Fish Hunting Ankylosaurs (Dinosauria, Ornithischia) from the Cretaceous of China
2016-07-19JIQiangWUXiaochunCHENGYennienTENFangfangWANGXuriJIYannan
JI Qiang, WU Xiaochun, CHENG Yennien, TEN Fangfang, WANG Xuri, JI Yannan
(1. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China; 2. Canadian Museum of Nature, Ottawa ON K1P 6P4, Canada; 3. National Museum of Natural Science, Taichung 40453, Taiwan, China; 4. Xinghai Museum of Paleontology, Dalian 116023, Liaoning, China; 5. China Geological Environmental Monitoring Institute, Beijing 100081, China)
Fish Hunting Ankylosaurs (Dinosauria, Ornithischia) from the Cretaceous of China
JI Qiang1, WU Xiaochun2, CHENG Yennien3, TEN Fangfang4, WANG Xuri1, JI Yannan5
(1. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China; 2. Canadian Museum of Nature, Ottawa ON K1P 6P4, Canada; 3. National Museum of Natural Science, Taichung 40453, Taiwan, China; 4. Xinghai Museum of Paleontology, Dalian 116023, Liaoning, China; 5. China Geological Environmental Monitoring Institute, Beijing 100081, China)
Abstract:All ornithischian dinosaurs are herbivorous or omnivorous. Ornithischian Liaoningosaurus paradoxus Xu et al., 2001 is an ankylosaur. Here we report a new specimen ofn L. paradoxus from China. It contains a number of fish skeletons. We interpret those remains as stomach or gut contents and hence as strong evidence for the meat-eating diet of the dinosaur. With elongate and fork-like denticles of cheek tooth crowns, L. paradoxus has a dentition capable of penetrating into animals like small fishes. The carnivorous adaptation of the dinosaur is also supported by the ungual modification to a sharp claw in both the fore-and hind-limbs. The evolution of a shield-like ventral armor plate and the loose sacrum-pelvic connection suggest that L. paradoxus may have adopted an aquatic way of life, using the ventral armor plate to protect the body from underwater attacks; as such, the open suture between the neural arch and centrum of the vertebrae cannot be used to indicate the juvenile nature of the type specimen. L. paradoxus is the first carnivorous ornithischian dinosaur since dinosaur was first known in the 18th century and represents not only the first aquatic or semi aquatic example of armored dinosaurs but also the smallest species of ornithischian dinosaur so far known.
Keywords:ankylosauria; nodosaurids; Early Cretaceous; Yinxian Formation; Western Liaoning
0Introduction
Dinosaurs consist of two groups: Ornithischia and Saurischia (including birds). The members of the former are all plant-eaters or omnivores and all meat-eating dinosaurs belong to the group Theropoda of the latter (Weishampel et al., 2004; Butler et al., 2010); there have been no exception since a dinosaur was first described scientifically in 1824 (Buckland, 1824).Liaoningosaurusparadoxuswas a nodosaurid ankylosaur (Xu et al., 2001) although later provisionally assigned as an indeterminate taxon within Ankylosauria (Vickaryous et al., 2004). The holotype ofL.paradoxusis less than 34 cm in total length. It was considered as a juvenile owing to open suture between the neural arch and the centrum of vertebrae and also
claimed as the smallest ankylosaur specimen then known (Xu et al., 2001). In this paper, we present convincing evidence of a carnivorous diet forL.paradoxusfrom a newly collected specimen and demonstrate that this dinosaur may have adapted to an aquatic or semi-aquatic way of life, unique among ornithischian dinosaurs.

Fig.1 The holotype of Liaoningosaurus paradoxus (Xu et al., 2001)
1The new specimen of L. paradoxus and its anatomical features
The new specimen, xhpm-1206 (Xinghai Museum of Paleontology), was collected from the Jianshangou Bed of the Lower Cretaceous Yixian Formation at Dawangzhangzi Village, Lingyuan County, west edge of Liaoning Province, about 160 km west and slightly south of Wangjiagou, Yixian, where the holotype ofL.paradoxus
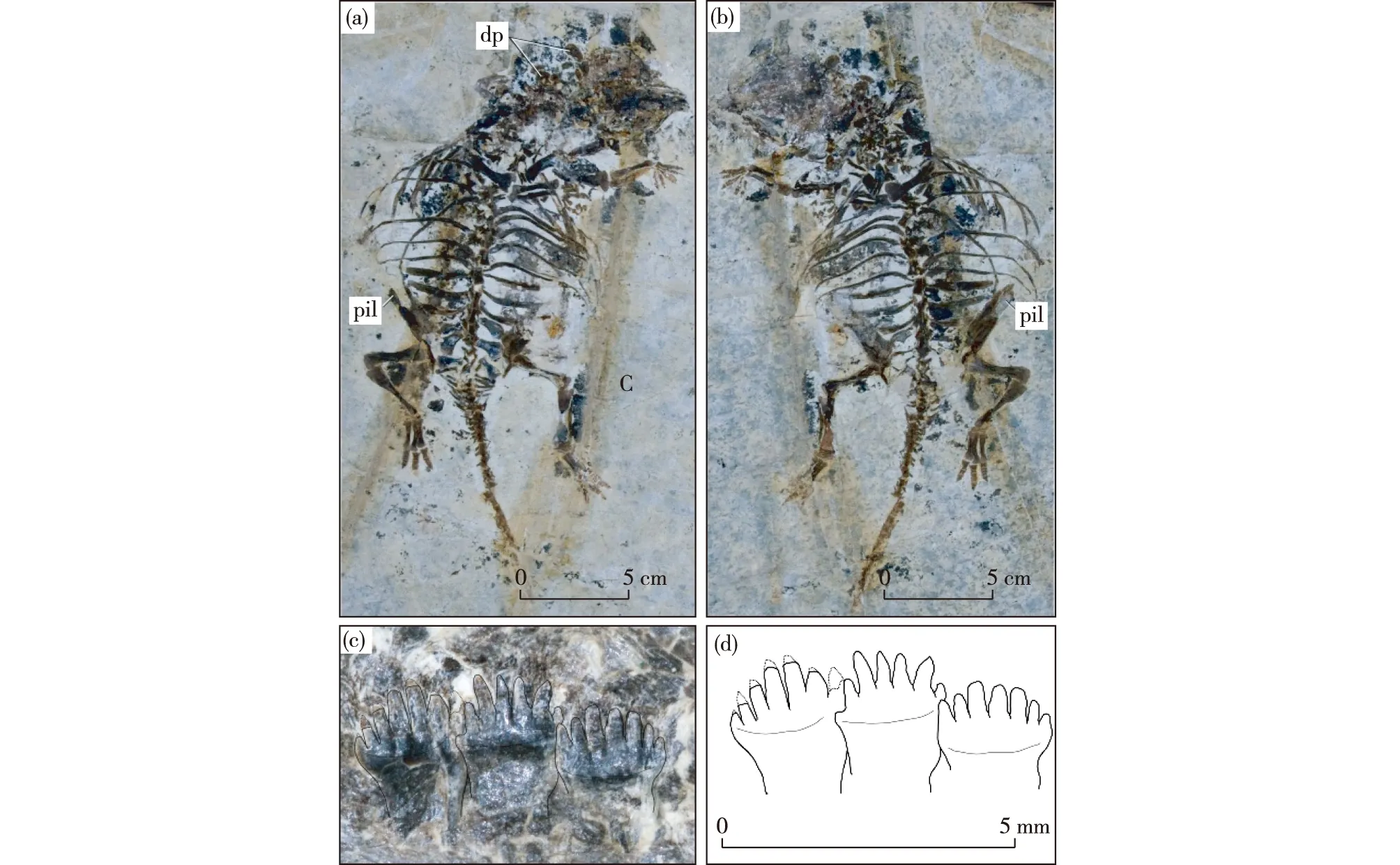
Fig.2 New specimen of Liaoningosaurus paradoxus (xhpm-1206)(a) the positive dorsal view; (b) the negative counterparts of the skeleton; (c) three maxillary teeth in lateral view, showing fork-like crown; (d) the outline of (c) dp-dorsal bony plate around the shoulder girdle; pil-preacetabular process of the ilium
was unearthed from the same formation. This specimen is a nearly complete skeleton but splits into counterparts during collection (Figs.2a, b). It can be compared withL.paradoxuson a number of similarities, such as the unexpected small manus relative to the pes (Figs.2a, b; Figs.3c, e). It is a little larger than the holotype in size as indicated by the skull-sacral length (with not tail), i.e., 19.3 cm versus about 17.2 cm of the latter.
The new specimen is exposed in dorsal view and shows a number of the anatomical features unknown or not accurately described in the holotype. For example, the manus bears five fingers rather than four and the humerus is almost as long as the femur or the tibia (Figs.2a, b; Fig.3a). The sacrum consists of three unfused vertebrae and the expanded distal end of the sacral ribs are separated from one another, which differ from those in other ankylosaurs whose synsacrum is formed and the sacral ribs are
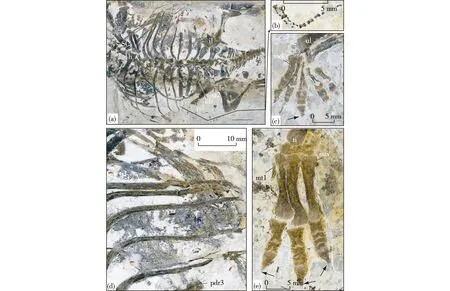
Fig.3 New specimen of Liaoningosaurus paradoxus (xhpm-1206)(Reddish arrows pointed at fish skeletons within the ribcage and black arrows pointed at sharp ungual)(a) the trunk region from the positive counterpart in dorsal view, showing gut contents, loose sacrum-pelvic joints, and sutural connections between caudal ribs and vertebrae; (b) close-up of a section of a reptilian tail; (c) the right manus from negative counterpart; (d) close-up of the area just anterior to the right ilium in the positive counterpart, showing fishes with a vague body outline; (e) the left pes from the negative counterpartfe-femur; h-humerus; il-ilium; ldr-the last dorsal rib; mc1-metacarpal 1; mt1, mtv-metatarsals 1, 5; pdr3-the third dorsal rib from the sacrum; pil- preacetabular process of ilium; sc-scapula; sr2-sacral ribs 2; ti-tibia; ul-ulna.
fused together distally (Coombs,1978; Coombs et al.,1990; Garcia et al., 2009). As preserved, the sacral ribs do not form a firm connection with either the pelvis (ilium) laterally or the sacral vertebrae medially (Fig.3a), this is also the case of the holotype (Fig.4a). Thus, it is supposed that this feature, together with the open suture between the neural arch and the centrum of vertebrae and the lack of a synsacrum, indicates that xhpm-1206 is a juvenile, as previously proposed for the holotype (Xu et al., 2001). The caudal transverse processes/ribs are sutured to but not fused with the vertebrae as in the sacral region (Fig.2a; Fig.3a); this is also observed in the holotype (Fig.3a) but not in the other ankylosaurs (Coombs et al., 1986; Coombs, 1995). In addition, hundreds of specimens ofL.paradoxushave been unearthed from the Lower Cretaceous of Western Liaoning, but no dorsal scutes have been found.
2Carnivorous adaptation of L. paradoxus
Incredibly, xhpm-1206 (the positive counterpart) shows a number of fish skeletons within its ribcage. There may be three explanations.
(1) Those fish skeletons were dead bodies lying on the bottom before the ankylosaur sunk down and later passively went into the ribcage of the covering ankylosaur skeleton because of underwater turbulence. This interpolation appears too speculative; the main reason against it is the fact that no fish skeletons are present outside the ankylosaur body. In other words, there should be some fish skeletons preserved around the ankylosaur skeleton or nearby if those fishes died earlier. It is true that one fish skeleton is seen anterior to the right femur, which may have been pushed out from the body cavity post-mortem (Fig.3a). Another line of evidence against the assumption comes from the nature of those fish skeleton, i.e., none of them shows a clear appearance as seen in true fish specimens collected from the same horizon.
(2) Those fishes may have died within the ribcage of the ankylosaur when they were scavenging or using the skeleton as a hiding place. This assumption is also not convincing. As described in the type specimen,L.paradoxusis ventrally covered by a bone plate that is hard to be penetrated by those small fishes. On the other hand, the exposed dorsal side of the new skeleton is nearly complete, showing no obvious breakage made by predators. As mentioned earlier, those fish skeletons all show a vague and incomplete appearance, which are in conflict with this assumption too.
(3) Contrary to the aforementioned two assum-ptions, it is proposed that those fish skeletons came from the stomach or gut contents of the specimen. Most of those fishes are incomplete and almost all of them display an unclear body outline, which should have been obscured by digesting acid during food processing in the stomach (Fig.2d). Besides, there is a section of a reptile’s tail on the posterolateral side of the ribcage (Figs.2a, b). It is hard to believe that a reptile broke its tail inside the ribcage when it withdrew after scavenging, so it should be gut leftovers of the reptile. Those fish skeletons are not tightly packed in the stomach but scattered on both sides of the body cavity; this is because they may have been expelled out by gases or compression from the stomach and gut post-mortem. Therefore, it is considered that those fish skeletons and the reptile′s tail are gut contents. In other words,L.paradoxusis literally a predator, eating these fishes in life. This strongly suggests that the ankylosaur may have adapted to an aquatic or a semi-aquatic way of life. Such a lifestyle of the dinosaur may be also demonstrated by the tooth morphology and a number of the structural modifications of the postcranial skeleton.
Teeth are well preserved in the holotype. Our further preparation reveals that the morphology of the cheek teeth differs considerably from that of any herbivorous dinosaur. The marginal denticles of the palmate crowns are deeply separated from each other and thus individual denticle is elongate and needle-like, leading to a fork-shaped outline of tooth crowns (Figs.2c, d). The denticles of the incisor of the type specimen are also much sharper than in other ankylosaurs although they are shorter than those of the cheek teeth (Figs.4b, c).With such a dentition,L.paradoxuswas capable of catching fishes or small reptiles and penetrating them.
3L. paradoxus′ adaptation to aquatic way of life
L.paradoxusis characterized by a shield-like ventral abdominal plate whose function was previously unknown (Fig.4a) (Xu et al., 2001). The evolution of the ventral plate may have been closely related to an aquatic way of life. The ventral plate, if complete, would have covered the entire ventral surface of the trunk (Xu et al., 2001), and would be functionally comparable to the ventral dermal armor (plastron) of the most primitive turtle,Odontochelys, from the marine Triassic of China (Li et al., 2008a). This turtle with no complete dorsal carapace has been considered as an aquatic animal and the evolution of a ventral dermal armor (plastron) has been taken to indicate an aquatic origin of turtles (Rieppel et al., 1999), i.e., the ventral plastron provides dermal protection for the ventral body surface from underwater attacks and such a ventral bony plate is not needed for land tetrapods (with four limbs) because the relevant body surface is not immediately exposed to predatory attack.

Fig. 4 Partial skeleton of the holotype of Liaoningosaurus paradoxus (IVPP V12560) in (a) to (c) and the sacral region of two Triassic marine reptiles in (d) and (e)(a) most parts of the postcranial skeleton in dorsal view; (b) close-up of a premaxillary tooth in lateral view; (c) the outline of (b); (d) the sacral region of a pregnant specimen of Keichousaurus hui (a Triassic marine reptile); arrows indicating embryos; (e) the sacral region of Wumengosaurus delicatomandibularis (a Triassic marine reptile)car1-the 1st caudal rib/transverse process; fi-fibula; is-ischium; pu-pubis; r-radius; sr-sacral rib; sr3-sacral rib 3; vp-shield-like ventral bony plate
As mentioned earlier, there were some features previously considered as evidence to indicate that the holotype ofL.paradoxuswas a juvenile specimen (Xu et al., 2001). The most striking features include the open neural arch-central sutures of vertebrae and the loose sacrum-pelvic connections (implying mobility of a certain kind in joints). Those features might support such an assumption of juvenileness if the dinosaur lived on land (Coombs et al., 1986) but may not if it is an aquatic or semi-aquatic animal. It has been well documented that in most Triassic marine reptiles such as eosauropterygians (Carroll et al., 1985; Rieppel, 1989; Sander, 1989; Storrs, 1991; Sato et al., 2010; Wu et al., 2011), sutures between the neural arch and the centrum of vertebrae remained open in adulthood (at least in young adults), the sacrum and pelvis were connected by kinetic joints in their whole life (Figs. 4d, e). On the other hand, it has been well supported that the reptiles with a kinetic pelvis (the sacral ribs lose any firm articulation with either the vertebral column medially or the ilium laterally) must have adapted to a fully aquatic way of life (Cheng et al., 2004; Ji et al., 2010). This is because aquatic forms do not need a strong and solid sacrum-pelvic base to support the total body weight when living in the water. In addition, the caudal ribs are always fused to the vertebrae in juveniles of terrestrial dinosaurs including ankylosaurs (Coombs et al., 1986; Coombs, 1995) but they are sutured with the vertebrae inL.paradoxus, as in the Triassic aquatic reptiles. Therefore, the aforementioned features do not provide firm evidence of juvenileness forL.paradoxusbut rather indicate an aquatic or a semi-aquatic habit of the ankylosaur. Another line to suggest that both the holotype and the new specimen ofL.paradoxusare adult may come from the fact that all known specimens of the ankylosaur are small, similar in size. There have been a number of the specimens ofL.paradoxusunearthed from the Yixian Formation since 2001. Similar to the holotype and xhpm-1206, none of them is large-sized.For example, NGMC-98-003(National Geological Museum of China) has a preserved length of 22 cm, with a skull-sacral length of about 16 cm, and GPMA-12-045(Geological and Paleontological Museum of Anhui) has a skull-sacral length of about 21 cm, suggesting a total body length still less than 45 cm if the tail length was nearly as long as the skull-sacral length as in extant alligators (Cong et al., 1988). With no exception, these specimens also show unfused neural arch-central sutures in vertebrae, no formation of a synsacrum, and no fusion of the caudal ribs with the vertebrae. Therefore, the type specimen ofL.paradoxusis not a juvenile but represents a true small-sized ankylosaur with a total body length less than 50 cm. This is evidently smaller thanFruitadens(65~75 cm) orTianyusaurus(around 70 cm), two of which are newly claimed the smallest ornithischian dinosaurs (Zheng et al., 2009; Butler et al., 2010) and thusL.paradoxusis now the smallest ornithischian dinosaur known in the world.
Compared with those fully aquatic Triassic reptiles (Figs.4d,e), the sacral ribs are not simply bar-shaped but massive and strongly expanded distally inL.paradoxus(Figs.3a,4a), as the case seen in the semi-aquatic choristoderans, such asIkechosaurusof the Early Cretaceous (Brinkman et al.,1993) orSimoedosaurusof the Late Cretaceous (Erickson,1987). With such sacral ribs and the strong dorsal blade of the ilium, the sacrum-pelvic connection ofL.paradoxusshould not have been too loose or too weak to support the body weight on land, and it was, together with the well-developed four limbs, most probably capable of walking on land. Therefore, we believe thatL.paradoxusmay have lived near a river-lake system, a watery environment and adapted to an amphibious way of life, probably like derived choristoderans such as the Late Cretaceous champsosaurs (Erickson, 1972), foraging mainly in the water. Taking the advantage of such a lifestyle,L.paradoxuswould have avoided intense competition with contemporaneous terrestrial carnivorous non-avian dinosaurs, birds, lizards, and mammals (Zhang et al., 2003; Ji et al., 2004; Li et al., 2008b).
4Conclusions
(1) OrnithischianLiaoningosaurusparadoxusXu et al., 2001 is an ankylosaur.A newly found specimen ofL.paradoxusfrom China contains a number of fish skeletons. We interpret those remains as stomach or gut contents and hence as strong evidence for the carnivorous diet of the dinosaur. With elongate and fork-like denticles of cheek tooth crowns,L.paradoxushas a dentition capable of penetrating into animals like small fishes. Its carnivorous adaptation is also supported by the ungual modification to a sharp claw in both the fore- and hind-limbs.
(2) The shield-like ventral armor plate and the loose sacrum-pelvic connection suggest thatL.paradoxusmay have adopted an aquatic way of life, using the ventral armor plate to protect the body from underwater attacks; as such, the open suture between the neural arch and centrum of the vertebrae cannot be used to indicate the juvenile nature of the type specimen.L.paradoxusis the first carnivorous ornithischian dinosaur since dinosaur was first known in the 18thcentury and represents not only the first aquatic or semi-aquatic example of armored dinosaurs but also the smallest species of ornithischian dinosaur so far known.

Fig.5 Reconstruction of L. paradoxus
Acknowledgements
ThisworkwassupportedbygrantsfromtheMinistryofScienceandTechnologyofChina(2012CD822000)andChinaGeologicalSurvey(121202114026801)toQ.J;fromCanadianMuseumofNature(RS34)toX.-c.W;andfromNationalScienceFoundation(NSC-100-2116-M-178-003),Taiwan,ChinatoY.-n.C.WearegratefultoXinghaiMuseumofPaleontologyforprovidingthespecimen.WethankYu-qingZhangforthepreparationofthefossilandNGMCandGPMAforprovidingthedataofthespecimensofL. paradoxusintheircare.X.-c.WwouldliketothankInstituteofGeology,ChineseAcademyofGeologicalSciencesforthehospitalityduringhisstayinBeijingandXingXuofInstituteofVertebratePaleontologyandPaleoanthropology(IVPP),ChineseAcademyofSciencesandLiang-fangYangandLie-wuLuofNGMCforarrangingtheexaminationoftheholotypeandotherspecimensofL. paradoxus.
References
BUCKLANDW, 1824.NoticeontheMegalosaurusorgreatfossillizardofStonesfield[J].TransactionsoftheGeologicalSocietyofLondon, 1(2): 390-396.
BRINKMANDB,DONGZhiming, 1993.NewmaterialofIkechosaurus sunailinae (Reptilia:Choristodira)fromtheEarlyCretaceousLuohongdongFormation,OrdosBasin,InnerMongolia,andtheinterrelationshipsofthegenus[J].CanadianJournalofEarthSciences, 30(10): 2153-2162.
BUTLERRJ,GALTONPM,PORROLB,etal, 2010.LowerlimitsofornithischiandinosaurbodysizeinferredfromanewUpperJurassicheterodontosauridfromNorthAmerica[J].ProceedingsoftheRoyalSocietyB:BiologicalSciences, 277(1680): 375-381.
COOMBSWPJr, 1978.ThefamiliesoftheornithischiandinosaurOrderAnkylosauria(Reptilia,Ornithischia)[J].Palaeontology, 21: 143-170.
CARROLLRL,GASKILLP, 1985.ThenothosaurPachypleurosaurusandtheoriginofplesiosaurs[J].PhilosophicalTransactionsoftheRoyalSocietyBBiologicalSciences, 309(1139): 343-393.
COOMBSWP,COOMBSWPJr, 1986.AJuvenileankylosaurreferabletothegenusEuoplocephalus (Reptilia:Ornithischia).JournalofVertebratePaleontology, 6(2): 162-173.
CONGLinyu,HOULianhai,WUXiaochun, 1988.TheGrossAnatomyofAlligatorsinensisFauvel[M].Beijing,China:SciencePress.
COOMBSWPJr,MARYANSKAT, 1990.Ankylosauria[C]//WEISHAMPELDB,DODSONP,OSMLSKAH.TheDinosauria.Berkeley,USA:UniversityofCaliforniaPress, 456-483.
COOMBSWPJr, 1995.Anewnodosauridankylosaur(Dinosauria:Ornithischia)fromtheLowerCretaceousofTexas[J].JournalofVertebratePaleontology, 15(2): 289-312.
CHENGYennien,WUXiaochun,JIQiang, 2004.Triassicmarinereptilesgavebirthtoliveyoung[J].Nature, 432(7015): 383-386.
ERICKSONBR, 1972.TheLepidosaurianReptileChamp sosaurusinNorthAmerica[M].SaintPaul,MN,USA:ScienceMuseumofMinnesota.
ERICKSONR, 1987. Simoedosaurus dakotensis,newspecies,adiapsidreptile(Archosauromorpha:Choristodera)fromthePaleoceneofNorthAmerica[J].JournalofVertebratePaleontology, 7(3): 237-251.
GARCIAG,PEREDA-SUBERBIOLAX, 2009.AnewspeciesofStruthiosaurus (Dinosauria:Ankylosauria)fromtheUpperCretaceousofVilleveyrac(southernFrance)[J].JournalofVertebratePaleontology, 23(1): 156-165.
JIQiang,CHENW,WANGWL,etal, 2004.MesozoicJeholBiotaofWesternLiaoning[M].Beijing,China:GeologicalPublishingHouse.
JIQiang,WUXiaochun,CHENGYennien, 2010.Cretaceouschoristoderanreptilesgavebirthtoliveyoung[J].Naturwissen, 97(4): 423-428.
LIChun,WUXiaochun,RIEPPELO,etal, 2008a.AnancestralturtlefromtheLateTriassicofsouthwesternChina[J].Nature, 456(7221): 497-501.
LIJinling,WUXiaochun,ZHANGFucheng, 2008b.TheChineseFossilreptilesandTheirKin[M].Beijing,China:SciencePress.RIEPPELO, 1989.Anewpachypleurosaur(Reptilia:Sauropterygia)fromtheMiddleTriassicofMonteSanGiorgio,Switzerland[J].PhilosophicalTransactionsoftheRoyalSocietyB:BiologicalSciences, 323(1212): 1-73.
RIEPPELO,REISZRR, 1999.Theoriginandearlyevolutionofturtles[J].AnnualReviewofEcologyandSystematics, 30(1): 1-22.
SANDERPM, 1989.Thepachypleurosaurids(Reptilia:Nothosauria)fromtheMiddleTriassicofMonteSanGiorgio(Switzerland)withthedescriptionofanewspecies[J].PhilosophicalTransactionsoftheRoyalSocietyB:BiologicalSciences, 325(1230): 561-666.
STORRSGW, 1991.AnatomyandrelationshipsofCorosaurus alcovensis (Diapsida:Sauropterygia)andtheTriassicAlcovaLimestoneofWyoming[J].BulletinofthePeabodyMuseumofNaturalHistory, 44: 1-151.
SATOT,CHENGYennien,WUXiaochun,etal, 2010.OsteologyofYunguisaurusChengetal., 2006 (Reptilia:Sauropterygia),aTriassicpistosauroidfromChina[J].PaleontologicalResearch, 14(3): 179-195.
VICKARYOUSMK,MARYANSKAT,WEISHAMPELDB, 2004.ChapterSeventeen:Ankylosauria[C]//WEISHAMPELDB,DODSONP,OSMLSKAH.TheDinosauria. 2nded.Berkeley,USA:UniversityofCaliforniaPress, 363-392.
WEISHAMPELDB,DODSONP,OSMLSKAH, 2004.TheDinosauria[M].Berkeley,USA:UniversityofCaliforniaPress.
WUXiaochun,CHENGYennien,LIChun,etal, 2011.NewinformationonWumengosaurus delicatomandibularisJiangetal., 2008 (Diapsida:Sauropterygia),withrevisionoftheosteologyandphylogenyofthetaxon[J].JournalofVertebratePaleontology, 31(1): 70-83.XUXing,WANGXiaolin,YOUHailu, 2001.AjuvenileankylosaurfromChina[J].Naturwissenschaften, 88(7): 297-300.ZHANGMiman,CHENPeiji,WANGYuanqing,etal, 2003.TheJeholBiota:TheEmergenceofFeatheredDinosaurs,BeakedBirdsandFloweringPlants[M].Shanghai:ShanghaiScienceandTechnologyPress.
ZHENGXiaoting,YOUHailu,XUXing,etal, 2009.AnEarlyCretaceousheterodontosauriddinosaurwithfilamentousintegumentarystructures[J].Nature, 458(7236): 333-336.
doi:10.3969/j.issn.1674-3636.2016.02.183
Corresponding author:WU Xiaochun. E-mail: xcwu@mus-nature.ca
Manuscript received Mar. 8, 2016; Edited by ZHAN Gengshen
About the first author: JI Qiang, male, researcher, doctor degree, PhD supervisor, majored in stratigraphic palaeontology, interested in Neopaleozoic conodonts biostrata and late mesozoic palaeovertebrates. E-mail: jirod@cags.ac.cn
