。虽然不同菌株的养分含量对铝的响应有所不同,但可以看出多数菌株表现为在受铝胁迫后N、P、K含量增加,这可能是其抵御铝胁迫的应激反应之一。研究表明,铝可以聚磷酸盐的形式累积于Pisolithustinctorius菌套菌丝液泡和细胞壁内,使铝以无毒形态隔离[6]。菌根真菌增加P吸收有利于降低Al3+活性,从而缓解铝毒。氮是蛋白质、遗传材料和其它关键有机分子的基本组成元素。钾是生物体内多种酶的活化剂。增加N、K的吸收有益于生物代谢,减轻铝毒。因此,在铝胁迫条件下,菌丝体内的N、K含量增加对于ECM真菌拮抗铝毒有积极意义。
参考文献(References):
[1]Huttová J, Tamás L, Mistrík I. Aluminium induced acid phosphatase activity in roots of Al-sensitive and Al-tolerant barley varieties. Rostlinna Vyroba, 2002, 48 (12): 556- 559.
[2]Hartley J, Cairney J W G, Freestone P, Woods C, Meharg A A. The effects of multiple metal contamination on ectomycorrhizal Scots pine (Pinussylvestris) seedlings. Environmental Pollution, 1999, 106(3): 413- 424.
[3]Smith S E, Read D J. Mycorrhizal symbiosis. London: Academic Press, 2008: 145- 156.
[4]Cumming J R, Weinstein L H. Aluminum-mycorrhizal interactions in the physiology of pitch pine seedlings. Plant and Soil, 1990, 125(1): 7- 18.
[5]Cumming J R, Swiger T D, Kurnik B S Panaccione D G. Organic acid exudation byLaccariabicolorandPisolithustinctoriusexposed to aluminum in vitro. Canadian Journal of Forest Research, 2001, 31(4): 703- 710.
[6]Schier G A, McQucttie C J. Response of ectomycorrhizal and nonmycorrhizal pitch pine (Pinusrigida) seedlings to nutrient supply and aluminum: growth and mineral nutrition. Canadian Journal of Forest Research, 1996, 26(12): 2145- 2152.
[7]Jones D, Muehlchen A. Effects of the potentially toxic metals, aluminium, zinc and copper on ectomycorrhizal fungi. Journal of Environmental Science and Health, 1994, 29(5): 949- 966.
[8]Foramina M, Hillier S, Charnock J M, Melville K, Alexander I J, Gadd G M. Role of oxalic acid overexcretion in transformations of toxic metal minerals byBeauveriacaledonica. Applied and Environmental Microbiology,2005, 71(1): 371- 381.
[9]Bellion M, Courbot M, Jacob C, Blaudez D, Chalot M. Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi. FEMS Microbiology Letters, 2006, 254(2): 173- 181.
[10]Itoo Z A, Reshi Z A. The multifunctional role of ectomycorrhizal associations in forest ecosystem processes. Botanical Review, 2013, 79(3): 371- 400.
[11]Khosla B, Kaur H, Reddy M S. Influence of ectomycorrhizal colonization on the growth and mineral nutrition ofPopulusdeltoidesunder Aluminum toxicity. Journal of Plant Interactions, 2009, 4(2): 93- 99.
[12]袁玲, 方德华, 汪智慧, 魏兴元, 黄建国. 钾对外生菌根真菌的分泌作用及氮、磷、钾含量的影响. 生态学报, 2001, 21(2): 254- 258.
[13]Khosla B, Reddy S. Response of ectomycorrhizal fungi on the growth and mineral nutrition of eucalyptus seedlings in Bauxite mined soil. American-Eurasian Journal of Agricultural & Environmental Sciences, 2008, 3(1): 123- 126.
[14]王明霞, 袁玲, 周志峰, 杨红军, 黄建国. 铝对外生菌根真菌草酸分泌及氮磷钾吸收的影响. 林业科学, 2012, 48(2): 82- 88.
[15]Page A L. Methods of Soil Analysis. Madison Wisconsin: Soil Science Society of America, 1982: 178- 185.
[16]辜夕容, 黄建国. 铝对外生菌根真菌草酸分泌及磷、钾、铝吸收的影响. 生态学报, 2010, 30(2): 357- 363.
[17]Heim A, Brunner I, Frossard E, Luster J. Aluminum effects onPiceaabiesat low solution concentrations. Soil Science Society of America Journal, 2003, 67(3): 895- 898.
[18]辜夕容, 梁国仕, 杨水平, 陈翠玲, 黄建国. 接种双色蜡蘑对马尾松幼苗生长、养分和抗铝性的影响. 林业科学, 2005, 41(4): 199- 203.
[19]Dighton J, Skeffington R A. Effects of artificial acid precipitation on the mycorrhizas of scots pine seedlings. New Phytologist, 1987, 107(1): 191- 202.
[20]Arnolds E. Decline of ectomycorrhizal fungi in Europe. Agriculture, Ecosystems & Environment, 1991, 35(2/3): 209- 244.
[21]Tahara K, Norisada M, Tange T, Yagi H, Kojima K. Ectomycorrhizal association enhances Al tolerance by inducing citrate secretion inPinusdensiflora. Soil Science and Plant Nutrition, 2005, 51(3): 397- 403.
[22]Naik D D, Desai S, Smith E W, Cumming J R. Ectomycorrhizal fungipaxillusinvolutusprime the poplar seedling against aluminum stress by modulating ascorbate glutathione and phospholipase pathway: A transcriptome study// Proceeding of 95th ESA Annual Meeting. Pittsburgh, Pennsylvania: ESA, 2010.
[23]Browning M H R, Hutchinson T C. The effects of aluminum and calcium on the growth and nutrition of selected ectomycorrhizal fungi of jack pine. Canadian Journal of Botany, 1991, 69(8): 1691- 1699.
[24]Ahonen-Jonnarth U, Van Hees P A W, Lundström U S, Finlay R D. Organic acids produced by mycorrhizalPinussylvestrisexposed to elevated aluminium and heavy metal concentrations. New Phytologist, 2000, 146(3): 557- 567.
[25]Lapeyrie F, Chilvers G A, Bhem C A. Oxalic acid synthesis by the mycorrhizal funguspaxillusinvolutus(Batsch. ex Fr.) Fr. New Phytologist, 1987, 106(1): 139- 146.
[26]Gonzalez J A Z, Costa M D, Silva I R, Neves J C L, De Barros N F, Borges A C. Accumulation of oxalic acid and calcium crystals in eucalypt ectomycorrhizas. Ⅰ-oxalic acid production and nutrient concentration in fine lateral roots colonized with ectomicorrhizal fungi. Revista Brasileira de Ciencia do Solo, 2009, 33(3): 541- 553.
[27]王明霞, 周志峰, 袁玲, 黄建国. 铝胁迫下钙信号抑制剂对外生菌根真菌分泌草酸的影响. 中国农业科学, 2012, 45(5): 902- 908.
[28]Itoo Z A, Reshi Z A. The multifunctional role of ectomycorrhizal associations in forest ecosystem processes. Botanical Review, 2013, 79(3): 371- 400.
[29]Khosla B, Kaur H, Reddy M S. Influence of ectomycorrhizal colonization on the growth and mineral nutrition ofPopulusdeltoidesunder Aluminum toxicity. Journal of Plant Interactions, 2009, 4(2): 93- 99.
Response of ectomycorrhizal fungi from different environments to aluminum stress
ZHOU Zhifeng, WANG Mingxia*, YUAN Ling, HUANG Jianguo
CollegeofResourcesandEnvironment,SouthwestUniversity,Chongqing400716,China
Abstract:In this study, to select aluminum (Al3+)-tolerant ectomycorrhizal (ECM) fungi and study the resistance mechanism of ECM fungi under Al3+-stress, 10 ECM fungi strains were selected from different environments and cultivated in liquid media with variable concentrations of Al3+(0, 0.20, 0.40, and 1.00 mmol/L). The biomass, efflux of organic acid, and composition of nutrient elements were measured during cultivation. Of the studied fungal strains, Sl 08 showed the strongest resistance to Al3+, while Pt 715, Ld 03, Bo 11, Sl 01, and Bo 15 were also resistant to Al3+. The growth of other tested fungal strains (Sl 14, Gc 99, Cg 04, and Sg 11) was obviously inhibited by Al3+, but that of Sg 11 was most sensitive to Al3+. These results indicated that the ability of ECM fungi to tolerate Al3+toxicity may be closely related to their original growth environments, and fungal strains isolated from acidic soils in south China might be more tolerant to Al3+compared to those isolated from calcareous soils in north China. ECM fungi excrete various organic acids, but the amount and type of these acids vary depending on the fungal strain. For example, ECM fungi could increase the secretion of oxalate to alleviate the effects of Al3+toxicity. In this study, oxalate secretion was observed to increase in the majority of Al3+-resistant strains, and the highest oxalate production was observed in the most Al3+-resistant strain (Sl 08). The secretion of oxalate was observed to decrease in some fungal strains (Bo 11 and Sl 01) that were still resistant to Al3+, whereas oxalate secretion was observed to increase in all Al3+-sensitive strains (Cg 04, Sg 11, Sl 14, and Gc 99). This finding suggests that oxalate secretion is not the only pathway through which ECM fungi can alleviate effects of Al3+toxicity. In addition, the fungal strains showed different characteristics in the uptake of nitrogen (N), phosphorus (P), and potassium (K) under Al3+stress. The N, P, or K content in most of the tested fungi increased in the presence of Al3+, and only Al-sensitive Sl 14 showed no clear difference in the uptake of N, P, or K under these conditions. The N, P, and K content increased both in the resistant (Pt 715 and Bo 11) and in the sensitive (Cg 04) strains, and the content of two of three elements increased in the resistant (Sl 01, N and K) and sensitive (Sg 11, P and K) strains, respectively. Furthermore, the content of one element (N, P, or K) increased in the resistant (Sl 08, Ld 03, Bo 15) and sensitive (Gc 99) strains. Therefore, it appears that oxalate secretion may be an essential pathway for ECM fungi to tolerate effects of Al3+toxicity. In addition, regulation of the uptake of the nutrient elements, N, P, and K, may be important for ECM fungi to alleviate stress induced by Al3+, and increasing the uptake of N, P, and K may be essential for the fungi to tolerate the effects of Al3+stress.
Key Words:environment; ectomycorrhizal fungi; Al3+; organic acids; nutrients
基金项目:国家自然科学研究基金资助项目(41171215);中央高校基本科研业务费资助项目(XDJK2014B047, XDJK2013C044)
收稿日期:2015- 03- 20;
修订日期:2015- 09- 28
*通讯作者
Corresponding author.E-mail: wangmx@swu.edu.cn
DOI:10.5846/stxb201503200533
周志峰, 王明霞, 袁玲, 黄建国.不同生境外生菌根真菌对铝胁迫的响应.生态学报,2016,36(10):2842- 2850.
Zhou Z F, Wang M X, Yuan L, Huang J G.Response of ectomycorrhizal fungi from different environments to aluminum stress.Acta Ecologica Sinica,2016,36(10):2842- 2850.
