分子标记辅助选择改良三系杂交稻恢复系R225稻瘟病抗性
2016-06-30聂元元李霞毛凌华颜满莲颜龙安蔡耀辉
聂元元 李霞 毛凌华 颜满莲 颜龙安 蔡耀辉*
(1江西省超级水稻研究发展中心/国家水稻工程实验室,南昌330200;2江西省农业科学院水稻研究所/国家水稻改良中心南昌分中心,南昌330200;*通讯作者)
分子标记辅助选择改良三系杂交稻恢复系R225稻瘟病抗性
聂元元1李霞2毛凌华1颜满莲1颜龙安1蔡耀辉1*
(1江西省超级水稻研究发展中心/国家水稻工程实验室,南昌330200;2江西省农业科学院水稻研究所/国家水稻改良中心南昌分中心,南昌330200;*通讯作者)
摘要:稻瘟病是水稻的主要病害,培育抗稻瘟病品种是防治稻瘟病的有效途径。本研究通过分子标记辅助选择与杂交育种相结合的方式,将稻瘟病抗性基因Pi1、Pi2和Pi9导入到三系杂交稻恢复系R225。对BC3F3代材料进行苗期和成熟期稻瘟病抗性鉴定,携带1个或2个抗性基因的目标株系抗性达到中抗以上水平,稻瘟病抗性显著高于各自的轮回亲本。SSR标记分析表明,改良株系的遗传背景回复率达到86.1%~95.3%。通过标记辅助选择获得的改良材料为三系杂交稻恢复系的培育提供了稻瘟病抗性亲本。
关键词:标记辅助选择;稻瘟病;三系杂交稻;恢复系
稻瘟病是由真菌Magnaporthe oryzae引起的水稻毁灭性病害,是水稻生产的主要病害,选育和应用抗病品种是防治稻瘟病、减少其危害的有效途径[1],同时也是保护环境、发展绿色水稻产业、实现水稻可持续性发展和确保粮食安全的迫切需要。然而,由于稻瘟菌小种的高度变异性,大面积推广的水稻品种含有的抗性基因往往在3~5年之后就失去抗性,导入广谱抗性基因和选育抗瘟性强的水稻品种是目前最常用的预防稻瘟病的方法[2]。
目前已标记定位的稻瘟病抗性基因达80多个,不同稻区稻瘟病的生理小种差异很大,因此,利用什么样的抗病基因进行品种改良,也会因稻区的不同而不同。此外,用于杂交稻育种的基因必须具有较强的显性作用。在本研究中选用的Pi1对我国792个稻瘟病小种的绝大部分表现抗性,其抗性频率达89.65%,是一个广谱的稻瘟病抗性基因[3],利用RM224和MRG4766对Pi1选择的准确率达100%[4]。Pi2也是一个广谱抗性基因,对455个菲律宾小种及来自16个国家和地区的43个小种中的36个小种表现抗性,Zhou等[5]已克隆该基因。而Pi9基因来自小粒野生稻Oryza minuta,对所有供试菲律宾小种,来自16个国家和地区的43个小种表现抗性[6],Qu等[7]已成功克隆了该基因。Pi2与Pi9是位于第6染色体着丝粒附近位置非常接近的抗病基因。
受体轮回亲本R225是江西省超级水稻研究发展中心选育的优良恢复系,株型适中,分蘖力强,穗大粒多,结实率高,后期落色好。所配组合荣优225于2009年通过江西省审定,并被列为江西省晚籼主栽品种之一,2012年通过国家审定,2014年通过农业部超级稻认定,同时被列为农业部主导品种。荣优225已申请植物新品种保护,具有较好的应用前景[8]。本研究通过分子标记辅助选择和回交育种的策略,利用携带稻瘟病抗性基因Pi1、Pi2基因的材料BL6与含Pi9基因的亲本材料75-1-127分别与R225杂交,将Pi1、Pi2和Pi9基因聚合到优良恢复系R225中,以选育稻瘟病抗性增强的优良株系。
1 材料与方法
1.1试验材料
以含有抗稻瘟病基因Pi1、Pi2的材料BL6和含有抗稻瘟病基因Pi9的材料75-1-127作为本试验的供体,以恢复系R225为受体,通过杂交、回交和分子标记辅助选择,建立回交BC1F1分离群体,同时分子标记检测,选择阳性植株与轮回亲本R225回交,在回交后自交两代获得BC3F3,获得含目标抗性基因的单株,每个世代均用分子标记进行目标基因选择(图1)。
1.2稻瘟病抗性鉴定
稻瘟病抗性鉴定采用大棚人工接种鉴定和井冈山稻瘟病自然鉴定。井冈山鉴定点是国家和省级区试水稻稻瘟病抗性鉴定点,溪沟纵横,四面环山,属典型的山阴梯田,日照时间短,雾露时间长,高温高湿,极有利于稻瘟病的发生。
1.3抗性基因的分子标记检测
采集水稻幼嫩叶片用CTAB法快速提取DNA[9],引物由上海生工合成。选择亲本间扩增片段差异较大引物,用琼脂糖凝胶电泳进行分子标记检测。

图1 分子标记辅助选择聚合稻瘟病抗性基因Pi1、Pi2和Pi9
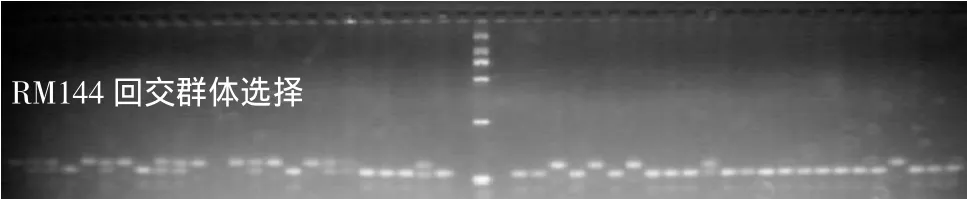
图2 Pi1、Pi2和Pi9基因选择标记RM144、AP22和PB8的分子标记检测结果
2 结果与分析
2.1目标基因选择的分子标记
分别合成RM144、RM224、MRG4766等3对与Pi1基因紧密连锁标记的引物[10],AP22、SRM24、Nbs2P3、RM527等4对与Pi2基因紧密连锁及基因内标记的引物[11],PB8和Nbs-Pi9等2对与Pi9基因紧密连锁或基因内标记的引物[12]。通过PCR扩增、琼脂糖凝胶电泳分子标记检测,筛选本实验中所涉及轮回亲本R225和相应供体亲本BL6、75-1-127间的多态性。根据筛选多态性结果,选择琼脂糖凝胶电泳可以检测供体-受体间差异的标记,分别为RM144对Pi1基因,AP22对Pi2基因,PB8对Pi9基因进行标记辅助选择(图2)。
2.2 BC3F3改良材料的稻瘟病抗性表型鉴定
分子标记辅助选择获得聚合Pi1、Pi2及Pi9基因改良株系116株,聚合Pi1、Pi2基因改良株系327株。对受体、供体亲本及改良株系在井冈山鉴定点进行稻瘟病抗性鉴定。苗期调查分级参照Tharreu D的6级统一标准将病斑分为6级,1~3级为抗病表型,4~6级为感病表型[13]。调查结果表明,轮回亲本R225自然状态下表现感病,抗病级别为5级;供体亲本BL6自然状态下表现抗病,抗病级别为1级,供体亲本75-1-127自然状态下表现抗病,抗病级别为1级;改良群体自然状态下抗病级别为2~4级,其中聚合了Pi1、Pi2及Pi9基因的改良株系抗病效果得到明显提高,抗病级别为1级,聚合了Pi1、Pi2基因的改良株系抗病效果也很明显,抗病级别为1~2级。
成熟后期重点调查穗颈瘟。轮回亲本R225自然状态下表现感病,抗病级别为6级,穗颈瘟严重;供体亲本BL6自然状态下表现抗病,抗病级别为1级;供体亲本75-1-127自然状态下表现抗病,抗病级别为1级;改良群体自然状态下抗病级别为3~4级,其中116株聚合了Pi1、Pi2及Pi9基因的改良株系中表现高抗的有21株,抗病级别为1级,其余95株表现抗病,但是也有少量单株出现穗颈瘟,抗病级别为2~3级;聚合了Pi1、Pi2基因的327株改良株系抗病级别为1~2级,其中96株表现高抗,抗病级别为1级。
对16份携带Pi1、Pi2及Pi9基因的BC3F3代改良材料进行稻瘟病抗性鉴定,调查材料包含R225/BL6、R225/75-1-127、R225/BL6//75-1-127等,每份材料调查30株。调查结果发现,R225表现为中感,抗病级别为4级;BL6表现为高抗,抗病级别为1级;75-1-127表现为高抗,抗病级别为1级;携带有抗性基因的改良株系则表现为抗及中抗水平,不含抗性基因的材料对稻瘟病表现为感病或中感(表1)。
2.3改良株系遗传背景的SSR标记检测
选取BC3F3代材料10个株系进行SSR背景分析,在R225与75-1-127中筛选到101对SSR多态性标记,在R225与BL6间筛选到114对SSR多态性标记,这些引物在水稻12对染色体上近似平均分布,检测的单株标记基因型中轮回亲本基因型的比率在86.1%~95.3%之间,改良株系遗传背景近似轮回亲本。
对获得的BC3F3代材料进行田间农艺性状调查,改良株系与相应轮回亲本R225表型较为相似,改良材料在株型、株高、穗型和后期落色等性状上与轮回亲本表现基本一致。而部分性状上有意识的倾向于选择供体亲本的性状,以弥补轮回亲本的一些缺点,例如,BL6来源的改良材料倾向于短圆粒,且谷壳颜色金黄的材料,而75-1-127来源的改良材料则分蘖力变强,生育期推迟3~5 d。
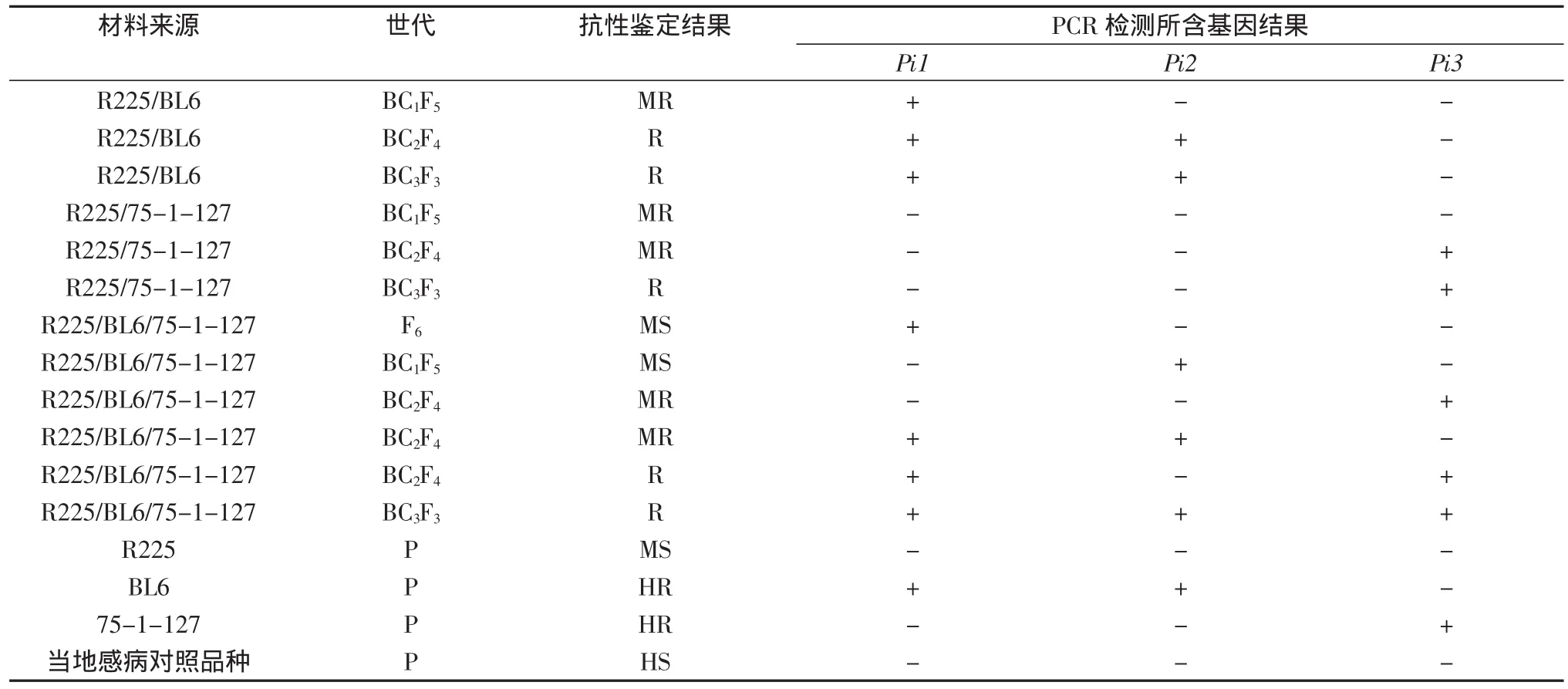
表1 改良后代株系及亲本稻瘟病抗性鉴定结果
3 讨论
随着生物技术的发展,人们发现越来越多的稻瘟病抗性位点与抗性基因[14]。目前已标记定位的稻瘟病抗性基因达68个,其中23个被成功克隆,但不同稻区稻瘟病的生理小种差异很大,因此,利用什么样的抗病基因进行品种改良,也会因稻区的不同而不同。此外,用于杂交稻育种的基因必须具有较强的显性作用[15]。本研究选用具有广谱抗性的稻瘟病抗性基因Pi1、Pi2 和Pi9作为目的基因,通过分子标记辅助选择技术进行水稻抗稻瘟病基因聚合育种,利用与抗性基因紧密连锁的SSR标记同时鉴定多个抗性基因的存在与否,同时结合田间表型鉴定稻瘟病抗性。
育种实践证明,要育成具有广谱抗性的杂交稻新组合,其双亲之一应具有广谱抗性[16-17]。如福建省农科院水稻所育成稻瘟病抗性不育系地谷A、福伊A,以及利用其配组选育而成抗稻瘟病的杂交稻组合地优77、福优964、福优明86等,在武陵山区等稻瘟病重发区种植,表现出较强的稻瘟病抗性水平[18]。本研究通过分子标记辅助选择聚合具有广谱抗性的3个稻瘟病抗性基因,改良恢复系R225稻瘟病抗性,从而达到改良其杂交稻组合稻瘟病抗性的目的。
参考文献
[1]Couch B C,Hohn L M. A multilocus gene genealogy concordan with host preference indicates segregation of a new species Magnaporthe oryzae,from M grisea[J]. Mycologia,2002,94: 683-693.
[2]Hittalmani S,Parco A,Mew T V,et al. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice[J]. Theor Appl Genet,2000,100: 1 121-1 128.
[3]Chen H L,Chen B T,Zhang D P,et al. Pathotypes of pyricularia grisea in rice fields of central and southern China[J]. Plant Disease,2001,85: 843-850.
[4]Mackill D J,Bonman J M. Inheritance of blast resistance in near-isogenic lines of rice[J]. Phytopathology,1992,82: 746-749.
[5]Zhou B,Qu S H,Liu G F,et al. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to magnaporthe grisea [J]. Mol Plant Microbe Interact,2006,19: 1 216-1 228.
[6]Liu G,Lu L,Zeng G L. Two broad-spectrum blast resistance genes,Pi9(t)and Pi2(t),are physically linked on rice chromosome 6[J]. Mol Genet Genomics,2002,267: 472-480.
[7]Qu S H,Liu G F,Zhou B,et al. The broad-spectrum blast resistance gene pi9 encodes a nucleotide -binding site -leucine -rich repeat protein and is a member of a multigene family in rice[J]. Genetics,2006,172: 1901-1914.
[8]聂元元,毛凌华,李永辉,等.三系杂交晚稻荣优225示范推广的现状与展望[J].杂交水稻,2014,29(2):37-39.
[9]楼巧君,陈亮,罗利军.三种水稻基因组DNA快速提取方法的比较[J].分子植物育种,2005,3(5):749-752.
[10]陈志伟,官华忠,吴为人,等.稻瘟病抗性基因Pi-1连锁SSR标记的筛选和应用[J].福建农林大学学报,2005,34(1):74-77.
[11]陈志伟,郑燕,吴为人,等.抗稻瘟病基因Pi-2(t)紧密连锁的SSR标记的筛选与应用,分子植物育种,2004,2(3):321-325.
[12]刘士平,李信,汪朝阳,等.基因聚合对水稻稻瘟病的抗性影响[J].分子植物育种,2003,1(1):22-26.
[13]Tharreau D,Lebrun M H,Talbot N J,et al. New tools for resistance gene characterization in rice[G]. In: Advances in Rice Blast Re-search. Proceedings of the 2nd International Rice Blast Conference. Montpellier,France,1998,Kluwer Academic Publishers,Wageningen,The Netherlands,2000: 54-62.
[14]安正帅,刘国兰,梅捍卫,等.标记辅助改良节水抗旱杂交稻亲本材料的稻瘟病抗性[J].分子植物育种,2010,8(6):1 172-1 176.
[15]卓伟,许旭明,张受刚,等.抗稻瘟病籼型水稻不育系广抗13A的选育[J].杂交水稻,2009,24(2):10-13.
[16]肖叶青,胡兰香,吴小燕,等.野败型不育系“赣香A”的稻瘟病抗性研究[J].分子植物育种,2010,8(6):1 102-1 107.
[17]刘毅,王加红,黎良通,等.分子标记辅助选择Xa23基因改良节水抗旱稻亲本的白叶枯病抗性研究[J].上海农业学报,2014,30 (1):13-16.
[18]张建福,凌忠专,王国英,等.籼型恢复系云恢290稻瘟病抗性遗传学分析[J].分子植物育种,2006,4(4):540-544.
Improving Blast Resistance of Parental Restorer Lines R225 by Marker-assisted Selection
NIE Yuanyuan1,LI Xia2,MAO Linghua1,YAN Manlian1,YAN Longan1,CAI Yaohui1*
(1Jiangxi Super-rice Research and Development Center/National Engineering Laboratory for Rice,Nanchang 330200,China;2Rice Research Institute,Jiangxi Academy of Agricultural Sciences/Nanchang Branch of Chinese National Center for Rice Improvement,Nanchang 330200,China;*Corresponding author)
Abstract:Rice blast is one of the most devastating diseases in rice. Cultivating rice blast resistant varieties is an effective way to control rice blast. Using marker assisted selection and backcrossing,blast resistance genes Pi1,Pi2 and Pi9 were introgressed into restorer parental lines R225. The BC3F3progenies harboring Pi1,Pi2 or Pi9 were selected by linked markers. The lines carrying one or two target genes are significantly higher than their respective recurrent parent. Background SSR analysis showed that the recovery rate of the target lines reached 86.1%~95.3%. The improved lines can be used as candidate parental lines for rice breeding with blast resistance.
Key words:marker-assisted selection(MAS);rice blast;three line hybrid rice;restorer line
中图分类号:S511.03
文献标识码:B
文章编号:1006-8082(2016)03-0060-04
收稿日期:2015-12-11
基金项目:“863”计划“绿色性状基因聚合与种质创新”(2014AA10A603);江西省农科院青年基金(2014CQN 001)
